
Abstract
To study the prevalence of p53 inactivation and MDM2/p21WAFI/CIP1 expression in severe combined immunodeficient (SCID) mice Epstein-Barr virus (EBV)-induced lymphoproliferation, 19 samples obtained after IP injection of peripheral blood mononuclear cells (PBMCs) from EBV-seropositive donors or lymphoblastoid cell lines (LCL) were analyzed. In all samples tested, overexpression of Ki-67 antigen was shown by immunohistochemistry, indicating a high proliferative index of SCID mice EBV-induced lymphoproliferation. P53 mutations were screened by functional assay in yeast in 14 samples. With this test, a p53-inactivating mutation was found in only one case; the remaining cases exhibited a wild-type p53 pattern. However, an accumulation of p53 protein was detected by immunohistochemistry in six of 19 samples. P21 expression was found in seven of 19 samples but was not correlated with the rate of p53 protein in tumors. In contrast, high levels of nuclear accumulation of MDM2 were found in all samples by immunohistochemistry. These results suggest that a high Ki-67 proliferative index in SCID mice EBV-induced lymphoproliferation is not due to the inactivation of p53 by mutation, but could be associated with an overexpression of MDM2, which would act by a p53-independent mechanism.
I
Severe combined immunodeficiency (SCID) mice (Mosier et al. 1988) reconstituted with peripheral blood mononuclear cells (PBMCs) from Epstein-Barr virus (EBV)-seropositive donors or lymphoblastoid cell lines (LCL) rapidly develop lethal solid human B-cell lymphoproliferation. These animals develop EBV-induced lymphoproliferation, that histologically and phenotypically resembles the aggressive lymphoproliferative disease that can arise in immunocompromised individuals (Nakamine et al. 1991; Rowe et al. 1991). Therefore, they constitute a useful tool for study of EBV-induced lymphomagenesis in humans.
Using these EBV-induced lymphoproliferations in SCID mice, studies were performed to assess the cell proliferation index, MDM2 overexpression, and the prevalence of p53 inactivation. P53 mutations and its accumulation were studied using both a functional assay in yeast and immunohistochemistry. Ki-67, MDM2, and p21WAFI/CIP1 expression was simultaneously analyzed by immunohistochemistry.
Materials and Methods
Human Leukocytes and Lymphoblastoid Cell Lines
PBMCs were obtained after informed consent from two EBV-seropositive volunteers (M and T) undergoing lymphapheresis. Prior EBV infection was confirmed by testing for the presence of serum anti-viral capsid antigen (VCA) and anti-Epstein-Barr nuclear antigen IgG. Human PBMCs were prepared by Ficoll-Hypaque gradient centrifugation. EBV-transformed LCLs were prepared in our laboratory by in vitro infection of PBMCs obtained from two volunteer donors, with the EBV B95-8 strain in the presence of cyclosporin A 0.2 μg/ml (Sandoz; Lyon, France). All cell lines were maintained at 37C in RPMI 1640 medium (ICN; Orsay, France) containing 2 mM L-glutamine, 1 mM pyruvate, 50 U/ml penicillin, 50 μg/ml streptomycin, and supplemented with 10% heat-inactivated fetal calf serum (GIBCO BRL; Cergy Pontoise, France). Two lymphoblastoid cell lines were establishe: sa and clo.
Reconstitution of SCID Mice with Human PBMCs or LCLs
CB-17 female scid/scid (SCID) mice were purchased from IFFA-Credo (L'Arbresle, France) and kept under specific pathogen-free conditions. A laminar flow isolator was used during manipulation of the animals, which were housed in groups of up to five in a filter-top cage on sterile bedding and were provided with sterile food and water ad libitum.
For our study, all mice were screened for the “leaky” phenotype, and animals showing more than 50 μg/ml IgG in their serum were excluded. To remove NK activity in vivo, animals were treated for 30 days with rabbit anti-asialo GM1 antiserum (Wako Chemicals, Osaka, Japan; 20 μ1/ mouse). Nine SCID mice (9 weeks old) were injected IP with 80
Histopathology and Immunohistochemical Analysis
Histopathological study was performed on 5
The presence of small nuclear EBV-encoded RNA was determined by in situ hybridization on paraffin sections using FITC-labeled, EBER1-specific oligonucleotides (Dako) according to the manufacturer's instructions. An anti-FITC MAb, an alkaline phosphate anti-alkaline phosphate (APAAP) mouse system, and the AP substrate BCIP/NBT (Dako) were used in the detection procedure.
P53 Functional Assay in Yeast
RNA was extracted from frozen samples using a Quik-prep micro-mRNA purification kit (Pharmacia; Orsay, France) and was resuspended in 200 μl of elution buffer with 50 U of RNase inhibitor (Sigma; Saint Quentin-Fallavier, France). Random hexamer-primed cDNA was synthesized from 20 μl mRNA in a final volume of 33 μl using a first-strand cDNA synthesis kit (Pharmacia). P53 cDNA was PCR-amplified with primers P3 and P4 as previously described (Flaman et al. 1995). The status of p53 cDNA was then determined using the yeast functional assay (Flaman et al. 1995). Briefly, yeast was co-transformed with PCR-amplified p53 cDNA and a linearized expression vector, and the p53 cDNA was cloned in vivo by homologous recombination. The recipient yeast strain (yIG397) is defective in adenine synthesis because of a mutation in the endogenous ADE2 gene but contains a second copy of the ADE2 open reading frame controlled by a p53-responsive promoter. Therefore, ADE2-mutant strains grown on low adenine plates turn red, yIG397 colonies containing mutant p53 (or no p53) are red, and colonies containing wild-type p53 are white.
Results
All samples obtained after IP injection of PBMCs from EBV-seropositive donors or LCLs into SCID mice were high-grade large B-cell immunoblastic lymphomas with large area of necrosis. The morphology was consistent with the histological description of EBV-associated lymphoproliferative disease or large B-cell lymphoma arising in immunosupressed patients (Na-kamine et al. 1991; Rowe et al. 1991; Veronese et al. 1994). Plasmacyte differentiation was more frequently observed in LCL SCID mice lymphoproliferation than in PBMC tumors (respectively six of 10 cases and one of nine cases (not shown). Immunohistochemical analysis indicated that all tumors were of B-cell origin, as shown by B-cell marker expression (CD20), and were associated with EBV, as confirmed by in situ hybridization with the EBER1 probe (not shown).
The expression of Ki-67 antigen (MIB1), a nuclear protein associated with cell proliferation and found throughout the cell cycle (G1-, S-, G2-, M-phases), was used to verify the proliferative activity of the tumor mass. Immunolabeling for Ki-67 showed strong nuclear reactivity (Figure 1a) in all tumors (Table 1). The percentage of Ki-67-reactive cells ranged from 47 to 97%. No differences were observed between the two groups of tumors (LCL vs PBMC) in terms of proliferative activity.
A p53 functional assay was used to determine p53 gene status in our series of EBV-induced lymphopro liferations. Using this method, previous studies have shown that tissue containing only wild-type p53 give 5-10% red colonies (Flaman et al. 1995), which corresponds to a background resulting mainly from PCR-induced point mutations. The p53 functional assay results from tumors are shown in Table 1. In 14 samples, more than 90% white colonies were found, indicating the presence of wild-type p53 in these samples, whereas one case (M3) exhibited a mutant p53 pattern with 40% red colonies (Figure 2). Because of the lack of enough tumor material, in four samples the p53 gene status could not be determined by the functional assay. We next investigated whether the p53 protein was expressed in the same samples, using IHC analysis (Figure 1d). The results obtained by immunohistochemistry were heterogeneous (Table 1). Positive cases were observed more frequently in the PBMC SCID mice lymphoproliferation group (five of nine cases) compared to the LCL SCID mice lymphoproliferation group (one of 10 cases). Among the positive cases from the first group, one case (M3) showing 20% p53-positive cells by immunohistochemistry was associated with p53 gene mutations, detected by the functional assay. Cases T2 and T3 showed 74 and 11% positive cells by immunohistochemistry, respectively, whereas no p53 mutations were detected by the functional assay. In M1 and M2 cases, 31 and 14% positive cells were detected but we could not evaluate the status of the p53 gene. In the LCL SCID mice lymphoproliferation group, only one case (clo2) showed significant p53 protein expression (16%) by immunohistochemistry, associated with a wild-type p53 gene.
MDM2 protein expression was studied and was compared to p53 expression and gene status in the same samples of tumors. In all the p53(+) and p53(−) samples that contained wild-type p53, a significant nuclear accumulation of MDM2 protein was found compared to control cells (Figure 1b), whereas in the negative control (a case of AIDS-related non-Hodgkin's lymphoma) we did not detect any expression of MDM2 protein (Figure 1c). The rate of MDM2 expression ranged from 40 to 70% of labeled cells (Table 1). Significant levels of MDM2 expression (>50% of labeled cells) were also seen in one unique case (M3) in which mutations of p53 were detected by functional assay.
p21WAFI/CIP1 protein expression (Figure 1e) was compared to p53 gene status and p53/MDM2 expression in the same SCID mice EBV-induced lymphoproliferations. In our series of tumors, the results were heterogeneous (Table 1): Seven of 19 cases showed more than 10% labeled cells (three cases in PBMC SCID mice lymphoproliferation and four cases in LCL SCID mice lymphoproliferation). However, MDM2 protein was also detected in the cases in which p21WAF1/CIP1 protein was expressed.
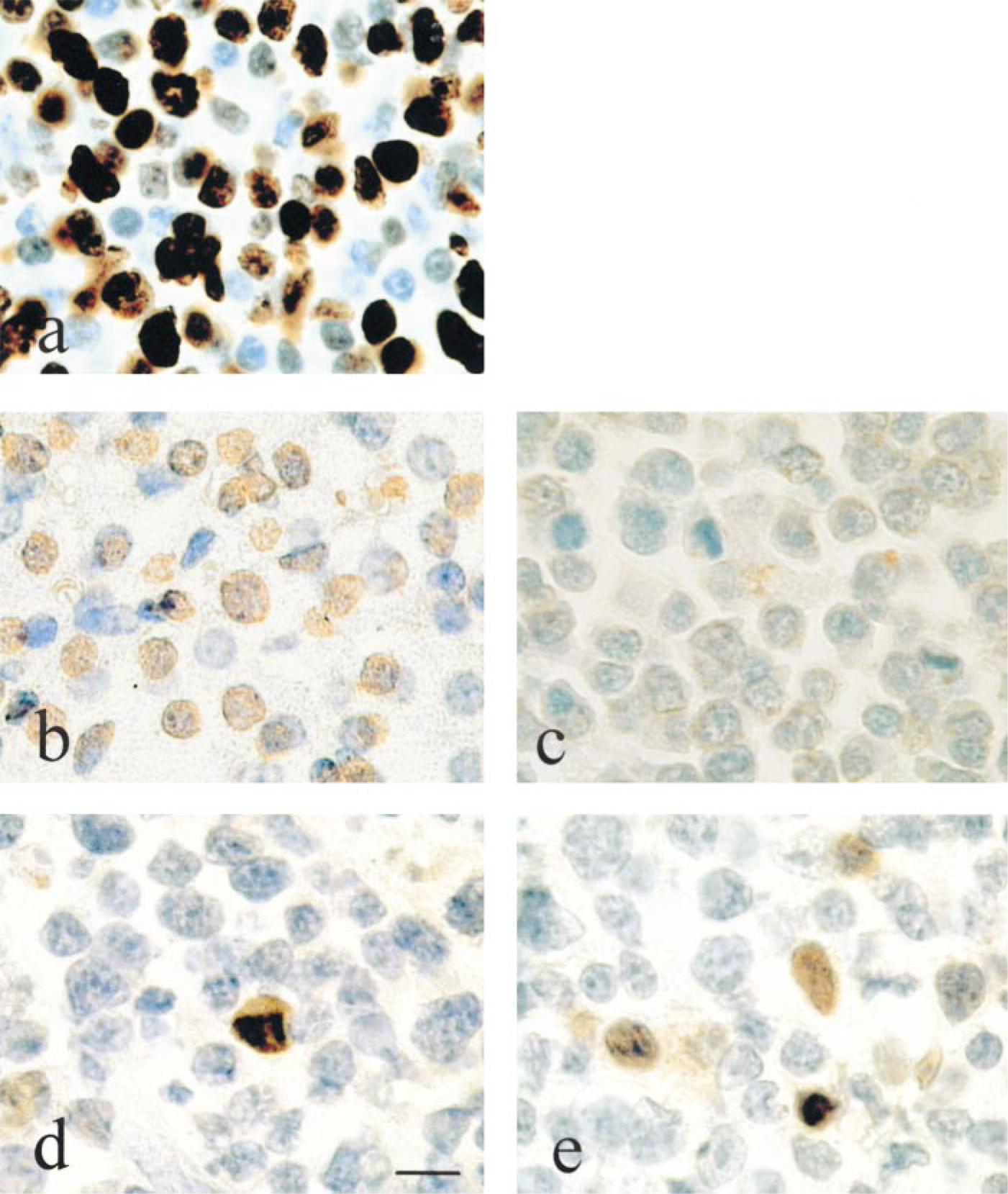
Immunohistochemical patterns for Ki-67, MDM2, p53 and p21WAFI/CIP1 in SCID mouse EBV-induced lymphoproliferation. Magnification is the same in all photographs. Immunolabeling of (a) Ki-67 antigen and (b) MDM2 protein, showing intense positivity in almost all nuclei. (c) An AIDS-related non-Hodgkin's lymphoma (negative control) in which no accumulation of MDM2 protein was detected. (d) p53 and (e) p21WAFI/CIP1 protein immunolabeling in the same samples shows nuclear positivity in some nuclei of tumor cells. Bar = 10 μm.
Discussion
The proliferative activity of our EBV-induced lymphoproliferation was investigated in terms of Ki-67 antigen detection, which has become a standard procedure for assessment of degree of cell proliferation in tumors (Barbareschi et al. 1994; Leers et al. 1997). High levels of this antigen were demonstrated in all samples, indicating that a majority of tumor cells are in the cell cycle. A detailed cell cycle analysis showed that Ki-67 nuclear antigen is expressed in G1, S, G2, and mitosis, but not in G0 (Gerdes et al. 1984). The amount of antigen varies throughout the cell cycle and reaches a maximum during G2- and M-phase (Sasaki et al. 1987; Wersto et al. 1988). Our results are in contradiction to those reported by Rochford et al. (1993), using flow cytometry and acridine orange to analyze the cell cycle ex vivo. Several reasons may explain the discrepancy between Ki-67 score and flow cytometry results. Perhaps most importantly, Ki-67 reactivity includes cells in the G1-phase of the cell cycle, whereas DNA flow cytometry kinetics data exclude this portion of the replicating cells. Furthermore nuclear debris may be variable from case to case, falsely decreasing DNA flow cytometric growth fractions, particularly in high-grade lymphomas with large nuclei, whereas in situ analysis of Ki-67 positive cells allows specific targeting of tumor cells only.
P53 mutations have been shown to occur in various human lymphomas, particularly in Burkitt's lymphoma (Gaidano et al. 1991). We have previously shown (Martin et al. 1998) that the functional assay in yeast allows detection of p53 mutations with a threshold of 10% of mutant cells, whereas the standard SSCP analysis was inefficient under 20% of mutant cells. Therefore, the results obtained in this study with the p53 yeast functional assay enable us to conclude that SCID mice EBV-induced lymphoproliferation is not associated with p53 mutations in all tested samples except one (M3). It is noteworthy that 20% of cells were p53(+) in this sample and that 40% of yeast colonies harbored a mutated p53 cDNA. These results could correspond to an oligoclonal tumor with a particular subset of cells selected for the p53 mutation. Moreover, it should be noted that this tumor was obtained 9 weeks after injection of PBMCs into an SCID mouse, whereas the other PBMC tumors were obtained 11-12 weeks after PBMC injection. The p53 results obtained with the functional assay showed that most SCID mouse EBV-induced lymphoproliferation was not associated with p53 mutations. These results are in agreement with previous results obtained by Murphy and co-workers (Funakoshi et al. 1995), who have shown that tumors arising in human PBL SCID mice did not contain mutations in p53, using immunoprecipitation analysis and wild-type and mutant-specific antibodies to p53. The immunohistochemical results of p53 expression were heterogeneous. We detected p53 accumulation in some cases but no detection of p53 expression in the others. The fact that no p53 mutations were detected by functional assay in the majority of our cases of EBV-induced lymphoproliferation, in spite of its overexpression, can be explained by two mechanisms. The first is an increase in the stability of p53 protein by interaction with another protein, particularly the EBV EBNA5 protein (Szekely et al. 1993). However, if such interactions were obtained in vitro, they remain to be demonstrated in vivo. The second is an overexpression of p53 protein induced by a viral protein such as LMP1 or EBNA2, via activation of
p53 status and expression of MDM2, p21WAF1/CIP1, and Ki-67 antigen in the two groups of SCID mouse EBV-induced lymphoproliferation
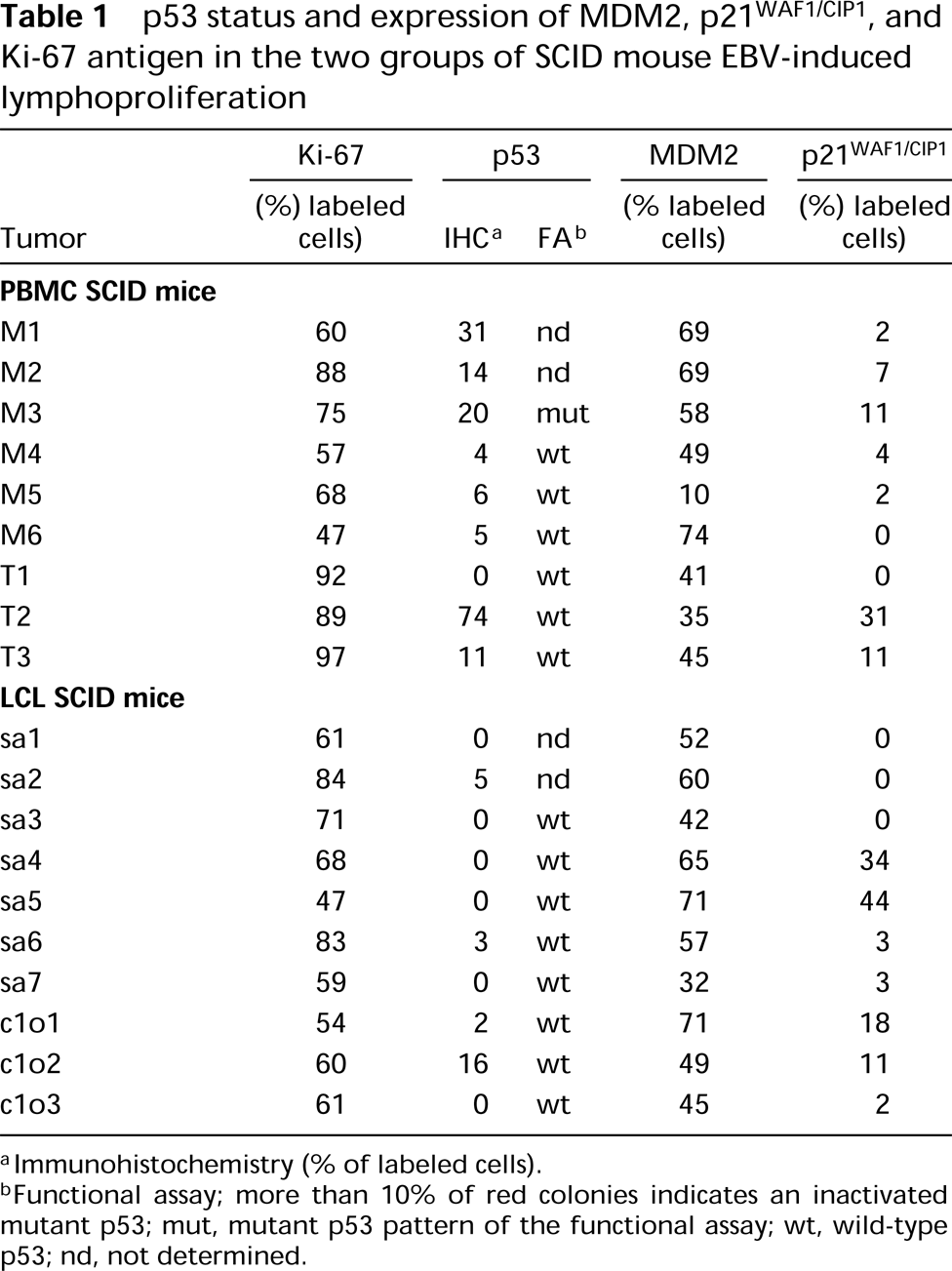
Immunohistochemistry (% of labeled cells).
Functional assay; more than 10% of red colonies indicates an inactivated mutant p53; mut, mutant p53 pattern of the functional assay; wt, wild-type p53; nd, not determined.
Recent studies have shown that the MDM2 gene is a target gene wild-type p53 protein and that an auto-regulatory feedback loop exists between the two molecules, which regulates both the activity of the p53 protein and the expression of MDM2 gene (Barak et al. 1993; Wu et al. 1993). The interactions between p53 and MDM2 in normal and neoplastic cells appear quite complex. Different polypeptides may arise from various spliced mRNAs from the MDM2 gene, from post-transcriptional modifications of MDM2 protein, or from the use of alternative promoters (Piette et al. 1997). The hypothesis that function of the p53 protein can be inactivated by overexpression of MDM2 is not very probable because MDM2 expression is seen independently of the level of p21WAFI/CIP1 expression in all cases of our series of EBV-induced lymphoproliferation. Moreover, previous data showed that MDM2/ p53 complex did not modify p53 protein stability but, conversely, could lead to reduction in p53 protein levels through enhanced proteasome-dependent degradation (Kubbutat et al. 1997). MDM2 could also act independently of p53. It was recently shown that MDM2 can promote tumorigenesis by interacting with Rb (Sun et al. 1998). On the other hand, the accumulation of MDM2 protein in tumor cells might be mainly a consequence of MDM2 gene amplification (Leach et al. 1993; Reifenberger et al. 1993). In our samples, amplification of the MDM2 gene is unlikely to explain the accumulation of MDM2 because previously published data have suggested that MDM2 gene amplification is uncommon in hematological malignancies (Quesnel et al. 1994). Further studies will be conducted to determine the mechanism of accumulation of MDM2. In a series of 17 cases of post-transplant lymphomas, which represent the closest pathological model for SCID mouse EBV-induced lymphoproliferation (Mosier et al. 1992) we found MDM2 expression in 13 of17 cases, analyzed by immunohistochemistry (not shown).
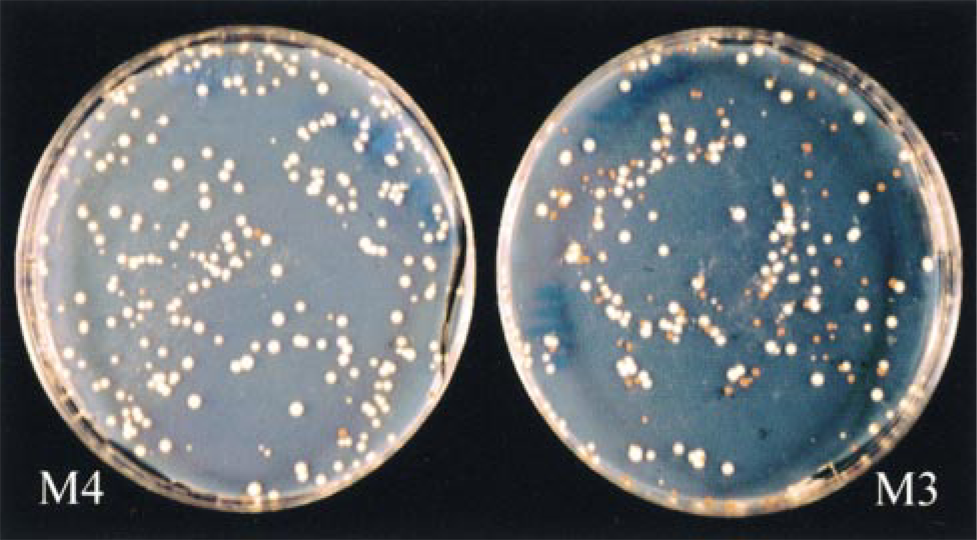
P53 functional assay in yeast in SCID mouse EBV-induced lymphoproliferation. Left, case M4 without inactivating mutation of the p53 gene (5% red colonies); right, case M3 with inactivating mutation of p53 (40% red colonies).
In B-cells infected by EBV, the increased levels of p53 protein imply an induction of p21WAFI/CIP1 expression (Cannell et al. 1998). However, in our series of EBV-induced lymphoproliferation, we were not able to show an obvious link between expression of the two proteins. In addition, the expression of p21WAFI/CIP1 protein in some samples of our series of EBV-induced lymphoproliferation, may be associated with a p53-independent mechanism, as was previously reported by in vivo and in vitro studies (Michieli et al. 1994; Macleod et al. 1995). Moreover, previous data have shown that the levels of p21WAFI/CIP1 expression were not sufficiently high to block the cell cycle in LCL lines (Cannell et al. 1998).
Footnotes
Acknowledgements
Supported by the Agence Nationale de Recherche sur le SIDA (ANRS) 97008 (MR), and the Fondation pour la Recherche Médicale SIDACTION (5ème appel d'offres)(MR).
We thank Dr Alberto Amadoria and Dr Ariana Veronisi (Institute of Oncology; Padova, Italy) for help in reconstitution of SCID mice with human PBMCs. We also thank Jean François Coulon, Martine Moity, and Eric Jeanpierre for excellent technical assistance.