
Abstract
We set out to determine the expression profiles of glycoproteins possessing N-acetyllactosamine, a precursor carbohydrate of sialyl Lex, during colorectal cancer development. We immunohistochemically analyzed the distribution of N-acetyllactosamine as well as of β4GalT-I, a member of the β1,4-galactosyltransferase family responsible for N-acetyllactosamine biosynthesis, in normal mucosa and in adenoma and carcinoma of the human colorectum. Using monoclonal antibody H11, N-acetyllactosamine was barely detectable in the normal mucosa. In low-grade adenoma, however, N-acetyllactosamine was weakly but definitely expressed on the cell surface, and its expression level was moderately increased in high-grade adenoma and markedly increased in carcinoma in situ as well as in advanced carcinoma. To detect β4GalT-I, we used a newly developed polyclonal antibody (designated A18G), which is specific for the stem region of human β4GalT-I. Faint expression of β4GalT-I was detectable in normal mucosa, and the expression level was moderately increased in low-grade adenoma and in high-grade adenoma and markedly increased in carcinoma in situ and advanced carcinoma. The expression of N-acetyllactosamine was highly correlated with the expression of β4GalT-I in these tumor cells. These results indicate that the expression level of β4GalT-I is apparently enhanced during tumorigenesis in the colorectum and that β4GalT-I mostly directs the carcinoma-associated expression of N-acetyllactosamine on the colorectal tumor cell surface.
N-acetyllactosamine (Galβ1 → 4GlcNAcβ → R) is a precursor carbohydrate of sialyl Lex (Figure 1A) (Feizi 1985; Fukuda 1996; Hakomori 1996). Therefore, it is possible that this particular carbohydrate plays a role in tumor progression. The serum level of paragloboside, a glycolipid possessing N-acetyllactosamine, is significantly elevated in various types of cancer patients, including those with colorectal cancer (Myoga et al. 1988). β1, 4-Galactosyltransferase (β4GalT) is the glycosyltransferase responsible for biosynthesizing N-acetyllactosamine by transferring Gal from uridine 5′-diphosphate-Gal (UDP-Gal) to the terminal GlcNAc of acceptor sugars in glycoproteins and/or glycolipids with a β1,4-linkage (Figure 1A). cDNAs of the first type of β4GalT, now designated β4GalT-I, have been cloned from bovine and human milk (Narimatsu et al. 1986; Shaper et al. 1986; Masri et al. 1988). For over 10 years it was believed that β4GalT-I was the only enzyme to form N-acetyllactosamine. Recently, however, five additional members of the human β4GalT family, which show significant homology with β4GalT-I in terms of amino acid sequence levels, have been successively cloned by a number of independent laboratories (Almeida et al. 1997; Lo et al. 1998; Sato et al. 1998; Schwientek et al. 1998). Previously, we showed that N-acetyllactosamine is expressed in an oncodevelopmental manner in the colorectum (Nakayama et al. 1992), but it has not been determined which type of β4GalT might actually direct the tumor-associated expression of N-acetyllactosamine.
In this study, we immunohistochemically analyzed the expression pattern of N-acetyllactosamine during the adenoma-carcinoma sequence in the colorectum to determine whether the expression of N-acetyllactosamine actually is enhanced during colorectal cancer development. In parallel, the expression of β4GalT-I was also evaluated using a newly developed polyclonal antibody, designated A18G, which is specific for β4GalT-I. The results presented here indicate that the expression level of β4GalT-I is apparently enhanced during tumorigenesis in the colorectum and that β4GalT-I mostly directs the carcinoma-associated expression of N-acetyllactosamine on the colorectal tumor cell surface.
Materials and Methods
Production of Polyclonal Antibody Against Human β 4GalT-I
The monospecific polyclonal antibody A18G, directed to the stem region of human β4GalT-I, was generated against a synthetic peptide, ASSQPRPGGDSSPVVDSG, corresponding to the stem region (amino acids 89–106) of human β4GalT-I (Masri et al. 1988). The peptide was conjugated with bovine serum albumin using glutaraldehyde, then immunized in rabbits. Intradermal injections were given every 3 weeks, using 1 mg of the antigen in complete Freund's adjuvant for the primary injection and 0.5 mg for the following two injections. The polyclonal serum was purified using an affinity column prepared with the antigen coupled to CNBr-activated beads (Pharmacia Biotech; Uppsala, Sweden) according to the manufacturer's instructions. For detection of N-acetyllactosamine, the mouse monoclonal antibody H11 was used. The establishment and characterization of this antibody has been described elsewhere (Myoga et al. 1988).
Construction of Mammalian Expression Vectors Harboring Human β 4GalT-II, -III, -V, or -VI cDNA
It is shown that transcripts for β4GalT-II, -III, -V, and -VI are expressed in human fetal brain (Lo et al. 1998). Therefore, we amplified full-length cDNAs encoding β4GalT-II (nucleotides −26 to +1149; the first nucleotide of the initiation codon is defined as +1), β4GalT-III (nucleotides −29 to +1214), βGalT-V (nucleotides −27 to +1197), and βGalT-VI (nucleotides −28 to +1178) by polymerase chain reaction (PCR) using a human fetal brain cDNA library (Invitrogen; Carlsbad, CA) as template. On the basis of published sequences of the β4GalTs (Almeida et al. 1997; Lo et al. 1998; Sato et al. 1998), specific primer sets used for the PCR were designed as follows: β4GalT-II, 5′-CCCAAGCTTGGGCGGGCCAGTGGC-3′ and 5′-GCTCTAGAGGCACCGAGAGCCTCTG-3′; βGalT-III, 5′-CCCAAGCTTCCTCTCCCTCCCTAGATCT-3′ and 5′-GC TCTAGAGTTTCATGATTAAGGTAGACAG-3′; β4GalT-V, 5′-CCC AAGCTTGCTCTCTAGCGGCCGCC-3′ and 5′GC TCTAGAGTGGGTAAAGCAAACGTACA-3′; and β4GalT-VI, 5′CCC AAGCTTGCCTGGGCTGCTGGCAG-3′ and 5′-GCTCTAGAGGTCTACCTTGCCACGACA-3′. The HindIII and XbaI sites are indicated in italics. After the digestion with HindIII and XbaI, these cDNAs were introduced into the HindIII and XbaI sites of a mammalian expression vector, pcDNAI (Invitrogen), resulting in pcDNAI-β4GalT-II, pcDNAI-β4GalT-III, pcDNAI-β4GalT-V, and pcDNAI-β4GalT-VI.
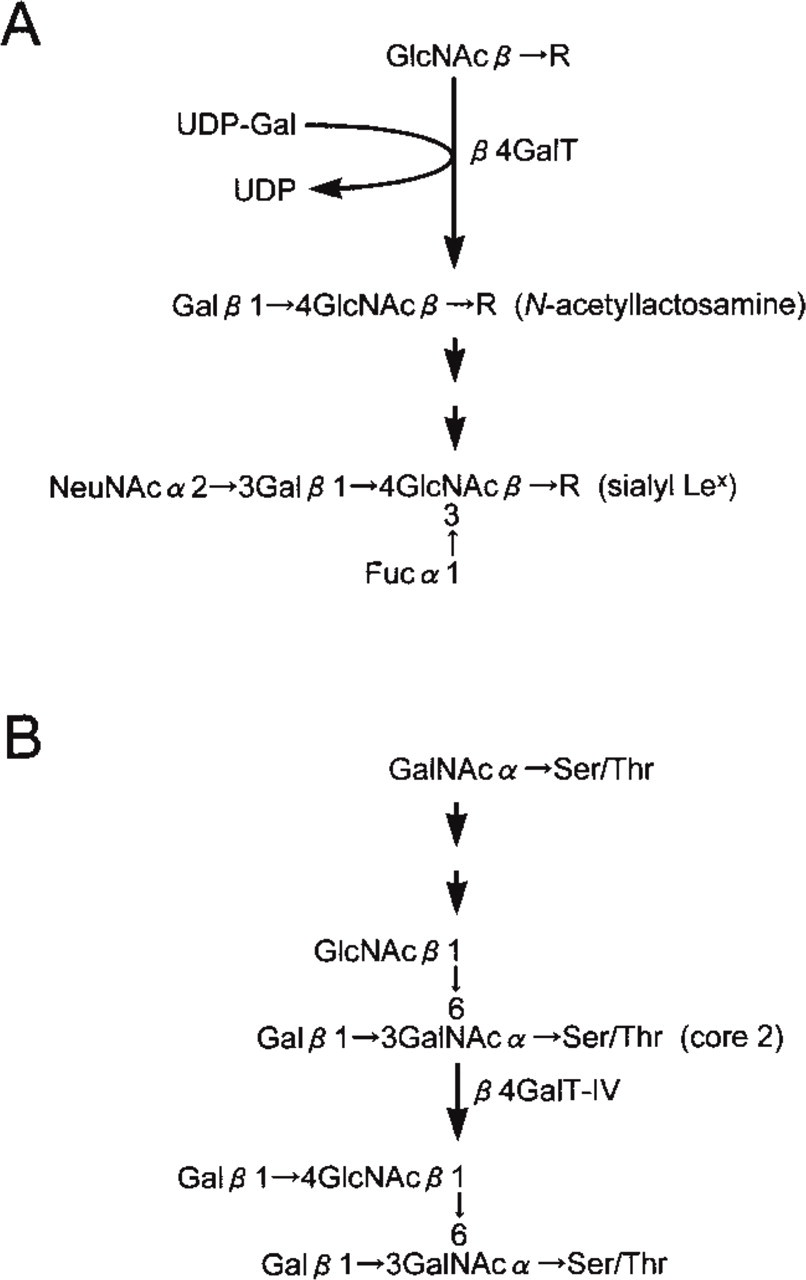
Synthetic pathways of oligosaccharides catalyzed by β1,4-galactosyltransferase (β4GalT). (A) Synthetic pathway of N-acetyllactosamine and sialyl Lex. β4GalT forms N-acetyllactosamine (Galβ1 → GlcNAcβ → R) by transferring Gal from uridine 5′-diphophate-Gal (UDP-Gal) to GlcNAcβ → R with a β1,4-linkage. N-Acetyllactosamine is specifically detected by the monoclonal antibody H11. (B) Biosynthetic pathway of O-glycans having core two-branched oligosaccharide, GlcNAcβ1 rR 6(Galβ1 → 3)GalNAc. Among the members of β4GalT family, β4GalT-IV is involved in the biosynthesis of N-acetyllactosamine attached to core two-branched oligosaccharide.
Transient Expression of Human β4GalTs in Transfected COS-1 Cells
Two micrograms of pCAG-GT encoding a cDNA of human β4GalT-I (Aoki et al. 1992) was transfected into COS-1 cells grown on glass coverslips in a Falcon 3005 tissue culture dish (Becton Dickinson; Franklin Lakes, NJ) using 20 μl of LipofectAMINE (Life Technologies; Gaithersburg, MD), as described previously (Nakayama et al. 1995). Similarly, 2 μg of pcDNAI-β4GalT-II, pcDNAI-β4GalT-III, pcDNAI-β4GalT-V, and pcDNAI-β4GalT-VI was separately transfected into COS-1 cells. In addition, 2 μg of pcDNA3-β4GalT-IV (Ujita et al. 1998), kindly provided by Dr. Jiuun Yeh-Chen (The Burnham Institute; La Jolla, CA) was also transfected into COS-I cells. Some 48 hr after the transfection, COS-1 cells were fixed with 4% paraformaldehyde in PBS, (pH 7.4) (Aoki et al. 1992). After washing with PBS, the COS-1 cells were permeabilized with 0.2% saponin and incubated with the A18G antibody. As the second antibody, fluorescein isothiocyanate-conjugated anti-rabbit IgG (Immunotech; Marseille, France) containing 0.2% saponin was used. For the cell surface labeling of the antibody, saponin was omitted from the procedure.
In parallel, COS-1 cells were transfected by pCAG-GT and then subjected to Western blot analysis (Towbin et al. 1979) using A18G antibody or another polyclonal antibody specific for β4GalT-I, which was established by immunizing a rabbit with the soluble form of β4GalT-I purified from human milk (Yamaguchi and Fukuda 1995). COS-1 cells (5.0 × 106) transfected by 20 μg of pCAG-GT and the nontransfected parental COS-1 cells (5.0 × 106) were lysed in PBS containing 1% Nonidet P-40 and a protease inhibitor cocktail, Complete (Boehringer Mannheim; Mannheim, Germany) on ice for 45 min. Then these samples were separated by sodium dodecyl sulfate (SDS)-polyacrylamide gel (10%) electrophoresis (PAGE) in the presence of β-mercaptomethanol and then transferred onto nitrocellulose membrane. The membrane was incubated with A18G antibody or the other β4GalT-I-specific polyclonal antibody, followed by horseradish peroxidase (HRP)-conjugated anti-rabbit immunoglobulins. The peroxidase activity was visualized using ECL Western blotting detection reagents (Amersham Pharmacia Biotech; Piscataway, NJ).
Colorectal Tumor Specimens and Tissue Preparation
Twenty-one polypectomies for colorectal polyp and 17 surgically resected specimens of advanced colorectal adenocarcinoma were selected from the pathology files of Central Clinical Laboratories (Shinshu University Hospital, Matsumoto, Japan). Histopathological examination was performed based on the criteria of the WHO classification (Jass and Sobin 1989), and it was found that 12 of 21 polypectomies consisted of more than two different degrees of dysplasia. In addition, tubular adenomas with low-grade or high-grade dysplasia were found around three cases of advanced carcinoma. Immunohistochemical results were evaluated for every dysplastic lesion, and we eventually examined 19 lesions of tubular adenoma with low-grade dysplasia (low-grade adenoma), 15 lesions of tubular adenoma with high-grade dysplasia (high-grade adenoma), 10 lesions of carcinoma in situ, and 17 lesions of advanced carcinoma (Table 1). We also examined normal-appearing mucosa found in a cut end of polypectomies or colorectomies as the normal mucosa. Transitional mucosa adjacent to the cancer was excluded from the evaluation because a distinctive pattern of glycosylation is noted in the transitional mucosa (Nakayama et al. 1987). All tissues were fixed for 24 hr in 20% buffered formalin, pH 7.4, then embedded in paraffin and sectioned serially at 3-μm thickness for subsequent immunohistochemical staining or hematoxylin and eosin staining.
Immunohistochemistry
Immunohistochemical staining for N-acetyllactosamine or β4GalT-I was performed on consecutive sections by an indirect immunoperoxidase method, as described (Nakane 1975). Because preliminary experiments had indicated that antigen retrieval was required for immunostaining with the A18G antibody, the tissue sections used for A18G were subjected to microwave irradiation for 10 min in 10 mM citrate buffer, pH 6.0, before incubation with the antibody (Shi et al. 1991). After blocking of endogenous peroxidase activity with 0.3% hydrogen peroxide, the serial tissue sections were incubated overnight at 4C with H11 antibody or purified A18G antibody. After rinsing with PBS, the tissue sections were incubated with HRP-conjugated anti-mouse immuno-globulins (DAKO; Glostrup, Denmark) for H11 antibody or anti-rabbit immunoglobulins (DAKO) for A18G antibody. After rinsing, the peroxidase activity was visualized with the aid of a diaminobenzidine/hydrogen peroxide solution. Counterstaining was carried out with hematoxylin. In control experiments performed by omitting the primary antibody, no specific staining was found.
Evaluation
To evaluate the results of the immunohistochemical staining for N-acetyllactosamine and β4GalT-I, a scoring system based on the criteria laid down by Basolo et al. (1996) was used, with minor modifications. Briefly, the staining intenity was scored from 0 to 3 (0, absent; 1, weak; 2, moderate; 3, strong). In addition, the proportion of antigen-positive cells to all epithelial cells in the tissue specimen was graded from 0 to 3 (0, < 5%; 1, 6–10%; 2, 11–50%; 3, >51%). By adding the two individual scores, the total score was determined. These observations were done by scanning the entire tissue specimens at a low-power magnification (X40 or X100) and then confirmed under a high-power magnification (X 400).
Expression of

Results
A18G Antibody Specifically Detects Human β4GalT-I Expressed in Transfected COS-1 Cells
To test the specificity of the A18G antibody, we transfected COS-1 cells with human βMGalT-I cDNA (pCAG-GT), then immunostained the transfected cells with A18G. Before the transfection, the permeabilized COS-1 cells were not stained with A18G antibody (Figure 2A). By contrast, after COS-1 cells were transfected with pCAG-GT, A18G antibody reacted strongly with the Golgi region of permeabilized COS-1 cells (Figure 2B) or with the cell surface of unpermeabilized COS-1 cells (Figure 2C). On the other hand, A18G did not react with the transfected COS-1 cells expressing β4GalT-II (Figure 2D), β4GalT-III (Figure 2E), β4GalT-IV (Figure 2F), β4GalT-V (Figure 2G), or β4GalT-VI (Figure 2H), irrespective of the permeabilization of the COS-1 cells.
In addition, Western blot analysis revealed that specific bands detected by A18G antibody were found in cell lysates derived from the COS-1 cells transiently transfected by pCAG-GT approximately at 62, 52, 49, and 46 kD (arrows in Figure 3, Lane 1) as well as 122 kD (arrowhead in Figure 3, Lane 1). Moreover, these proteins were also recognized by the other polyclonal antibody raised against the soluble form of β4GalT-I (Figure 3, Lane 2). By contrast, these bands were not detectable in the parental COS-1 cells by A18G antibody (Figure 3, Lane 3). These results combined indicate that all the multiple bands found in the transfected COS-1 cells represent β4GalT-I, because they were specifically detected by the two different kinds of antibodies specific for β4GalT-I. Yamaguchi and Fukuda (1995) demonstrate that β4GalT-I forms homodimers resistant to SDS and β-mercaptoethanol between the membrane-spanning domains of these molecules to be retained in the Golgi apparatus. Thus, the proteins reactive with A18G antibody indicated by arrows or arrowhead in Figure 3, Lane 1 probably represent monomers or a dimer of β4GalT-I, respectively, as described (Yamaguchi and Fukuda 1995). The variations of molecular masses found within the monomers of β4GalT-I, might result from the difference in glycosylation of β4GalT-I, as suggested previously (Yamaguchi and Fukuda 1995).
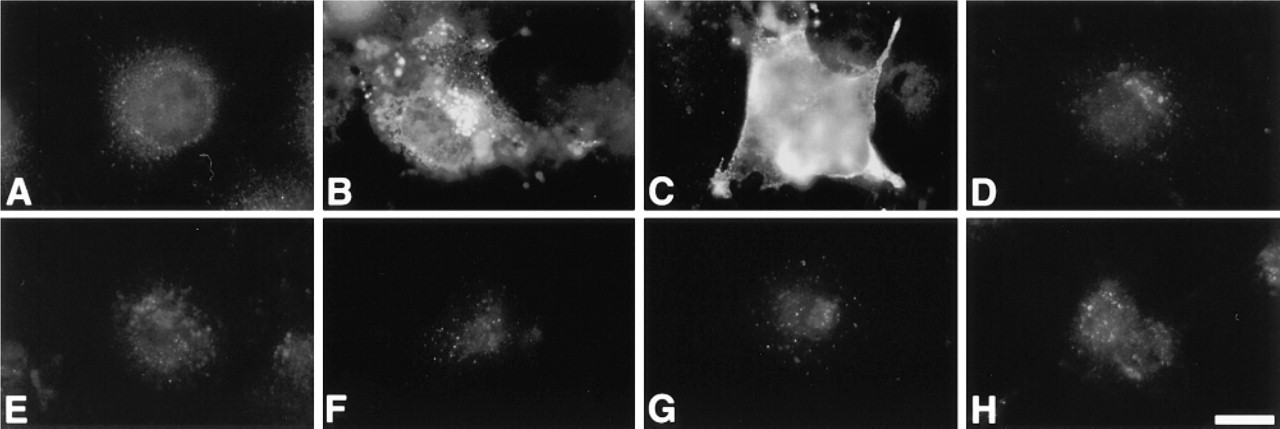
Specific detection of human β4GalT-I expressed in the transfected COS-1 cells by A18G antibody using an immunocytochemical technique. (A) A parental COS-1 cell permeabilized with saponin is not stained by A18G antibody specific for β4GalT-I. After transfection with pCAG-GT encoding human β4GalT-I, A18G antibody reacts with the Golgi region of a permeabilized COS-1 cell (B) or the cell surface of an unpermeabilized COS-1 cell (C). In contrast, COS-1 cells transfected by pcDNAI-β4GalT-II (D), pcDNAI-β4GalT-III (E), pcDNA3-β4GalT-IV (F), pcDNAI-β4GalT-V (G), or pcDNAI-β4GalT-VI (H) were not stained with A18G antibody. Immunostaining with A18G antibody was carried out with (A,B,D-H) or without saponin treatment (C). Bar = 20 μm.
These results have established that the A18G antibody used in this study specifically recognizes the human β4GalT-I but not other β4GalTs (i.e., β4GalT-II to β4GalT-VI). In addition, A18G does not react with the monkey β4GalT-I that is endogenously expressed in COS-1 cells, which are derived from African green monkey kidney cells.
Expression of N-acetyllactosamine Is Enhanced During Colorectal Tumorigenesis
The expression profile of N-acetyllactosamine in normal mucosa and in adenoma and carcinoma of the colorectum is shown in Table 1. The results showed that N-acetyllactosamine was detectable on the apical surface but not in the Golgi region or other intracellular components of epithelial cells, and its expression level was progressively enhanced during the adenoma-carcinoma sequence in the colorectum. N-acetyllactosamine was barely detectable in normal mucosa (Figure 4A). In adenoma, N-acetyllactosamine was weakly expressed along the apical surface of low-grade adenoma cells (not shown) and moderately on the luminal surface of high-grade adenoma cells (Figure 4E). In carcinoma, N-acetyllactosamine was strongly expressed on the apical surface of carcinoma cells in carcinoma in situ (Figure 4G) and in advanced carcinoma (Figure 4I), irrespective of the histological differentiation of the adenocarcinoma cells.
Expression of β4GalT-I Is Correlated with That of N-acetyllactosamine During the Adenoma-Carcinoma Sequence in Colorectal Tumors
The general trend followed by the β4GalT-I expression in normal mucosa, adenoma, and carcinoma in the colorectum is shown in Table 1. In general, β4GalT-I was detectable in the Golgi region of epithelial cells but not on the cell surface, and its expression level was progressively enhanced during the transition from normal mucosa to adenoma and from adenoma to carcinoma. In normal mucosa, the expression of β4GalT-I was weak (Figure 4B). Moderate expression of β4GalT-I was found in low-grade adenoma (Figure 4D) and also in high-grade adenoma (Figure 4F). By contrast, β4GalT-I was strongly expressed in carcinoma in situ, and the expression level appeared to be stronger than in the adenomas (Figure 4H). In advanced carcinoma, β4GalT-I appeared to be more strongly expressed than in carcinoma in situ, irrespective of the histological differentiation of the adenocarcinoma cells (Figure 4J).
Discussion
In this study we have demonstrated that the expression of N-acetyllactosamine, a precursor carbohydrate of sialyl Lex, is progressively enhanced during development of colorectal cancer. N-Acetyllactosamine is well known as the antigenic determinant for the carbohydrate antigens i [(Galβ1 → 4GlcNAcβ1 → 3)nGalβ1 → 4 GlcNAcβ → R] and I [Galβ1→ 4GlcNAcβ1 → 3(Galβ1 →4 GlcNAcβl → 6)Galβ1 → 4GlcNAcβ → R], which are present in fetal and adult erythrocytes, respectively (Fukuda et al. 1979; Watanabe et al. 1979). The i and I antigens are also called poly- N-acetyllactosamine because they are composed of N-acetyllactosamine repeats. As shown in the present study, the erythrocytes in colorectal tissues were positive for Hll antibody (see Figure 4). It was reported previously that the I antigen is increased in colon cancer tissues (Yang et al. 1994). In the present study, we could not distinguish the i antigen from the I antigen because the H11 antibody used in this study reacts with both (Myoga et al. 1988). However, the expression of N-acetyllactosamine was progressively enhanced during the adenoma-carcinoma sequence in the colorectum. This result prompted us to test if the expression of β4GalT was also enhanced during the development of colorectal cancer, because β4GalT is responsible for the biosynthesis of N-acetyllactosamine.
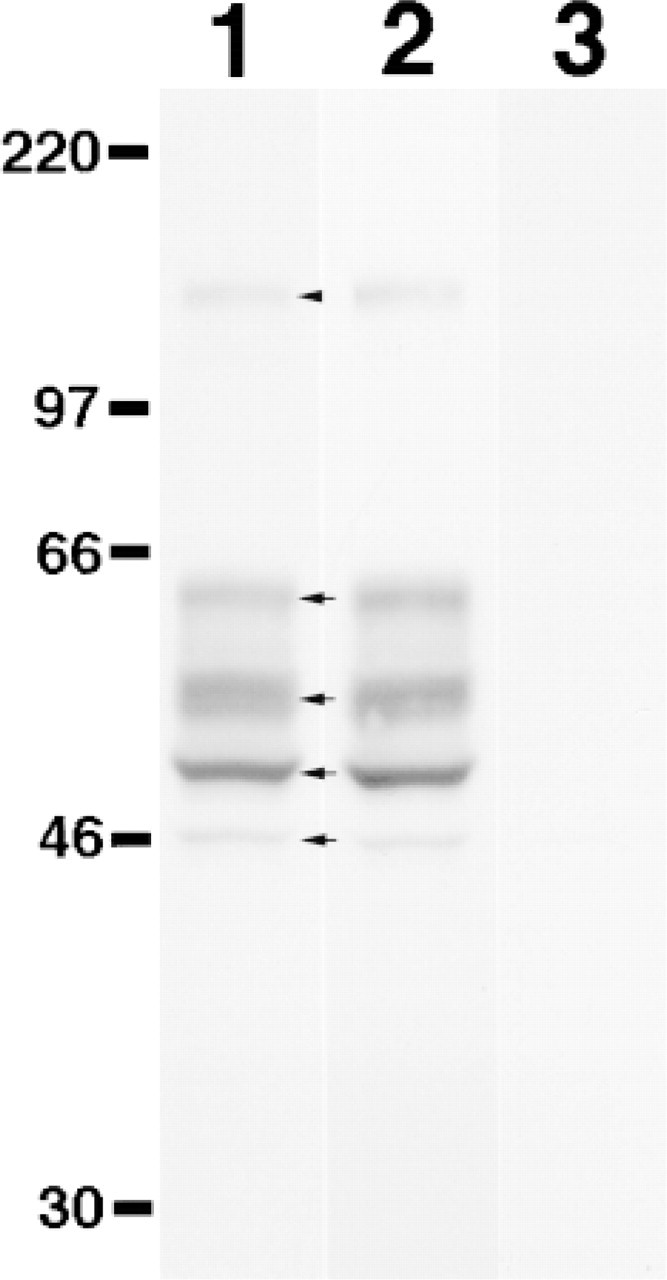
Western blot analysis of the COS-1 cells transfected with human β4GalT-I cDNA. Cell lysates prepared from COS-1 cells (Lanes 1 and 2) transiently transfected with pCAG-GT or the nontransfected parental COS-1 cells (Lane 3) were separated by SDS-PAGE (10% polyacrylamide) under reducing conditions, and then subjected to Western blot analysis using the A18G antibody (Lanes 1 and 3) or another polyclonal antibody raised against the soluble form of β4GalT-I (Lane 2). Arrows and arrowhead in Lane 1 indicate the monomers and dimer of β4GalT-I, respectively. Molecular markers are shown in kD.
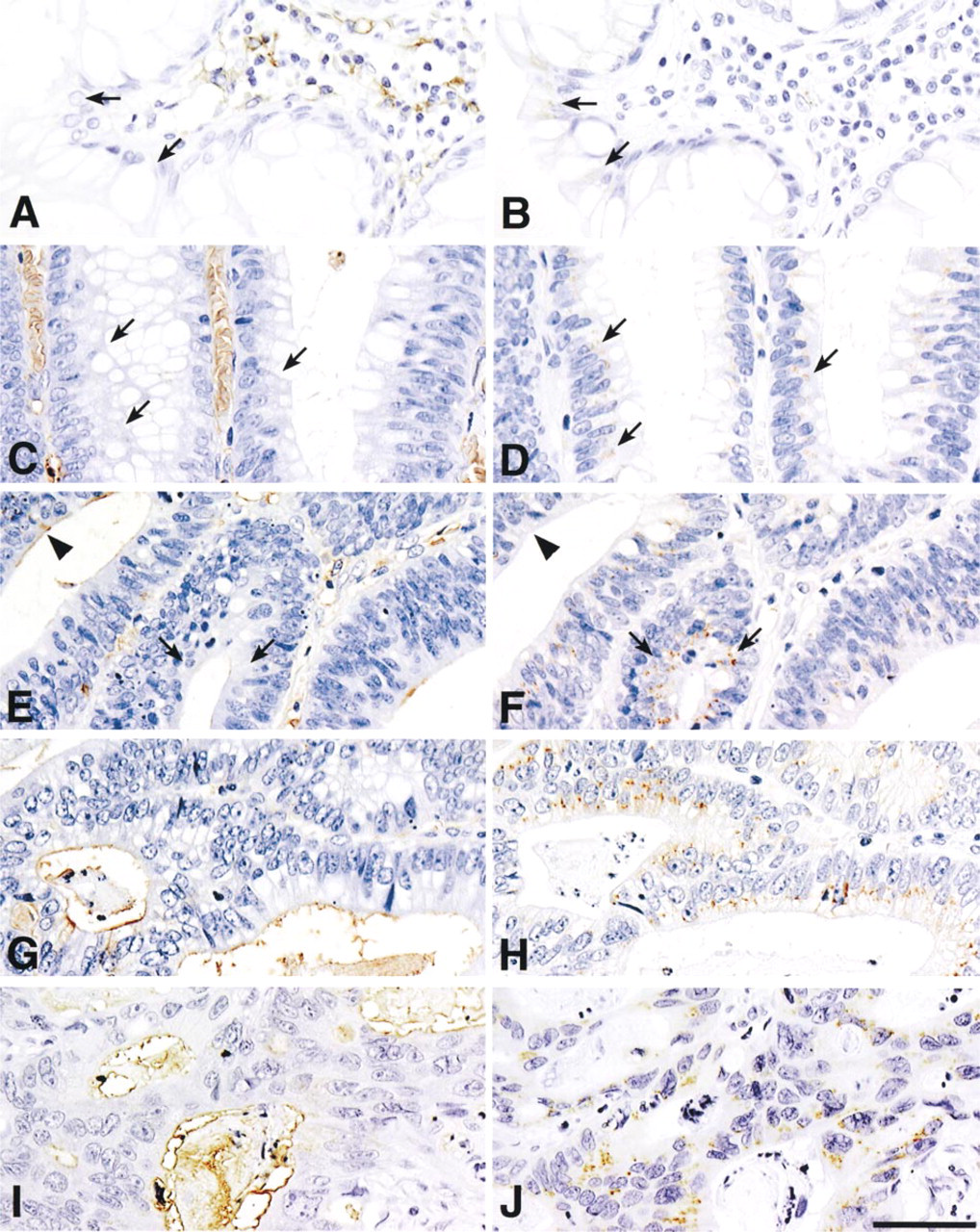
Expression of N-acetyllactosamine (A,C,E,G,I) and β4GalT-I (B,D,F,H,J) in normal mucosa (A,B), low-grade adenoma (C,D), high-grade adenoma (E,F), carcinoma in situ (G,H), and advanced carcinoma (I,J) of the colorectum. Expression of N-acetyllactosamine is progressively enhanced during colorectal cancer tumorigenesis. Expression of β4GalT-I is similarly enhanced during the adenoma-carcinoma sequence. Arrows show epithelial cells expressing β4GalT-I but not N-acetyllactosamine; arrowheads indicate epithelial cells expressing N-acetyllactosamine but not β4GalT-I. Immunostaining was performed with H11 antibody (A,C,E,G,I) or with A18G antibody (B,D,F,H,J). Bar = 50 μm.
Recently, five novel members of the human β4GalT gene family, β4GalT-II to β4GalT-VI, have been cloned by utilizing the expressed-sequence tag database (Almeida et al. 1997; Lo et al. 1998; Sato et al. 1998; Schwientek et al. 1998). The deduced amino acid sequences of the newly cloned β4GalTs reveal that they are typical membrane-bound proteins with Type II topology, composed of a short N-terminal cytoplasmic domain, a membrane-spanning domain, and a short stem region followed by a large catalytic domain, as shown in virtually all the glycosyltransferases cloned thus far (Field and Wainwright 1995). In particular, the catalytic domain is largely conserved in the β4GalT gene family, and the homology of the newly cloned β4GalTs with respect to the first type of β4GalT, β4GalT-I was 55% amino acid identity for β4GalT-II, 50% for β4GalT-III, 41% for β4GalT-IV, 38% for β4GalT-V, and 33% for β4GalT-VI (Lo et al. 1998). Most recent studies, using a variety of synthetic acceptor oligosaccharides and recombinant β4GalTs, have demonstrated that among the members of the β4GalT family, β4GalT-I is the most efficient at forming N-acetyllactosamine as well as poly -N-acetyllactosamine in N-glycans (Ujita et al. 1998). Consequently, we examined the expression pattern of β4GalT-I in the adenoma-carcinoma sequence in the colorectum.
Several authors have carried out immunohistochemical studies in various cells, including HeLa cells and bovine and rat epithelial cells, using polyclonal or monoclonal antibodies against β4GalT-I (Roth and Berger 1982; Suganuma et al. 1991; Taatjes et al. 1992; Kawano et al. 1994). However, because most of them were raised by immunizing animals with the entire molecule of the isolated enzyme as the immunogen, it appeared possible to us that these antibodies might show crossreactivity with other members of the β4GalT family. Therefore, we developed a new polyclonal antibody, A18G, against the stem region of human β4GalT-I, because the stem region of glycosyltransferases generally shows the least homology in the glycosyltransferase family (Paulson and Colley 1989). The stem region of β4GalT-I, which was used as the immunogen for production of A18G, exhibits no homology with those of the other β4GalTs. Using the A18G antibody, we have shown that this particular antibody reacted with β4GalT-I but not with other members of β4GalT family expressed in the transfected COS-1 cells (see Figure 2). In addition, we have also demonstrated that A18G actually reacts with not only the monomers of β4GalT-I but also the dimer of β4GalT-I (see Figure 3). These results have established that the A18G antibody used in the present study is specific for human β4GalT-I.
Using the A18G and H11 antibodies, we next examined the co-expression of β4GalT-I and N-acetyl-lactosamine in serial sections at various stages in the adenoma-carcinoma sequence of colorectal tumors. We found that the expression of β4GalT-I in the carcinoma cells showed a good correlation with that of N-acetyllactosamine (Figures 4G-4J). Previously, we demonstrated that the expression of N-acetyllactosamine in the human colorectum was oncodevelopmentally regulated (Nakayama et al. 1992). It has also been reported that β4GalT activity in the sera is elevated in patients with various types of cancers, including colorectal cancers (Nozawa et al. 1990; Nishiwaki et al. 1992). These results strongly suggest that β4GalT-I participates in the biosynthesis of N-acetyllactosamine, and possibly poly -N-acetyllactosamine, in N-glycans on colorectal cancer cells. However, we cannot exclude the possibility that β4GalTs other than β4GalT-I could be also involved in the biosynthesis of N-acetyllactosamine. In 10 of 15 lesions in high-grade adenoma, β4GalT-I was not detected in some of the adenoma cells expressing N-acetyllactosamine on their cell surface (Figure 4E and 4F, arrowheads), suggesting that β4GalTs other than β4GalT-I can be involved in the biosynthesis of N-acetyllactosamine. Very recently, it has been shown that β4GalT-IV requires core two-branched O-glycans for its preferential acceptor (Ujita et al. 1998) (Figure 1B). Because core two-branched O-glycans are specifically expressed in colorectal cancer cells (Shimodaira et al. 1997), it is possible that β4GalT-IV participates in the carcinoma-associated expression of N-acetyllactosamine in these cancer cells.
It is reported that increased activity of plasma membrane sialidase of the JB6 mouse epidermal cell is associated with transformation induced by phorbol ester (Miyagi et al. 1992). Taking into account in fact that an a2,3- or a2,6-linked sialic acid can be attached to the galactose residue of N-acetyllactosamine by sialyl-transferase, ST3Gal IV, or ST6Gal I (Tsuji 1996), it is also possible that the desialylation of sialyl N-acetyl-lactosamine by action of the sialidase contributed partly to the increased expression of N-acetyllactosamine in these colorectal carcinoma cells, because the H11 antibody does not recognize the sialylated forms of N-acetyllactosamine (Suzuki et al. 1991; Nakayama et al. 1992).
It is also noteworthy that in 13 of 19 lesions in low-grade adenoma, some adenoma cells expressing β4GalT-I were negative for N-acetyllactosamine on the cell surface (Figure 4C and 4D, arrows). In such adenoma cells, it is most likely that N-acetyllactosamine synthesized by β4GalT-I was subsequently sialylated and eventually could not be detected by the H11 antibody.
It has been reported that β4GalT-I is expressed on the cell surface of various cells, including sperm, and it was suggested that this cell surface-associated β4GalT-I could serve as a cell-adhesion molecule by recognizing oligosaccharides either on the cell surface of adjacent cells or in the extracellular matrix, as exemplified in the binding of sperm to the egg coat (Shur 1993). In the present study, a cell surface localization of β4GalT-I was not found in colorectal tumor cells. However, we were able to demonstrate a cell surface expression of β4GalT-I in COS-1 cells transfected by a β4GalT-I cDNA (Figure 2C). This result suggests that the overexpression of β4GalT-I in these transfected cells directs the cell surface expression of this particular enzyme.
In conclusion, we have demonstrated by the use of H11 and A18G antibodies that the expression of β4GalT-I in tumor cells increases progressively during colorectal cancer development and that the enhanced activity of β4GalT-I may, in turn, regulate the cell surface expression on these tumor cells of those carbohydrate antigens having N-acetyllactosamine at their termini. Further studies will be required to determine how the expression of β4GalT-I is regulated during the development of colorectal cancer.
Footnotes
Acknowledgements
Supported by Grants-in-Aid for Scientific Research on Priority Area (10178104) and C (09670222) from the Ministry of Education, Science, Sports and Culture of Japan (to JN), and Grant PO1 CA71932 from the NCI, National Institutes of Health (to MF and MNF).
We are grateful to Dr Tsutomu Katsuyama for encouragement and discussion during this study, to Dr Yukiko Matsumoto and Noriko Kubota for their excellent technical assistance, Dr Jiuun Yeh-Chen for providing pcDNA3-β4GalT-IV, and to Drs Edgar Ong and Mu-Xia Zhang for critical reading of the manuscript.