
Abstract
HNF-3/forkhead homologue-4 (HFH-4), a transcription factor of the wingedhelix/forkhead family, was detected by immunohistochemistry in tissue of the developing mouse. HFH-4 protein was present in epithelial cells of the lung, trachea, oviduct, and embryonic esophagus, and in ependymal cells lining the spinal column and ventricles of the brain. In lung, trachea, and nose, HFH-4 was expressed in a distinct subset of epithelial cells that also expressed β-tubulin IV, a ciliated cell marker. Cellular sites of HFH-4 and β-tubulin IV expression were distinct from that of Clara cell secretory protein (CCSP), which was detected in nonciliated epithelial cells in the conducting airway of the lung. HFH-4 and β-tubulin IV, but not CCSP, were detected in the respiratory epithelium of thyroid transcription factor-1 (TTF-1) gene-targeted mice. The presence of HFH-4 and β-tubulin IV in TTF-1 gene-targeted mice demonstrates that differentiation of ciliated epithelium does not require TTF-1. Co-localization of HFH-4 and β-tubulin IV staining in various tissues during mouse development supports a role for HFH-4 in the differentiation of ciliated cell lineages.
Keywords
HNF-3/
In vitro, HFH-4 activated the transcription of CCSP and hepatocyte nuclear factor-3α (HNF-3α) genes (Lim et al. 1997). CCSP, however, is expressed only in non-ciliated respiratory epithelial cells (Singh et al. 1988). Although potential HFH-4 binding sites have been described in several other gene promoters in vitro (Lim et al. 1997), little is known about the target genes regulated by HFH-4 in vivo. To further clarify the temporal and spatial distribution of HFH-4 in vivo, immunohistochemistry of HFH-4 and other epithelial cell markers was determined in the developing mouse.
Materials and Methods
Animals Used
FVB/N and heterozygous TTF-1 (+/-) mice were housed in pathogen-free conditions according to institutional and NIH guidelines. Gestational age of fetal mice was determined by weight and crown-to-rump length. Tissues were fixed overnight at 4C in 4% neutral buffered paraformaldehyde, washed in a graded series of ethanols, and embedded in paraffin. Adult mouse lung was inflation-fixed with 4% neutral buffered paraformaldehyde at 20 cm of water pressure for 1 min, followed by fixation overnight at 4C. Five-μm sections were mounted on poly-lysine-coated slides.
Antibodies and Immunohistochemistry
A rabbit polyclonal antibody against a synthetic peptide spanning amino acid residues 1-101 of mouse HFH-4 fused to GST was generated as described previously (Lim et al. 1997) and either affinity-purified and used at a dilution of 1:1000 or Affi-Gel (BioRad Laboratories; Hercules, CA)-purified and used at a dilution of 1:2000. A rabbit polyclonal antibody generated against a synthetic peptide spanning amino acid residues 110-122 of rat TTF-1 was kindly provided by Dr. Roberto DiLauro (Guazzi et al. 1990) and was used at a dilution of 1:8000 on fetal mouse lung or 1:1000 on adult mouse lung. Rabbit antiserum against rat CCSP was a gift from Dr. Gurmukh Singh (Singh et al. 1988) and was used at a dilution of 1:20,000 on fetal mouse lung or 1:40,000 on adult mouse lung. A mouse monoclonal antibody to β-tubulin IV was obtained from BioGenex (San Ramon, CA) and was used at dilutions of 1:20, 1:40, and 1:80. Specificity of all antibodies has been established previously (Singh et al. 1988; Guazzi et al. 1990; Bannerjee et al. 1992; Lim et al. 1997).
Immunohistochemical staining for HFH-4 was carried out as follows. Five-μm paraffin sections were deparaffinized and slides were heated to 90C in 0.1 M citric acid, 0.1 M sodium citrate (pH 6.0) for 20 min. Sections were then treated with 3% hydrogen peroxide in methanol for 15 min, blocked with 2% normal goat serum for 2 hr, and incubated overnight at 4C with HFH-4 primary antibody, and developed with a biotinylated goat anti-rabbit secondary antibody and a Vector Elite ABC kit (Vector Laboratories; Burlingame, CA). Antigen localization was enhanced with Ni-DAB, followed by incubation with Tris-cobalt and counterstaining with Nuclear Fast Red.
For the co-localization studies, double antibody staining was carried out sequentially. HFH-4, CCSP, or TTF-1 immunohistochemistry was performed first as described above, except that slides were not counterstained with Nuclear Fast Red. After enhancement with Tris-cobalt, slides were washed with water, PBS, then PBS-0.1% Triton X-100 for 5 min each. The tissues were blocked in 5% normal horse serum for 4 hr, followed by incubation overnight at 4C with the β-tubulin IV primary antibody. The reaction was developed using a biotinylated horse anti-mouse secondary antibody and the Vector ABC-alkaline phosphatase kit with Vector-Red as the substrate (Vector Laboratories) and the tissues counterstained with methyl green (Dako; Carpinteria, CA). All immunohistochemical reactions were run with a control reaction lacking the primary antibody to ensure specificity of the staining.
Results
Distribution of HFH-4 Protein in Developing Mouse Tissues
At Day 13.5 p.c., HFH-4 was detected in the nuclei of cells in choroid plexus epithelium (Figure 1A) and ependymal cells lining the lateral ventricle of the brain (Figure 1B). At Day 18.5 p.c., HFH-4 protein was detected in tracheal epithelium (Figure 1C), nasal cavity epithelium (Figure 1E), esophageal epithelium (Figure 1F), and ependymal cells lining the spinal column (Figure 1G). Staining was limited to the nucleus in all tissues. In adult mice, HFH-4 protein was detected in tracheal (Figure 1D) and oviduct epithelium (Figure 1H). HFH-4 was not detected in esophageal epithelium in adult mouse (not shown). HFH-4 was not assessed in brain, nasal epithelium, or testis in adult mice or in developing kidney epithelium.
Developmental Pattern of HFH-4 Expression in Lung
The distribution of HFH-4 protein in lung was examined from Day 13.5 p.c. to adulthood. HFH-4 was not detected in the lung at Day 13.5 p.c. (Figure 2A), corresponding to the early pseudoglandular stage of lung development. By the canalicular stage of lung development at Day 16.5 p.c., HFH-4 was readily detected in nuclei of columnar cells throughout the conducting airway (Figure 2B), including bronchi and bronchioles. This pattern of expression was also observed in the saccular stage of lung development on Day 18.5 p.c. (Figure 2C) and in lungs of neonates (not shown) and adults (Figure 2D). At all time points, HFH-4 was detected in subsets of columnar or basal cells in the conducting airway epithelium and staining was restricted to nuclei. HFH-4 staining was never observed in the alveolar epithelium.
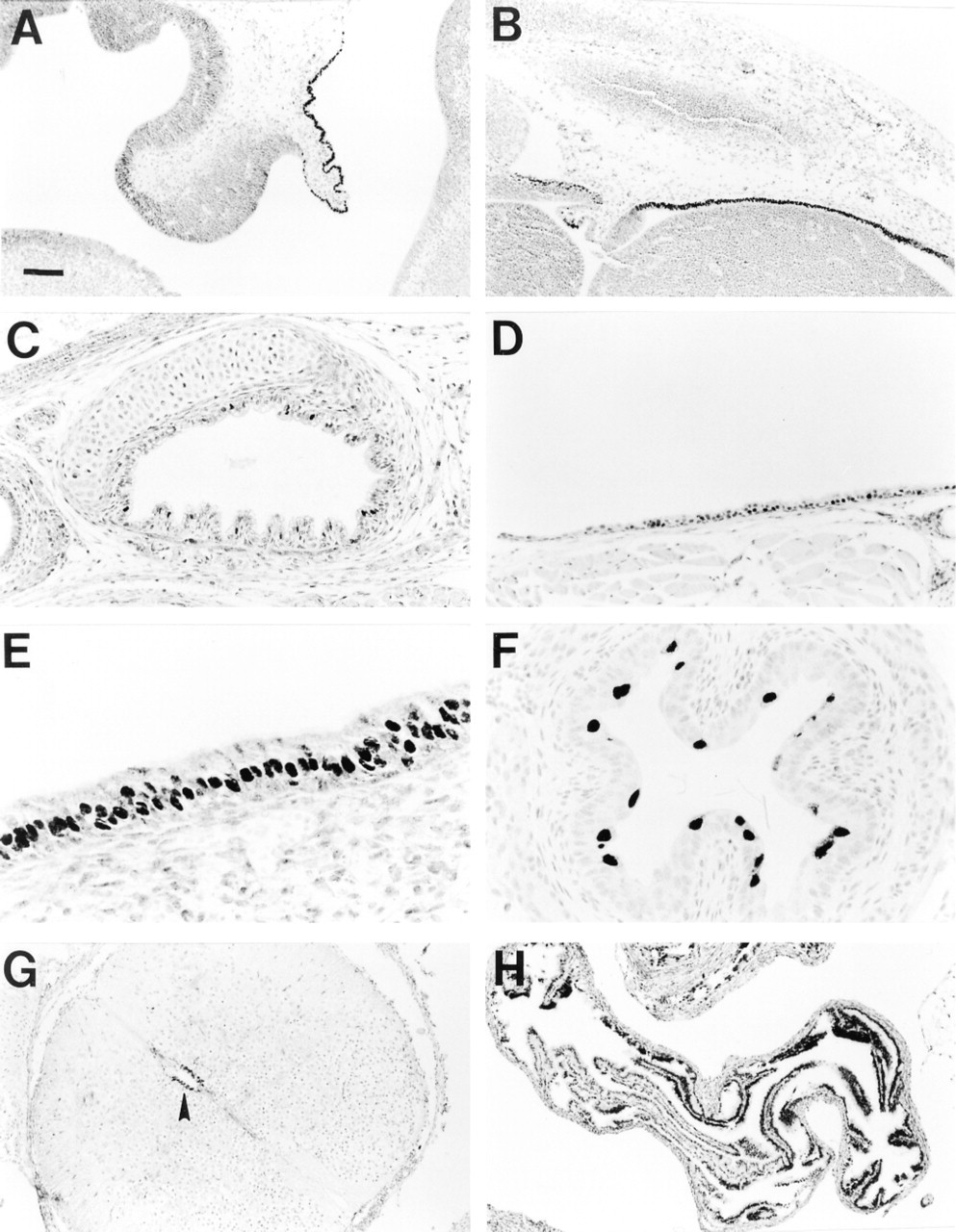
Immunohistochemical localization of HFH-4 in mouse tissues. HFH-4 was detected in nuclei of choroid plexus epithelial cells (A) and ependymal cells lining the lateral ventricle of the brain (B) at Day 13.5 p.c. Staining was present in tracheal epithelium at day 18.5 p.c. (C) and in adult (D). HFH-4 staining in Day 18.5 p.c. mice was also detected in nasal cavity epithelium (E), esophageal epithelium (F), and ependymal cells lining the spinal canal (G, arrowhead) as well as adult oviduct epithelium (H). Bars: A,B,G,H; = 94 μm; C,D = 47 μm; E,F = 24 μm.
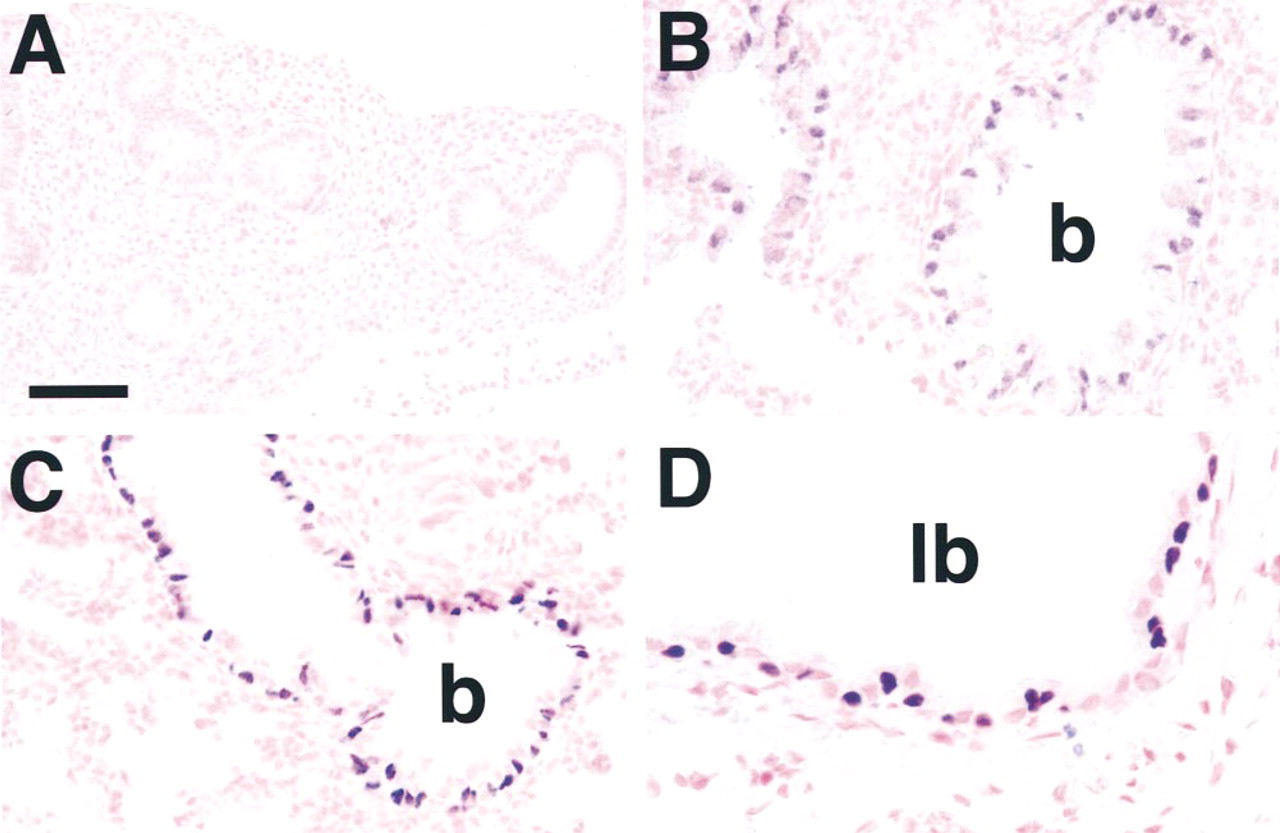
HFH-4 expression during lung development. Staining for HFH-4 was undetectable in lung epithelium at Day 13.5 p.c. (A). HFH-4 was detected in nuclei of a subset of airway epithelial cells at Day 16.5 p.c. (B) and continued at Day 18.5 p.c. (C) and in adult (D). b, bronchiole; lb, lobar bronchus. Bars: A = 40 μm; B,C = 20 μm; D = 10 μm.
Co-localization of HFH-4 and β-tubulin IV
To identify the cell type expressing HFH-4 in the conducting airway epithelium, immunohistochemistry with HFH-4 and β-tubulin IV was performed. Double labeling immunohistochemistry demonstrated that HFH-4 and β-tubulin IV were co-localized in airway epithelial cells. At Day 18.5 p.c., HFH-4 and β-tubulin IV were co-localized in bronchial (Figure 3A), bronchiolar (not shown), and nasal epithelium (Figure 3B). Although HFH-4 was present in the trachea at Day 18.5 p.c., β -tubulin IV was not detected (not shown). In the adult mouse, HFH-4 and β-tubulin IV co-localized in bronchial (Figure 3C), bronchiolar (not shown), and tracheal epithelium (Figure 3D). Staining for HFH-4 and β -tubulin IV was also present in tracheal submucosal glands (Figure 3D, arrow). Although most cells stained for both HFH-4 and β-tubulin IV, cells staining for HFH-4 or β-tubulin IV alone were also detected.
β-Tubulin IV and CCSP Are Expressed in Distinct Subsets of Respiratory Epithelial Cells
CCSP is a secreted protein synthesized by nonciliated respiratory epithelial cells (Singh et al. 1988). Immunohistochemical staining for CCSP and β-tubulin IV was performed on adult and embryonic lung sections. CCSP staining was noted in dome-shaped, nonciliated respiratory epithelial cells (Figures 4A and 4B, brown staining), whereas β-tubulin IV staining was detected in a distinct subset of ciliated cells (Figures 4A and 4B, red staining) at both Day 18.5 p.c. (Figure 4A) and in adult lung (Figure 4B).
Relationships Between TTF-1 and HFH-4
Because TTF-1, a transcription factor of the Nkx family, is required for lung morphogenesis and expression of both CCSP and surfactant proteins (Bohinski et al. 1994; Kelly et al. 1996; Kimura et al. 1996; Zhang et al. 1997), sites of expression of TTF-1 and β-tubulin IV were compared. As previously noted, TTF-1 is found in epithelial cells of the conducting airway, although intensity of TTF-1 staining varies among conducting airway epithelial cells and decreases with advancing age (Zhou et al. 1996). Epithelial cells staining for β-tubulin IV also stained for TTF-1 in lung at Day 18.5 p.c. (Figure 4C) and in adult (Figure 4D).
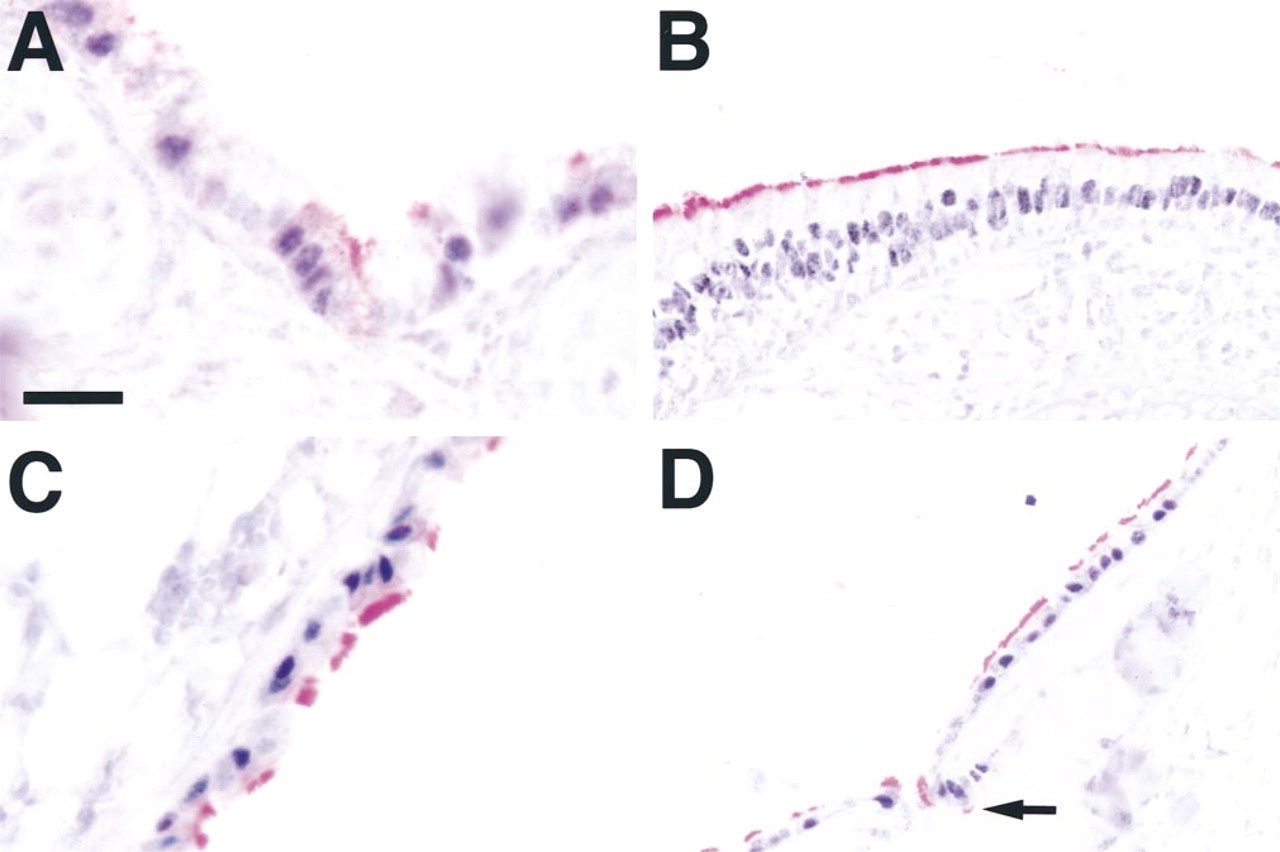
Localization of HFH-4 and β-tubulin IV. Staining for HFH-4 (black) and β-tubulin IV (red) was found to co-localize in the same cells at Day 18.5 p.c. (A,B) in bronchial (A) and nasal (B) epithelium. Co-localization of HFH-4 and β-tubulin IV was also seen in adult mice (C,D) in bronchial (C) and tracheal (D) epithelium. Arrow D indicates staining for HFH-4 and β-tubulin IV in tracheal submucosal gland. Bars: A,C = 10 μm; B,D = 20 μm.
Mice with a targeted ablation of the TTF-1 gene have cystic lung sacs lined with a poorly differentiated epithelium (Kimura et al. 1996) and a tracheo-esophageal fistula consisting of a shared tracheo-esophageal lumen down to the level of the bilateral emergence of two primary bronchi (Minoo and Kimura, unpublished observations). To determine whether TTF-1 is required for HFH-4 or β-tubulin IV expression, lung tissue from TTF-1 null mutant (-/-) mice (Day 16.5 p.c.) was examined for the presence of HFH-4 protein. HFH-4 protein was detected in bronchial and esophageal epithelium of TTF-1 (-/-) mice on Day 16.5 p.c. (Figures 5A–5C) and in the tracheo-esophageal epithelium above the level of the emergence of the bronchi (Figure 5D). HFH-4 protein was not detected in the epithelium lining the cystic lung of TTF-1 (-/-) mice (Figure 5A) on Day 16.5 p.c. but was readily detected in cells of the conducting airways of wild-type mice (see Figure 2B). In addition, β-tubulin IV was detected in bronchial and esophageal epithelium, but not cystic lung epithelium, of TTF-1 (-/-) mice (Figures 5E and 5F). Surfactant protein B, C, and CCSP were not detected in epithelial cells of the lung rudiments of the TTF-1 (- /-) mice (not shown; and Minoo and Kimura, personal communication).
Discussion
This study demonstrates nuclear staining of HFH-4 protein in epithelial cells in various organs of the developing mouse. HFH-4 staining was detected in epithelia at sites consistent with the presence of ciliated cells in various organs, including choroid plexus, nasal cavity, oviduct, tracheal, and lung airway epithelium. This pattern of expression is consistent with a role for HFH-4 in determining ciliated cell differentiation and in the regulation of target genes involved in differentiation of ciliated epithelia. The present findings are consistent with previous studies of HFH-4 mRNA localization (Clevidence et al. 1994; Hackett et al. 1995; Lim et al. 1997; Pelletier et al. 1998). In addition, we report the presence of HFH-4 protein in tracheal and embryonic esophageal epithelium, as well as in ependymal cells lining the spinal canal, sites consistent with expression of HFH-4 in ciliated cells.
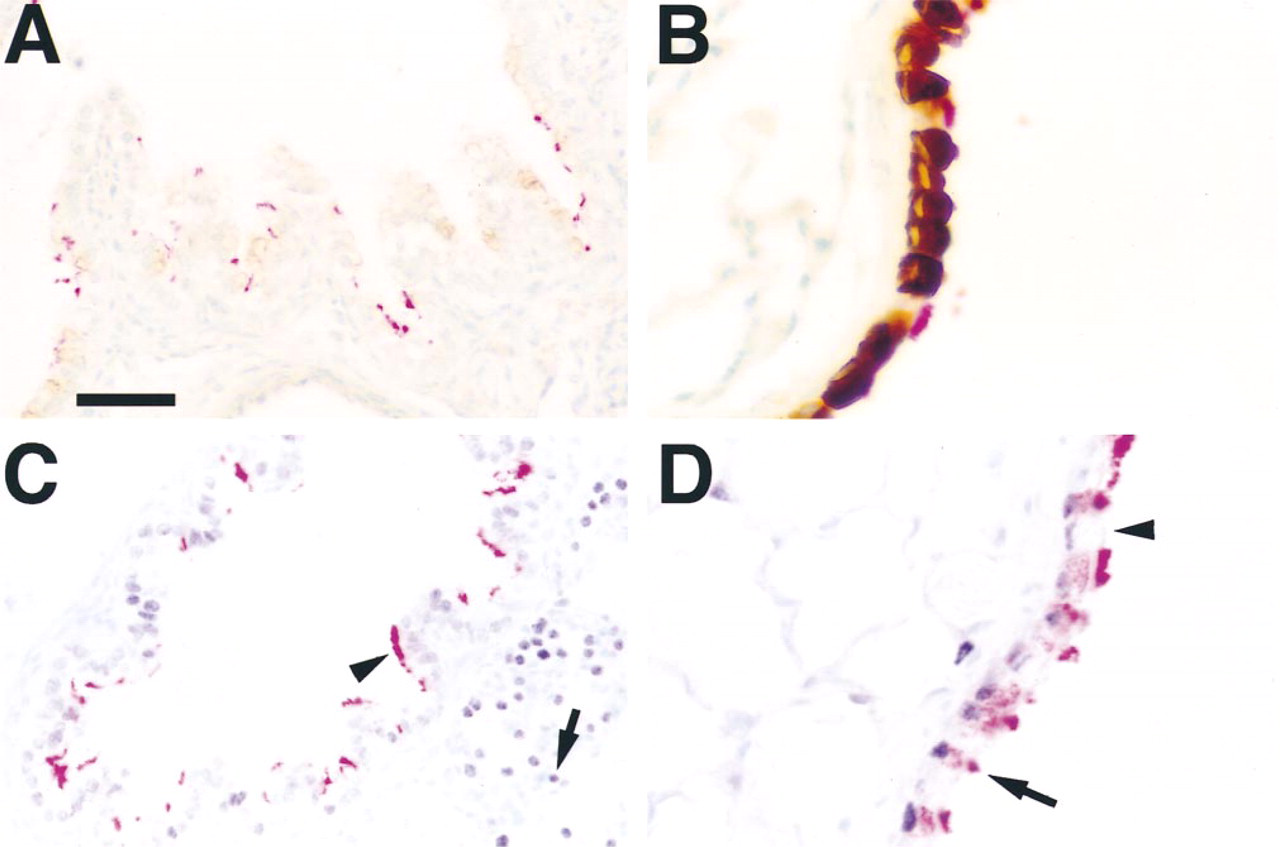
Localization of CCSP, TTF-1, and β-tubulin IV. Staining for CCSP (brown/black) and β-tubulin IV (red) was found in distinct subsets of cells in bronchial epithelium at Day 18.5 p.c. (A) and in adult (B) mouse lung. Staining for TTF-1 (black) and β-tubulin IV (red) was determined in Day 18.5 p.c. (C) and adult (D) lung. TTF-1 staining was most intense in alveolar Type II cells (C, arrow) at Day 18.5 p.c., with less intense staining in airway cells that were positive for β-tubulin IV (arrowhead). TTF-1 staining was found in β-tubulin IV-positive cells in adult mouse airway (D, arrow) and in nonciliated epithelial cells (D, arrowhead). Bars: A,C = 20 μm; B,D = 10 μm.
The developmental expression pattern of HFH-4 was examined more closely in the lung. Nuclear staining of HFH-4 was first detected in airway epithelium of the mouse lung on Day 16.5 p.c. but was not detectable on Day 13.5 p.c., consistent with previous studies demonstrating the presence of HFH-4 mRNA on Day 14.5 p.c. in the mouse (Hackett et al. 1995). HFH-4 staining was present in a subset of airway epithelial cells throughout development and was never detected in the alveolar epithelium. During the pseudoglandular stage of lung development, the respiratory epithelium differentiates from a pseudostratified columnar epithelium to a columnar epithelium in the proximal airways and to a cuboidal epithelium in the distal tubules. The onset of HFH-4 expression in conducting airways correlates with the transition from columnar to cuboidal epithelium, suggesting a role for HFH-4 in regulating this process.
On close inspection, HFH-4 staining appeared to be found in ciliated cells in the conducting airway. Less frequently, staining of what appeared to be basal cells was also observed. Co-localization with antibodies against HFH-4 and β-tubulin IV was performed to determine if staining for these two markers could be detected in the same cell. β-Tubulin IV has been shown to be a marker for ciliated cells in bovine trachea (Renthal et al. 1993) and mouse airway (Fanucchi et al. 1997). β-Tubulin IV and HFH-4 co-localized in ciliated epithelial cells of the mouse respiratory tract, including nasal cavity, tracheal, bronchial, and bronchiolar epithelium. β-Tubulin IV was undetectable, however, in tracheal epithelium at Day 18.5 p.c.; therefore, co-localization with HFH-4 was not observed. Cells staining for HFH-4 or β-tubulin IV, but not for both proteins, were also noted in the conducting airways. In at least some of these cells, this observation may be associated with antigen availability and nuclear localization related to the plane of section. Because HFH-4 staining was detected in some fetal tissues (e.g., trachea) before the appearance of β-tubulin IV, cells staining selectively for HFH-4 may also represent precursor cells capable of differentiation into ciliated cells. Cells staining for HFH-4 alone were also more abundant in fetal than in adult lung tissue (see Figure 3), consistent with the observation that HFH-4 expression preceded that of β-tubulin IV during development. The timing of HFH-4 expression precedes the differentiation of ciliated cells in the lung, which occurs predominantly during the postnatal period in rodents and other species (Kawamata and Fujita 1983; Leigh et al. 1986). The finding that HFH-4 is expressed in tissues with ciliated epithelia is consistent with the hypothesis that HFH-4 plays a role in the differentiation of ciliated or pre-ciliated cells but that its presence is not required for β-tubulin IV transcription per se.
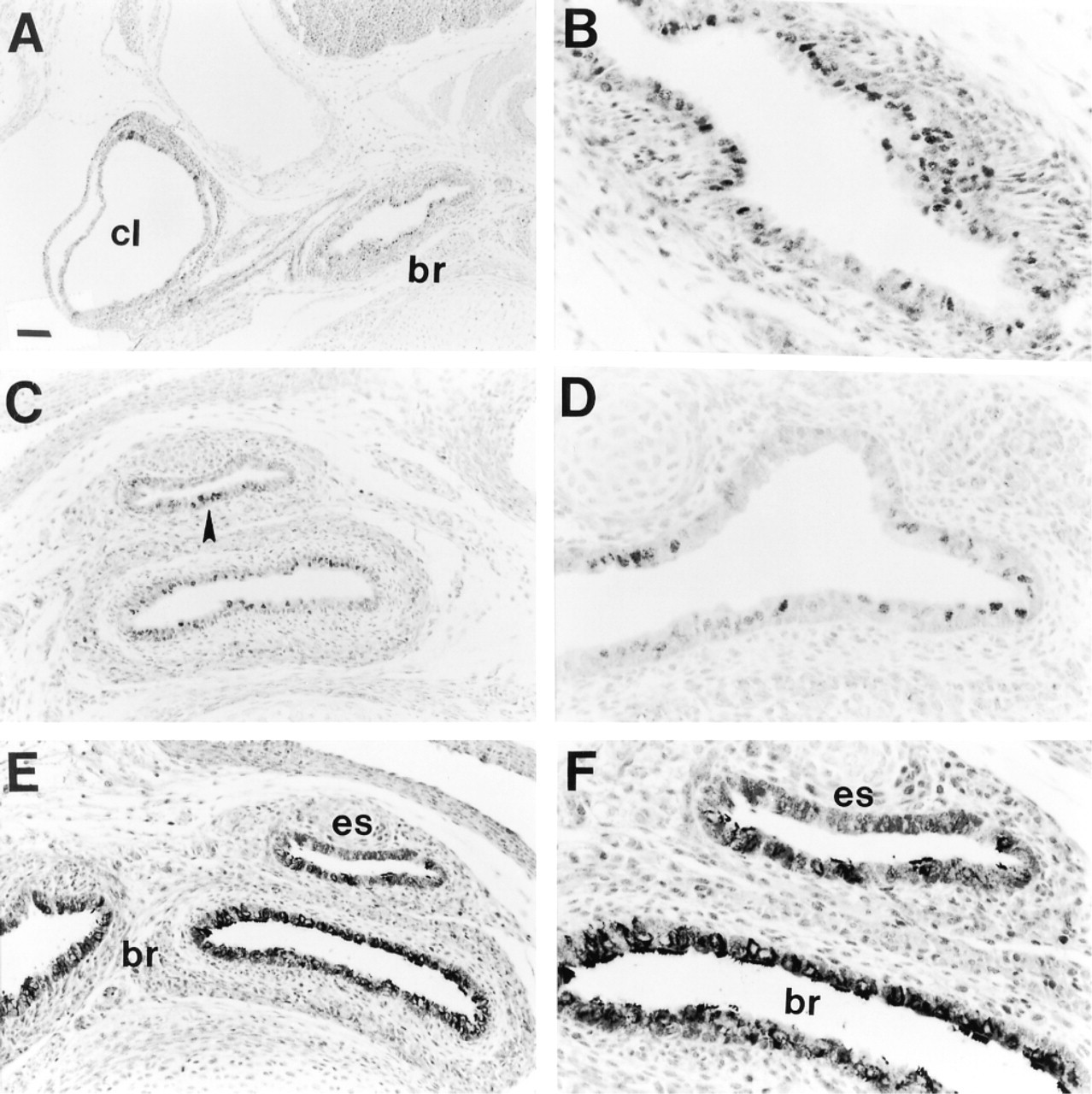
Ciliated cells are present in lungs of TTF-1-deficient mice. Immunohistochemical staining for HFH-4 (A-D) and β-tubulin IV (E,F) was performed on lung tissue from Day 16.5 p.c. TTF-1 (- /-) mice. (A) Low-magnification view of chest cavity showing HFH-4 staining in epithelial cells of bronchus (br) but not in cystic lung (cl). The apparent staining in cystic lung epithelium in A is dark nuclear counterstain due to the arrangement of several layers of cells on top of one another. (B) High-magnification view of the bronchus shown in A, demonstrating nuclear staining for HFH-4 in bronchial epithelium. HFH-4 was also detected in esophageal epithelium below the level of the emergence of the primary bronchi (C, arrowhead) and in tracheo-esophageal epithelium above the level of the emergence of the primary bronchi (D). Staining for β-tubulin IV was also detected in bronchi and esophagus, but not in cystic lung (not shown), of TTF-1 (- /-) mice (E,F). Bars: A = 94 μm; C,E = 47 μm; B,D,F = 24 μm.
The respiratory epithelium is derived from an out-pouching of the foregut endoderm, giving rise to a primitive pulmonary epithelium consisting of columnar cells that are believed to be pluripotent. The primitive pulmonary epithelium differentiates into the cells of the conducting airways and alveolar compartment (Mason et al. 1997). Nonciliated secretory epithelial cells, also termed Clara cells, constitute a large percentage of the surface of conducting airways in the mouse (Plopper et al. 1997). Several studies support the concept that nonciliated secretory cells serve as a stem-cell population during normal differentiation (Plopper et al. 1992) or after injury (Johnson and Hubbs 1990). Basal cells may also serve as a stem-cell population in the airway epithelium (Inayama et al. 1988). It appears that either cell type can give rise to ciliated cells (Nettesheim et al. 1990; Liu et al. 1994), whereas ciliated cells themselves are considered terminally differentiated (Mason et al. 1997). In addition, there is a great deal of plasticity in the respiratory epithelium. For example, fetal tracheal epithelium can adopt an alveolar Type II cell-like phenotype after grafting of distal lung mesenchyme, and fetal distal lung epithelium can adopt a tracheal-like phenotype after grafting of tracheal mesenchyme (Shannon 1994; Shannon et al. 1998). The presence of HFH-4 in ciliated epithelium and basal cells, together with the absence of CCSP, suggests that HFH-4 is involved in ciliated but not Clara cell differentiation.
Mouse models in which HFH-4 protein is ectopically expressed or genetically ablated support a role for HFH-4 in regulating ciliated cell, but not Clara cell, differentiation. Expression of HFH-4 in the distal respiratory epithelial cells of transgenic mice induces ciliated cell characteristics and blocks differentiation of the Clara cell and alveolar Type II cell lineages (unpublished data). In addition, gene targeting of HFH-4 in mice results in a complete lack of ciliated epithelial cells (Chen et al. 1998), supporting the concept that HFH-4 is a critical regulatory transcription factor in the differentiation of ciliated cells.
The poorly differentiated lung structure found in TTF-1 (- /-) mice provides a unique system for studying lung development in the absence of a gene known to play an important role in respiratory epithelial cell differentiation (Kimura et al. 1996). Lack of CCSP, SP-B, and SP-C expression in TTF-1 (- /-) mice demonstrates the critical role of TTF-1 in Clara cell and alveolar Type II cell differentiation and gene expression. The presence of HFH-4 staining in lungs of TTF-1 (- /-) mice indicates that TTF-1 is not required for expression of HFH-4, and the presence of β-tubulin IV in bronchi of TTF-1 (- /-) mice supports the concept that ciliated cell differentiation can occur in the absence of TTF-1. HFH-4 was detected in tracheoesophageal and bronchial epithelium, but not in cystic lung epithelium, of TTF-1 (- /-) mice, suggesting that some level of proximal/distal compartmentalization has occurred in the lungs of TTF-1 (- /-) mice, although distal lung parenchyma has not developed normally. The fact that HFH-4 is expressed in a proximal to distal gradient and that ciliated cells have developed in the absence of TTF-1 suggests that some early components of the developmental program of the lung are determined independently of the onset of TTF-1 expression at Day 9 p.c.
In summary, HFH-4 is expressed primarily in ciliated cells of the respiratory epithelium and in cells that may serve as precursors of ciliated cells in the fetal and postnatal lung. HFH-4 is detected in ciliated epithelia of other tissues, suggesting a role for HFH-4 in differentiation and gene expression in ciliated cells and their precursors. The presence of HFH-4 and β -tubulin IV in the conducting airway epithelium of TTF-1 (- /-) mice indicates that HFH-4 marks subsets of cells distinct from Clara cells or alveolar Type II cells in fetal lung development.
Footnotes
Acknowledgments
Supported by NIH grants HL 56387, by the Program of Excellence in Molecular Biology of the Heart and Lung and HL 41496 (JAW), and by a grant from the Council for Tobacco Research (RHC).
We thank Ms Sherri Proffit for excellent technical help with immunohistochemistry.