
Abstract
The majority of studies in short- and middle-term caloric restriction (CR) have been primarily focused on physiological parameters, improvements in aging, modulation of oxidative stress, and long-term negative effects on cognitive functions. However, single-housing associated with CR may pose many logistical problems. Thus, it is necessary to study the effects of CR under conditions in which animals are group-housed. The aims of this study were to (i) observe the possible differences in the proportion of the weights and social behaviour under ad libitum and CR (at 70%) conditions; (ii) examine the eventual inequalities in the proportion of the weights and social behaviour (the time spent eating under the feeder as an indicator of dominance and empathy, and the number of ‘pushes’ as an indicator of aggressiveness) in sibling and non-sibling rats under CR conditions; and (iii) compare the concentrations of corticosterone (stress biomarker) in serum under ad libitum and CR conditions. The results indicated the effectiveness of CR in different groups independent of the relationship between the rats. No extreme changes in weight were observed in the CR rats. Behavioural observations also indicated the differences in the total time spent under the feeder and in the number of pushes (higher in both cases for the sibling rats). However, no significant differences in corticosterone levels were observed. Our results suggest the viability of group-housing rats during long periods of CR maintenance.
The effects of caloric restriction (CR) on health and behaviour have been extensively studied. 1 For example, CR can increase life expectancy,2–5 result in cognitive function decrement after chronic CR, 6 modify biological parameters, 7 enhance or decrease drug effects,8,9 induce anxiolytic effects,10–13 and induce hyperactivity.14,15 Typically, CR experiments require extended time periods. Weight control and the potential for aggressive behaviour avoidance16,17 can force single-housing of the animals, which can result in many logistical issues (such as more cages and racks, more space for placement, and more personnel for monitoring experiments, etc.) as well as problems with animal well-being such as stress. 18 Legislation application (for a review see 19 ) reduces installations and resources, and can be used for the individual maintenance of animals during long periods of time. Moreover, single-housing negatively impacts animal well-being (for a review see 20 ), which could be a potential confounding variable in future protocols applied to an individualized animal. This finding is especially observed in aging animal experiments, where several years of CR maintenance are necessary. Unlike the potentially harmful behaviours associated with diet restriction, other studies have indicated an increase in social behaviour associated with CR. 21 Similarly, previous studies, where food was administrated using a diet board method (food pellets tightly embedded in grooves cut into the aspen board) in grouped male rats, showed an effective weight gain decrease and lower fatty acid, triglyceride and cholesterol levels, without affecting behavioural parameters usually affected by CR as the diurnal activity or other biological markers such as total protein, albumin and alanine aminotransferase in male rats,22,23 but not in female rats. 24 Using the diet board method, a higher corticosterone response was recorded but without observing any pathology associated with this reaction. 25 However, a majority of behavioural studies in diet restriction has only included individualized animals.
In the present study, the main objective was to compare how a 70% CR affect weight, eating and relationship behaviours of sibling and non-sibling grouped rats. Another objective was related to the corticosterone used as an indicator of stress and its relation to animal welfare. 26 In addition to the weight and behavioural effects, a further objective was to study the stress response in sibling and non-sibling grouped rats from the serum corticosterone level analysis.
Materials and methods
Ethical approval and other ethical considerations
Animals were maintained according to the EU Directive 2010/63/EU and the Spanish Royal Decree 53/2013. The University of Granada Research Ethics Committee approved the experimental protocol (reference number 127-CEEA-OH-2015). All animal procedures carried out in this study were subject to review by the animal welfare responsible and designated veterinarian of the Animal Facility of Centro de Investigación Biomédica de la Universidad de Granada (CIBM/UGR).
Animals
Thirty-three male Wistar rats (Rattus norvegicus) from CIBM/UGR were used in this study. The rats were divided into groups of three and kept in transparent methacrylate cages (35 × 50 × 20 cm) (Tecniplast, Buguggiate, Italy) in rooms at 22 ± 1℃ and with a 12:12 h light/dark cycle (lights on at 07:30 h). Siblings and non-siblings were housed together 6–7 weeks after their birth (22 days after weaning). At the beginning of the experiment, the average weight was 155.7 ± 22.05 g. Over the 23-day experiment, water was accessible ad libitum and food was administered as described in the procedure below.
Procedure
The same weight proportions and living conditions were maintained in every cage in the restriction groups and in the ad libitum group. To control the effectiveness of the restriction process, 12 non-related rats (Group Ad Libitum, housed in groups of three rats per cage) were designated as the control group. Twenty-one rats were given a 70% alimentary restriction. Three rats were grouped in each cage and were distributed into two groups. In the first group, 12 non-sibling rats (Group Restricted–Non-Related) were given a 70% alimentary restriction and the second group of nine sibling rats (Group Restricted–Related) was also given a 70% alimentary restriction. The tail of each rat from each cage was marked with a different colour (red, blue or unmarked) to differentiate the rats.
Weight registration and behavioural observation
Every day at 11:00 h, each rat was weighed with a bascule (Model TP12, Denver Instruments, NY, USA) and the food was (Harlan, Mucedola, Italy) dispensed. For the Ad Libitum group 200 g of pellets were given, while the groups on the alimentary restriction were given 70% of the food that the Ad Libitum group rats ate each day (any wasted food that the Ad Libitum group rats left was weighed). The food was dispensed randomly to each cage so the time that the rats had to wait before they could eat (and their resultant added stress) would be the same under every condition. After calculation of the mean and standard deviation from the registered weight, the proportion of the weights of the three animals that were in each cage was calculated.
The proportion of weights in each cage was calculated using the weight of the rat which was highest as 100%. Using the rule of three, the other two percentages were calculated and the average in each cage was obtained. If the proportions were more extreme, the average value would be lower (much less than 100). However, if the proportions were more equal, the average value would be higher (close to 100).
The animals were observed and recorded every day using a digital Sony HD video camera for 5 min in each cage immediately after food was provided in both groups under the CR conditions. After the experimental procedure ended, the social behaviours of the animals in the recorded videos were analysed. Given that only two behaviours were observed, the analyses of the behaviours were focused on the time (measured with a timekeeper) that each rat was eating under the feeder and the number of ‘pushes’ that each rat imposed on their cagemates while they were eating. The ‘push’ action is defined as a defence from the other rats with the front or back paws reflecting fighting behaviour or trying to submit (submission response) to the rats that receive it. 27
Hormonal analysis
Fifty microlitres of blood were taken from the tail vein of 16 rats on the 10th day of the experiment. Out of the 16 rats, five were non-related rats on the ad libitum food. Six were non-sibling rats on the 70% alimentary restriction. The last five were sibling rats on the 70% alimentary restriction. From these blood samples, serum was obtained for the hormonal analysis. Hormonal analysis was performed using the Milliplex map rat stress hormone kit (RSHMAG-69K-03) and the Luminex 200™ HTS, FLEXMAP 3D (Austin, Texas, USA). The serum samples were prepared using the following method: Blood was allowed to clot for at least 30 min before centrifugation at 1000 g for 10 min. The serum was removed and assayed immediately. In each serum sample, 150 µL of the antibody beads and assay buffer was included in the mixing bottle to obtain a total volume of 2850 µL. Next, the samples were incubated on a shaker overnight at 4℃. Samples were then collected on the Lumina 200™. Median fluorescent intensity (MFI) was recorded using a weighted five-parameter logistic or spline curve-fitting method to calculate the analyte concentrations in the samples.
Statistical analysis
The statistical analyses were performed with SPSS 22.0 (IBM Corp, Armonk, NY, USA). Weights, proportion of the weights, time eating under the feeder and number of pushes were compared using repeated measures analysis of variance (ANOVA). If the three groups were included in the analysis, interactions were analysed with pairwise Tukey tests derived from the main analysis. The significance level in all the cases was P < 0.05. For the corticosterone statistical analysis, Cohen’s d (http://www.uccs.edu/∼lbecker/) was applied to determine the magnitude of the effect. Values of the Cohen’s d that were equal or under 0.2 indicate a lower magnitude of the effect. Values between 0.5 and 0.7 indicate a mild magnitude of the effect. At the minimum, values over 0.8 indicate a greater magnitude of the effect. 28
Results
Figure 1 shows the weights of the three groups of rats (Ad Libitum, Restricted–Non-Related and Restricted–Related). Repeated ANOVA (3 × 23) showed a significant effect of the interaction. Analysis of the interaction showed a significant increase in body weight in the three groups (Ad Libitum: F = 400.8; P < 0.001. Restricted–Non-Related: F =148.216; P < 0.001. Restricted–Related: F = 106.572; P < 0.001). Comparison of the three groups each day resulted in a significantly higher weight in the Ad Libitum group versus the Restricted–Non-Related and Restricted–Related groups (P < 0.02) Restricted–Non-Related and the Restricted–Related groups showed no differences in their weights across days. On the last day, the proportion of weight in the restricted groups was 79% versus the Ad Libitum rats. Analysis of the proportion of the weights showed no differences between the groups. Equally, analysis of the standard deviation as a dependent variable showed no differences between the groups. This finding confirmed that the restricted rats had similar weights.
Mean (±SEM) weights (g) across 23 days (D). Ad Libitum rats (Adl.No-Rel) showed significantly higher weights than Restricted–Non-Related (Res.No-Rel) and Restricted–Related (Res.Rel) rats. *P < 0.05, significantly different Adl.No-Rel versus Res. No-Rel or Res.Rel.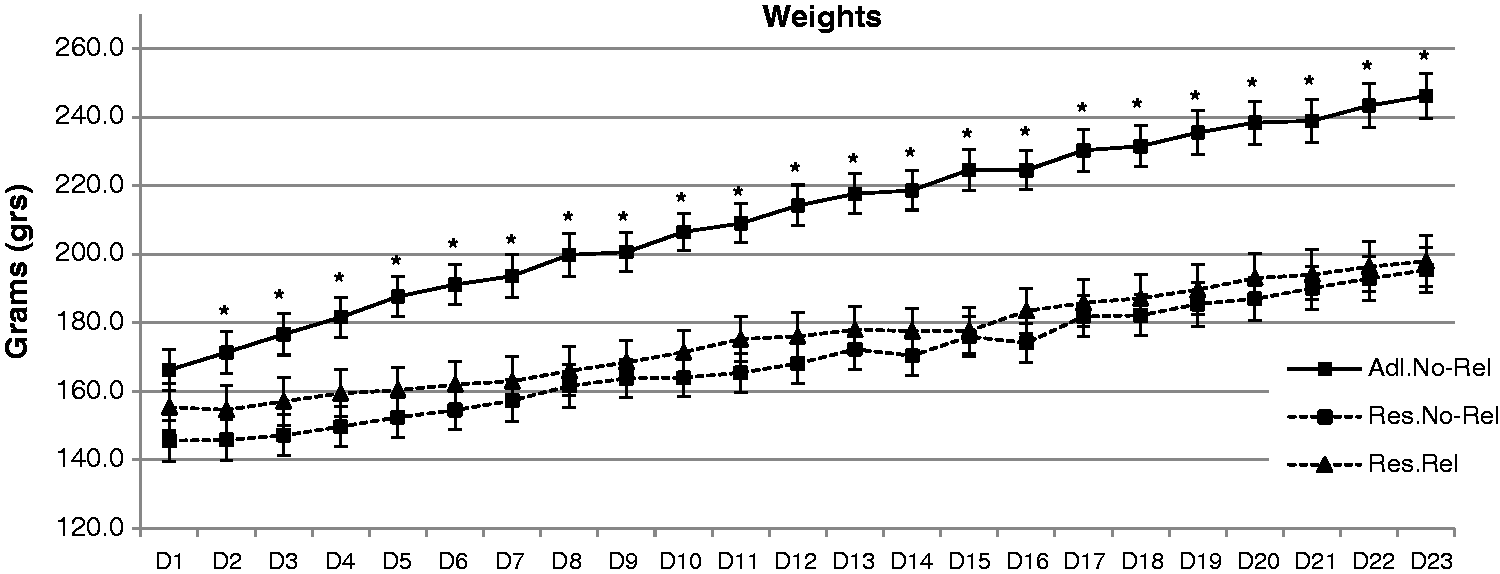
Figure 2 shows the time spent under the feeder for the Restricted–Non-Related and Restricted–Related rats. Ad Libitum rats were excluded, and no significant changes in their behaviour were observed when the food was administered. A visual inspection of the data in each cage confirmed that normally, the same rat spent more time under the feeder than the other two rats. A repeated ANOVA (3 × 23) of the times spent under the feeder showed a significant effect of the interaction (F = 6.478; P < 0.001). Analysis of the interaction showed that the Restricted–Non-Related group spent significantly more time under the feeder versus the Restricted–Related group on day 2 (P < 0.001). Also the Restricted–Related group spent more time under the feeder than the Restricted–Non-Related group on days 17, 18, 20 and 23 (P < 0.03).
Mean (±SEM) times under the feeder (segs) during the 23 days (D). Restricted–Non-Related (Res.No-Rel) rats spent significantly more time under the feeder on the second day. Restricted–Related (Res.Rel) rats spent more time under the feeder on days 17, 18, 20 and 23. *P < 0.05, significantly different Res. No-Rel versus Res.Rel.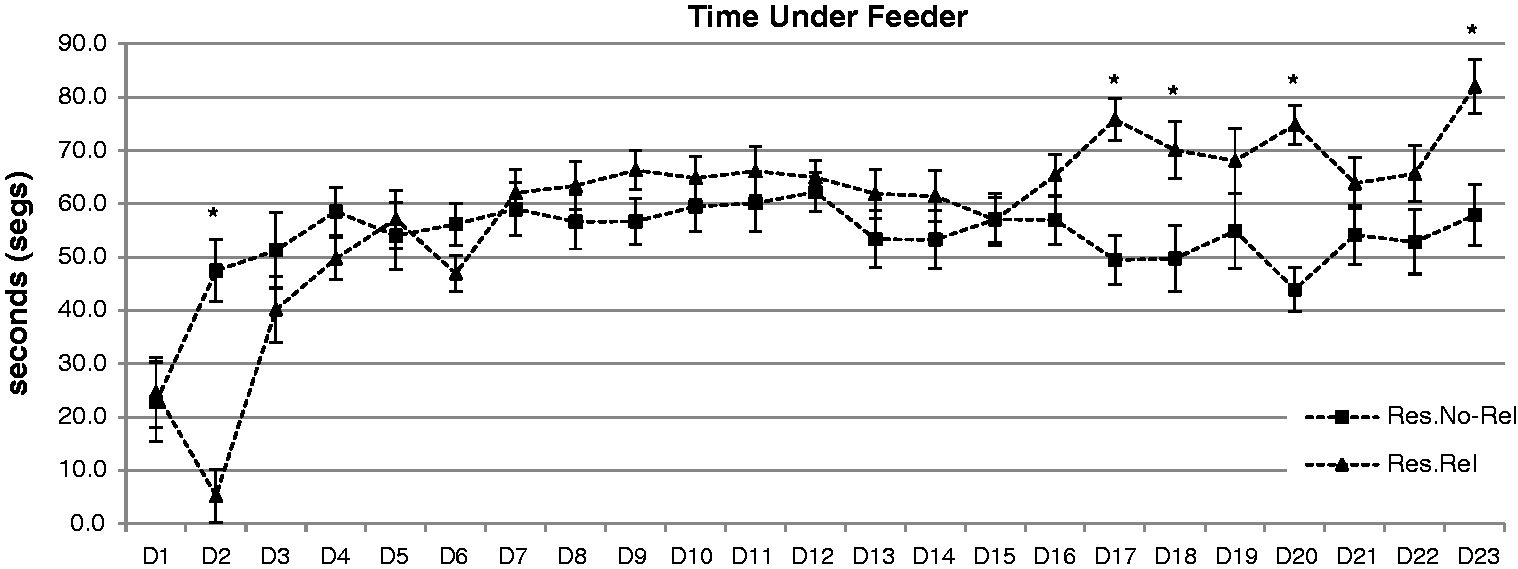
Figure 3 shows the number of pushes for the Restricted–Non-Related and the Restricted–Related rats. Again, Ad Libitum rats were excluded, and no significant changes in their behaviour were observed when the food was administered. A repeated ANOVA (3 × 23) indicated a significant effect of the variable day (F = 3.308; P < 0.01) and the variable group (F = 216.9; P < 0.0001). Thus, independent of the group, differences in the number of pushes were recorded across days. Additionally, the Restricted–Related rats had significantly more pushes than the Restricted–Non-Related rats.
Mean (±SEM) number of pushes during the 23 days (D). Restricted–Related (Res.Rel) rats showed a higher number of pushes than Restricted–Non-Related (Res.No-Rel) rats.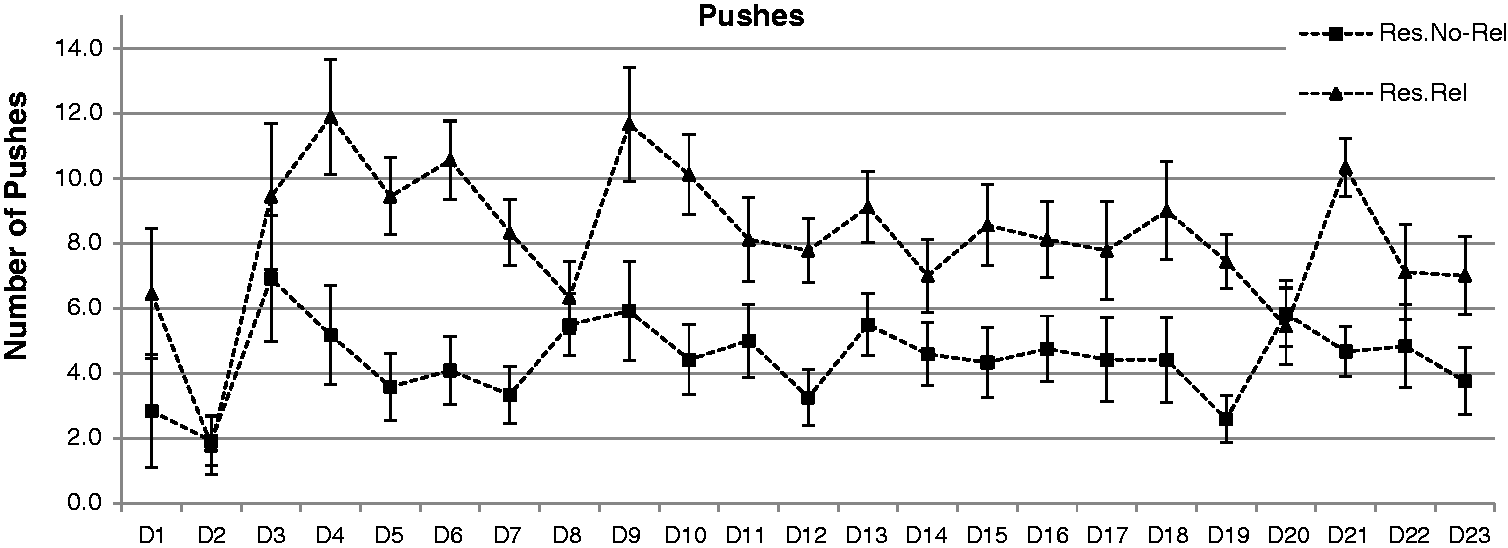
Figure 4 shows the corticosterone levels obtained for the three groups. A unifactorial ANOVA showed no significant differences between the groups. However, Cohen’s d was used due to the reduced number of cases in each group (only five samples). Comparisons between all of the groups showed a significant magnitude of effect between the Restricted–Related rats and the Ad Libitum non-related rats (d = 1.54), as well as between the Restricted–Related rats and the Restricted–Non-Related rats (d = 1.2). Ad Libitum non-related and Restricted–Non-Related groups had a lower magnitude of the effect (d = 0.28).
Median (±SD) of corticosterone concentration in Ad Libitum non-related (Adl.No-Rel), Restricted–Non-Related (Res.No-Rel) and Restricted–Related (Res.Rel) rats.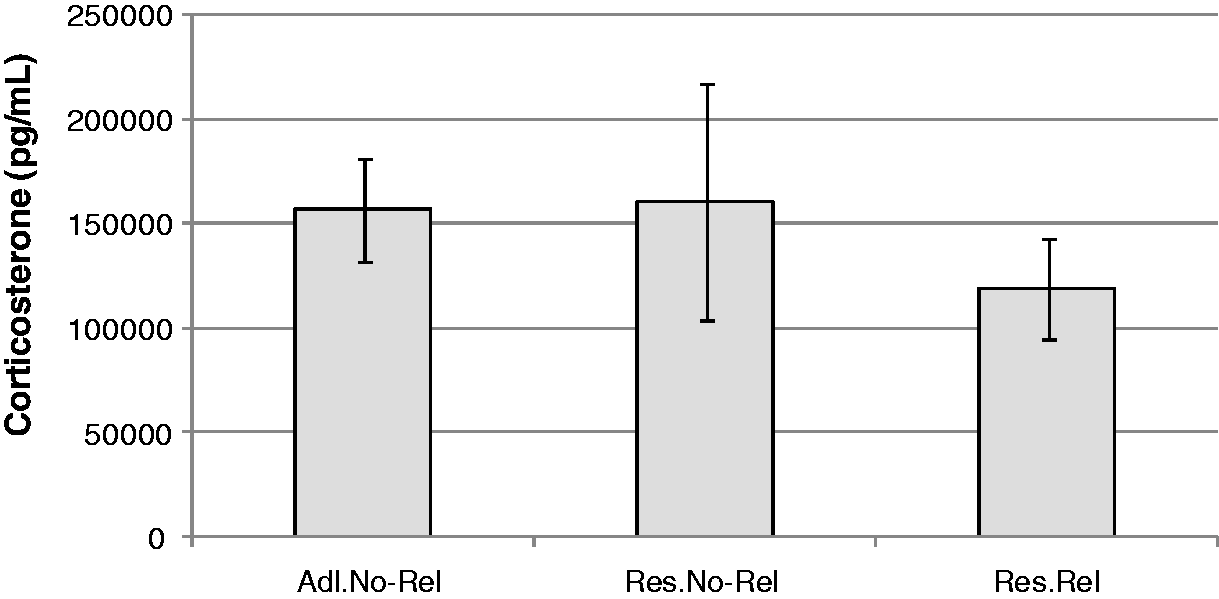
Discussion
The results confirm that the effects of CR were associated with housing rats in groups of three, independent of sibling or non-sibling relationship. Both restricted groups displayed a significant reduction in their weights of 79%. Similar procedures of restriction used in individualized rats have shown a weight reduction of 77% 29 or 76% 30 at 10 weeks, which is very similar to the reduction obtained after the group housing of the rats in this study. However, the small increase in the weight proportion associated with the housing is congruent with other studies that have demonstrated less weight loss associated with CR in grouped housing. 31 The absence of differences in weight proportions confirms that similar weights were observed in each cage independent of sibling presence in the restricted rats. These data are especially relevant when the time spent under the feeder is being analysed. A visual inspection of the data in each cage confirms that normally, the same rat passed under the feeder more than the other two. This finding indicates that all rats had at least the same amount of food. This observation is similar to other studies where advanced social behaviour cooperation has been demonstrated in rats.32–34 In our study, advanced social behaviours could have been manifested in the rats, which displayed an equitable distribution of food among the three rats in each cage. This advanced social behaviour cooperation is inherent in these rats and might explain and enable CR in grouped rats.
The results obtained in the time spent under the feeder can equally be interpreted as another example of social behaviour cooperation. The longer time spent under the feeder observed in the Restricted–Related rats could reflect a higher social behaviour of cooperation. With this behaviour, related rats would be facilitating the access to the food of another rat. From our point of view, these pushes do not reflect aggressive/dominant behaviour (considering that no other aggressive behaviour such as bites, screams or similar were recorded). In this context, pushes would indicate a consequence of the social interaction established among the rats. If we considered that the Restricted–Related rats spent more time under the feeder, it would be normal that these rats would display a higher number of social interactions (pushes). This interpretation is in accordance with the corticosterone data. The absence of differences (even the lower levels of corticosterone obtained in the Restricted–Related rats) between the different groups is indicative of low levels of stress and the absence of aggressive behaviour. There was also a similar level of corticosterone between the restricted and Ad Libitum rats, which confirms the non-stressful effects of CR in grouped rats. Surprisingly, the group restriction prevented the increase in corticosterone levels typically associated with CR. 1 These data open up future research where single and grouped rats may be compared in their wellness using corticosterone levels, for example, as an indicator of this. Additionally, according to the corticosterone data, the best condition for CR in grouped rats would be with related rats. However, these data should be considered with caution, as only a small number of samples were analysed. In conclusion, the results from this study suggest the viability and the suitability of group housing rats for at least up to a month of CR maintenance.
Footnotes
Acknowledgments
Animals were maintained according to the EU Directive 2010/63/EU for animal experiments. The University of Granada Research Ethics Committee approved the experimental protocol. We acknowledge the technical assistance of Ms Ana Fernández Ibánez, who helped with the corticosterone analysis.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was partially funded by the Master’s program in Applied Ethology and Animal Behaviour, University of Pablo Olavide (Sevilla, Spain) and grant PSI-2013-44945-P from the Ministry of Economy and Competitiveness, Spain.