
Abstract
Thrombospondin-1 (TSP-1) and an alternatively spliced fibronectin (Fn)-EIIIA isoform are adhesive proteins associated with embryogenesis and tissue remodeling. We compared, by immunohistochemistry and in situ hybridization, the course of TSP-1 and Fn-EIIIA expression in a model of glomerulonephritis induced by Habu snake venom (HV) and characterized by mesangial cell migration, proliferation, and extracellular matrix (ECM) synthesis. At 24 hr after HV, TSP-1 and Fn-EIIIA proteins localized in the central aspects of lesions associated with platelets and macrophages and at the margins of lesions coinciding with mesangial cell migration (determined by Thy-1 staining). Mesangial cells at this time expressed TSP-1 but not Fn-EIIIA mRNA. TSP-1 protein and mRNA peaked in lesions at 48 hr and were associated with cell proliferation (determined by PCNA, α-smooth muscle actin phenotype, and expression of β-PDGF receptor mRNA). TSP-1 expression declined at 72 hr when expression of ECM synthesis peaked, as determined by increased expression of collagen Type IV, laminin, and TGF-β1 protein and mRNA. Mesangial cell expression of Fn-EIIIA was first observed at 48 hr and was most abundant at 72 hr after HV. Therefore, platelet-and macrophage-derived Fn-EIIIA and TSP-1 in early lesions are associated with mesangial cell migration. Mesangial cell upregulation of TSP-1 is associated with migration and proliferation but not maximal ECM accumulation, whereas mesangial cell expression of Fn-EIIIA is associated with proliferation and ECM accumulation. These results suggest distinctive temporal and spatial roles for TSP-1 and Fn-EIIIA in remodeling during glomerular disease.
Keywords
T
Previous studies in our laboratory have shown a progressive course of events involving mesangial cell migration, proliferation, and extracellular matrix (ECM) synthesis during remodeling in a model of proliferative glomerulonephritis induced by Habu snake venom (HV) (Barnes and Abboud 1993; Barnes et al. 1994a,b). We have shown a sequential expression of cellular fibronectin by platelets, macrophages, and mesangial cells during the course of HV-induced glomerular disease, suggesting functional roles for fibronectin in glomerular disease (Barnes et al. 1994a, 1995b). TSP-1 has been linked to PDGF-mediated mesangial cell proliferation in vitro and in vivo (Marinides et al. 1994; Hugo et al. 1995).
TSP-1 and Fn-EIIIA may have important roles in remodeling during the course of glomerular disease that may be related to specific cell behaviors such as migration, proliferation, or ECM synthesis. We were interested in comparing the course of glomerular expression, localization, and cellular sources of these adhesive proteins and their respective mRNAs in the HV model. Our observations suggest distinct temporal and spatial roles for TSP-1 and Fn-EIIIA involving mesangial cell migration, proliferation, and ECM synthesis during remodeling in glomerular disease.
Materials and Methods
Induction of Glomerulonephritis
Glomerular lesions were induced in male Sprague-Dawley rats (Charles River; Raleigh, NC) weighing 200–250 g as previously reported (Barnes 1989; Barnes and Abboud 1993; Barnes et al. 1994a,b). To increase the incidence of glomerular lesions, the rats were unilaterally nephrectomized and 24 hr later were administered HV (Trimeresurus flavoviridis; Sigma Chemical, St Louis, MO) at a dose of 3 mg/kg IV. The rats were sacrificed at 24 (n = 5), 48 (n = 5), or 72 (n = 5) hr after injection of HV, and slices of renal cortex were obtained and immersed in 10% neutral buffered formalin for subsequent processing for light microscopic evaluation. Additional slices of cortex were snap-frozen in liquid nitrogen for subsequent immunofluorescence microscopic identification of cell types and the presence of TSP-1 and Fn-EIIIA protein in glomerular lesions. Separate frozen slices were obtained for in situ hybridization as discussed below. All animal experimentation was conducted in accord with the NIH Guide for the Care and Use of Laboratory Animals.
Characterization of Cell Types in Glomerular Lesions
Previous studies have shown that HV-induced glomerular lesions progress from early mesangial cell migration to development of proliferative micronodules over the course of the disease. Central aspects in early glomerular microaneurysms were deficient of mesangial cells but contained macrophages (Barnes 1989; Barnes and Abboud 1993; Barnes et al. 1994a,b). As the disease progressed mesangial cells filled the central aspects of lesions, forming micronodules consisting almost exclusively of mesangial cells.
Mesangial cells in glomerular lesions were identified as before using mouse monoclonal anti-desmin (Dako; Carpinteria, CA), and mouse monoclonal anti-rat Thy 1.1, clone OX 7 (Accurate Chemical & Scientific; Westbury, NY) as phenotypic markers. Mouse monoclonal anti-α-smooth muscle actin, clone IA4 (Sigma) was also used as a phenotypic marker for activated mesangial cells in late lesions (see below). Platelets were identified using a rabbit antibody to platelet factor 4 (PF4) produced in our laboratory (Barnes 1989). Monocytes and macrophages were identified by mouse monoclonal anti-rat myeloid cell, clone ED-1 (Sero-Tech; Bioproducts for Science, Indianapolis IN).
Fluorescein isothiocyanate (FITC)- or biotin-labeled rat monoclonal anti-mouse IgG, IgM, or goat anti-rabbit IgG were used as second antibodies to identify their respective primary antibodies. Controls consisted of nonimmune IgG or IgM of the appropriate species of primary antibody or diluent without primary antibody. Acetone-fixed frozen sections (6 μm) were treated in a similar fashion as previously described. Sections were incubated with nonimmune IgG of the same species as the second antibody to block nonspecific binding. Methods employing the avidin-biotin complex (ABC) technique for immunoperoxidase localization of diaminobenzidine (DAB) were performed as suggested by the manufacturer (Vector Laboratories; Burlingame, CA). Sections were incubated with 0.6% hydrogen peroxide in methanol to block nonspecific peroxidase activity and with 0.01% avidin, 0.001% biotin to block endogenous biotin activity.
Glomerular Localization of Adhesive Proteins
Glomerular localization of Fn-EIIIA was assessed using a mouse monoclonal antibody (MAb) specific for cellular fibronectin (clone FN-3E2; Sigma). This antibody recognizes the EIIIA domain of cellular fibronectin and does not cross-react with fibronectin derived from plasma (Barnes et al. 1995b). A mouse anti-rat TSP-1 MAb was obtained from Gibco BRL (Gaithersburg, MD) or produced in rabbits immunized against purified human TSP-1 (Calbiochem; La Jolla, CA) in our laboratory. Immunizations and processing of serum were performed as previously described (Barnes 1989).
The presence of adhesive proteins in lesions was assessed by immunofluorescence or immunoperoxidase histochemistry as described above, using appropriate second antibodies. Controls consisted of nonimmune mouse IgM, rabbit IgG, or PBS in place of primary antibody.
Image Analysis
Intensity of peroxidase immunostaining of TSP-1 and Fn-EIIIA in glomerular lesions was evaluated by image analysis measuring the optical density of staining intensity in lesions in captured images. Images of 10 randomly selected glomeruli in each experiment were viewed by a Sony 3CCD color video camera and captured by a Targa+ frame grabber (Truevision; Indianapolis, IN) installed in a personal computer. The data were analyzed using Image-Pro Plus (Media Cybernetics; Silver Spring, MD) image analysis software in which staining intensity was calibrated from 0 to 1 with background (white) level set at 0 and a value of 1 set at black depicting the most intense (dense) level of DAB reaction product. Therefore, each measurement represents an averaged optical density within designated areas of interest (glomerular lesions). Staining intensity was standardized in which no staining above background would have a measurement of 0, whereas a measurement of 1 would be recorded if the entire lesion stained black. Fresh frozen tissue sections were cut at a thickness of 6 μm and slides were stained in batches so that all sections would be treated identically to detect each specific antigen. Data were statistically evaluated by Student's t-test for unpaired data.
In Situ Hybridization
Preparation of riboprobes, tissue preparation, in situ hybridization, and autoradiography were identical to methods used previously (Barnes et al. 1994a,1995a,b). Fragments of cDNA subcloned into plasmids pGEM (Promega; Madison, WI) or Bluescript (Stratagene Cloning Systems; La Jolla, CA) (Table 1) were used to generate 35S-labeled riboprobes to detect TSP-1, Fn-EIIIA, lysozyme, PDGFβ receptor, laminin B1, and Type IV collagen (α1). The plasmids were generously provided by Drs. Hynes, Van De Water, Williams, Yamada, and Vogeli or obtained through the American Type Culture Collection (ATCC; Rockville, MD). The TGF-β1 probe was synthesized in our laboratory (J. Kanalas) by reverse transcription-polymerase chain reaction using primers yielding a 497-BP fragment from base 936–1433 of the reported sequence for rat TGF-β1 (Qian et al. 1990). The fragment was cloned into the pGEM T-vector (Promega) according to the manufacturer's instructions. All experiments were performed simultaneously with sections incubated with sense riboprobes as negative controls.
Analysis of Cell Proliferation
Proliferation of mesangial cells in lesions was assessed in a separate group of 15 rats by identification of cellular incorporation of [3H]-thymidine into nuclear DNA using autoradiographic techniques as reported elsewhere (Barnes et al. 1994b). Rats were unilaterally nephrectomized and given HV as described above. One hour before sacrifice at 24, 48, or 72 hr after HV, the rats were anesthetized and [3H]-thymidine 6.7 Ci/mmol, (Dupont NEN Research Products; Boston, MA) was injected IV into a tail vein at a dose of 1 μCi/g body wt. Kidney cortex was processed for paraffin embedment, sectioned, and autoradiography performed as previously described. Twenty-five randomly selected glomeruli in each kidney section were evaluated for the total number of cells showing incorporation of labeled thymidine by identification of nuclei with at least five grains in the adjacent emulsion.
Cell proliferation was also assessed by immunohistochemical examination of markers of cell proliferation using antibodies to PCNA antigen and α-smooth muscle actin (Johnson et al. 1991). PDGFβ receptor mRNA was assessed by in situ hybridization.
Assessment of Extracellular Matrix
In addition to TSP-1 and Fn EIIIA, glomerular lesions were assessed for the localization of ECM proteins, laminin, and collagen Type IV, by indirect immunofluorescence microscopy utilizing rabbit anti-rat laminin (Chemicon International; Temecula, CA) and anti-mouse collagen IV (Collaborative Biomedical Products; Becton Dickinson Labware, Bedford, MA) as primary antibodies. Localization of TGF-β, which regulates extracellular matrix assembly (Border and Ruoslahti 1990; Yamamoto et al. 1994), was examined by a rabbit pan-specific TGF-β antibody (R&D Systems; Minneapolis, MN). Labeled rat anti-rabbit IgG was used as a second antibody to detect rabbit primary antibodies. mRNAs encoding Type IV collagen (α1), laminin B1, and TGF-β1 were detected by in situ hybridization.
cDNA for in vitro transcription of riboprobes
Results
Morphology
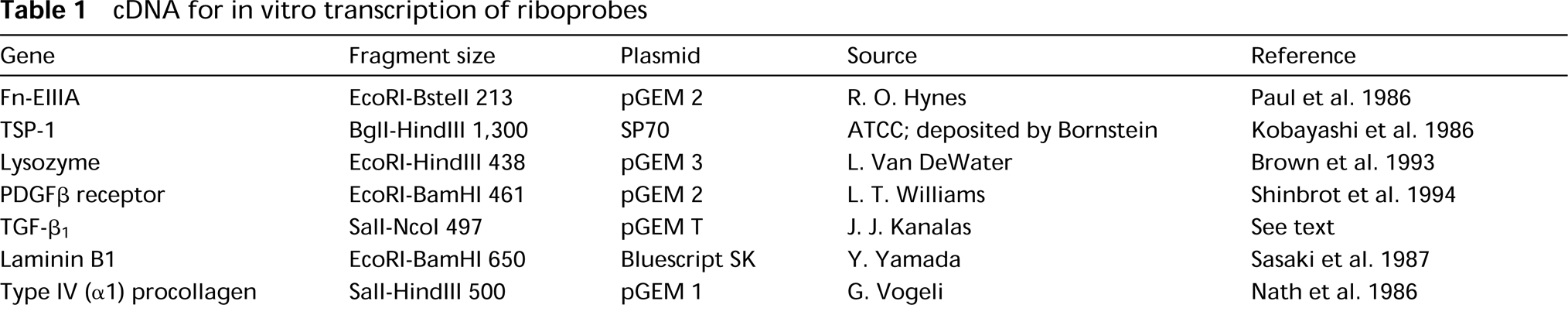
Glomerular lesions appeared as previously described and involved the development of microaneurysms in early lesions containing platelet aggregates, leukocytes, erythrocytes, and plasma proteins, progressing to micronodules composed of confluent masses of cells. Cell types were identified in discrete locations in glomerular lesions throughout the course of the disease. In early lesions (24 hr), mesangial cells were identified by the phenotypic markers Thy-1 and desmin localized at the margins of glomerular lesions adjacent to the glomerulus-tuft interface and the periphery of microaneurysms (Figure 1a), but not in the central aspects of glomerular lesions. Conversely, platelets identified by PF4 staining and macrophages identified by ED-1 surface antigen and lysozyme mRNA were observed primarily in the central aspects of microaneurysms and only occasionally at the margins of lesions. By 48 hr after HV, microaneurysms were filled with mesangial cells (Figure 1b) and macrophages, and by 72 hr lesions were characterized by confluent masses of mesangial cells that formed micronodules (Figure 1c) containing few macrophages. Platelets diminished over time as lesions filled with mesangial cells, as previously reported (Barnes 1989).
Expression of TSP-1
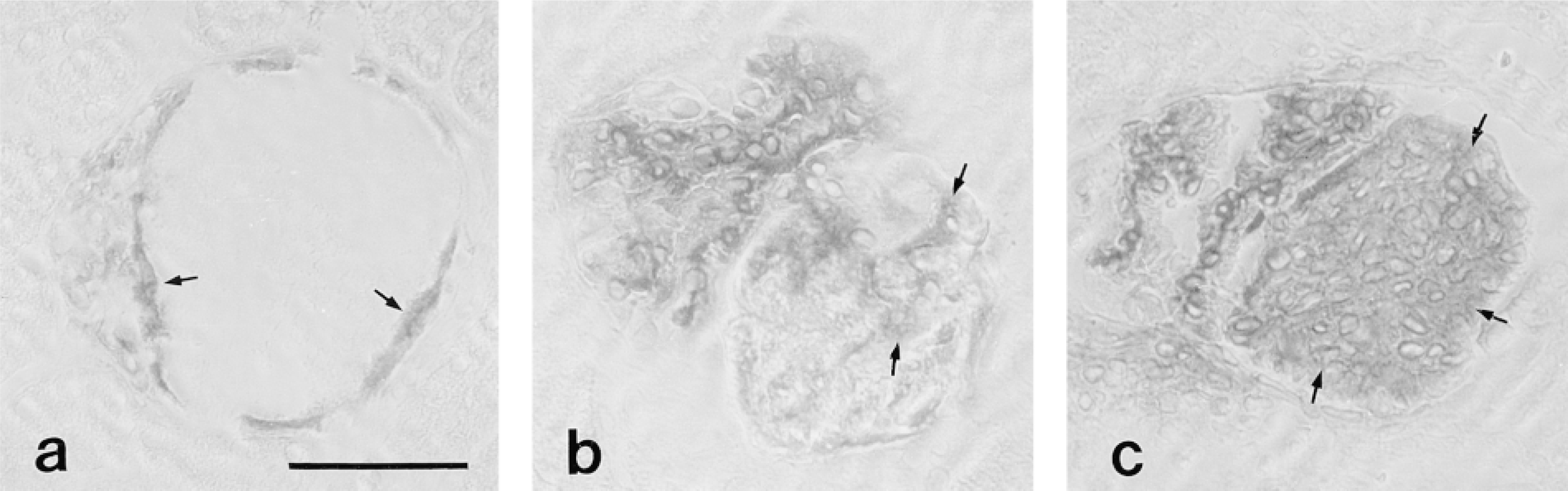
Normal glomeruli did not express TSP-1 protein or mRNA. Immunofluorescence staining of TSP-1 protein in glomerular lesions peaked at 48 hr and diminished by 72 hr after HV-induced glomerulonephritis (Figures 2 and 4). Localization of TSP-1 protein in glomerular lesions at 24 hr after HV showed staining of platelet aggregates in the central aspects and margins of glomerular lesions (Figure 2a). Staining for TSP-1 at the margins of lesions was more intense than that observed within the central aspects of glomerular lesions at this time. Intensity of staining for TSP-1 peaked in glomerular lesions at 48 hr after HV (Figure 2b), and was associated with an influx of mesangial cells and macrophages within lesions at this time. At 72 hr after HV, staining for TSP-1 protein was associated with the mesangial cell population within micronodules but was less intense than at 48 hr (Figures 2c and 4). TSP-1 was also associated with parietal epithelial cells in Bowman's capsule adjacent to glomerular lesions in a similar distribution and time course as described above for Fn-EIIIA.
In situ hybridization frequently revealed glomeruli with a highly localized expression of TSP-1 mRNA in cells at the glomerular tuft-lesion interface (Figure 2d), corresponding to the localization of intense protein staining at these locations. Cells identified as macrophages within the central aspects of microaneurysms at 24 hr after HV did not express TSP-1 mRNA, indicating that these cells do not synthesize detectable levels of TSP-1 mRNA in this setting. However, at 48 hr after HV, cells in the central aspects and margins of lesions expressed abundant TSP-1 message (Figure 2e), corresponding to peak mesangial cell proliferation (see below). Message for TSP-1 diminished 72 hr after HV (Figure 2f), but low levels could be detected in most cells within micronodules. Parietal epithelial cells lining Bowman's capsule expressed TSP-1 mRNA at 24, 48, and 72 hr after HV, corresponding to the detection of protein by immunohistochemistry.
Immunoperoxidase localization of Thy-1 antigen marking the position of mesangial cells in glomerular lesions at 24 (a), 48 (b), and 72 (c) hr after injection of HV. Mesangial cells (arrows) are present at the interface between an intact glomerular capillary tuft and the lesion and the marginal aspects of glomerular microaneurysms at 24 hr after HV. Mesangial cells are present within the microaneurysm at 48 hr and form a confluent mass of cells (micronodule) by 72 hr after HV. Bar = 50 μm.
Expression of Fn-EIIIA
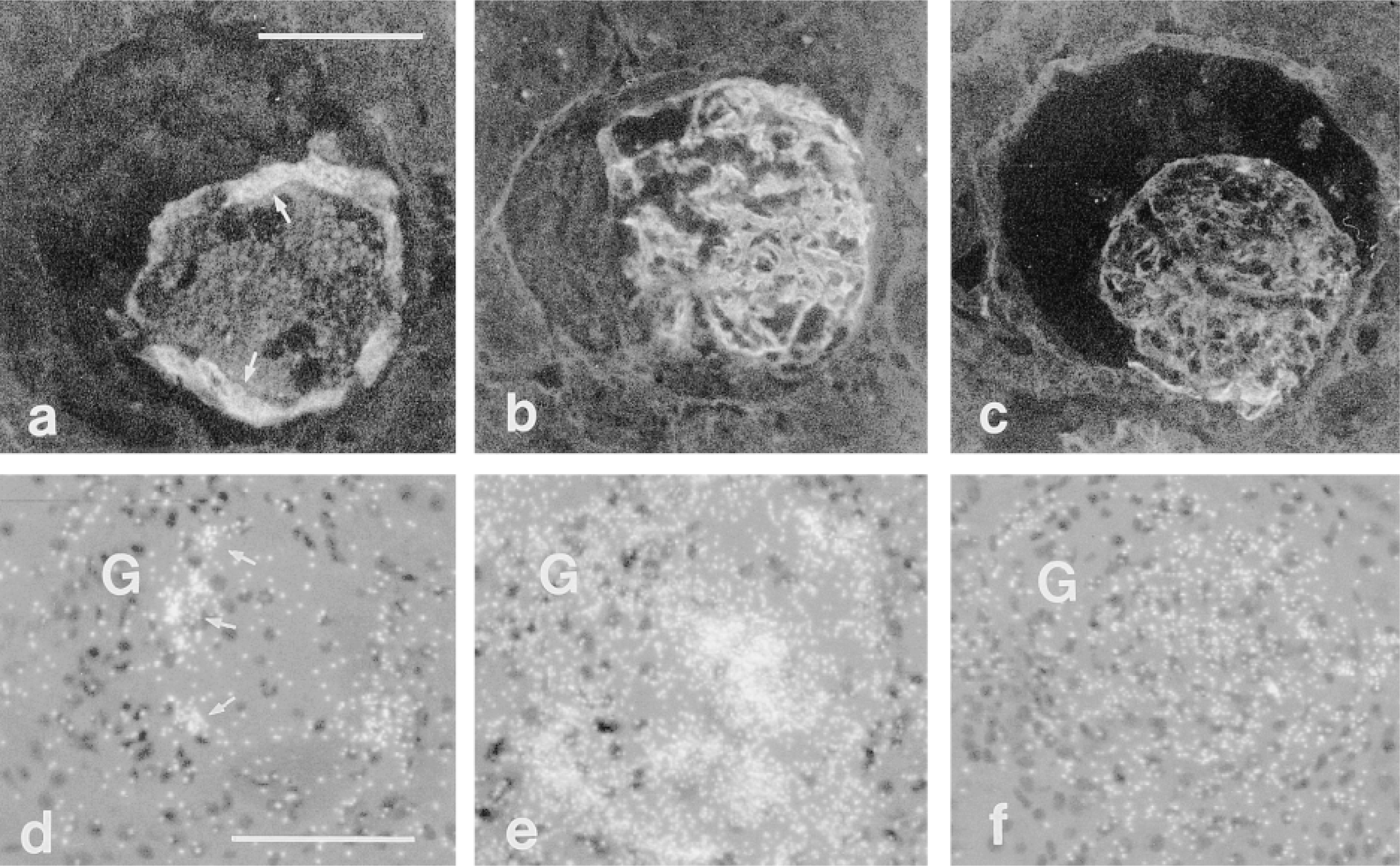
Expression of EIIIA-Fn was identical to that previously described (Barnes et al. 1995b). Little Fn-EIIIA was detectable in the mesangium of normal glomeruli. Staining of Fn-EIIIA in glomerular lesions followed a progressive increase in staining intensity over the course of HV-induced glomerulonephritis (Figures 3 and 4). At 24 hr after HV, faint staining of Fn-EIIIA was observed in platelet aggregates within central aspects and was often partitioned to the margins of lesions (Figure 3a). Expression of Fn-EIIIA protein progressively increased in intensity over time and was most abundant at 72 hr after HV (Figures 3b and 3c), corresponding to mesangial cell occupation of lesions. In situ hybridization revealed expression of Fn-EIIIA mRNA first in cells identified to be macrophages within the central aspects of lesions (Figure 3d), and only occasional expression at the margins of lesions at 24 hr after HV. Message for Fn-EIIIA mRNA increased at 48 hr (Figure 3e) and peaked at 72 hr after HV (Figure 3f), coinciding with Fn-EIIIA protein expression and mesangial cell occupation of micronodules at these times.
Cell Proliferation
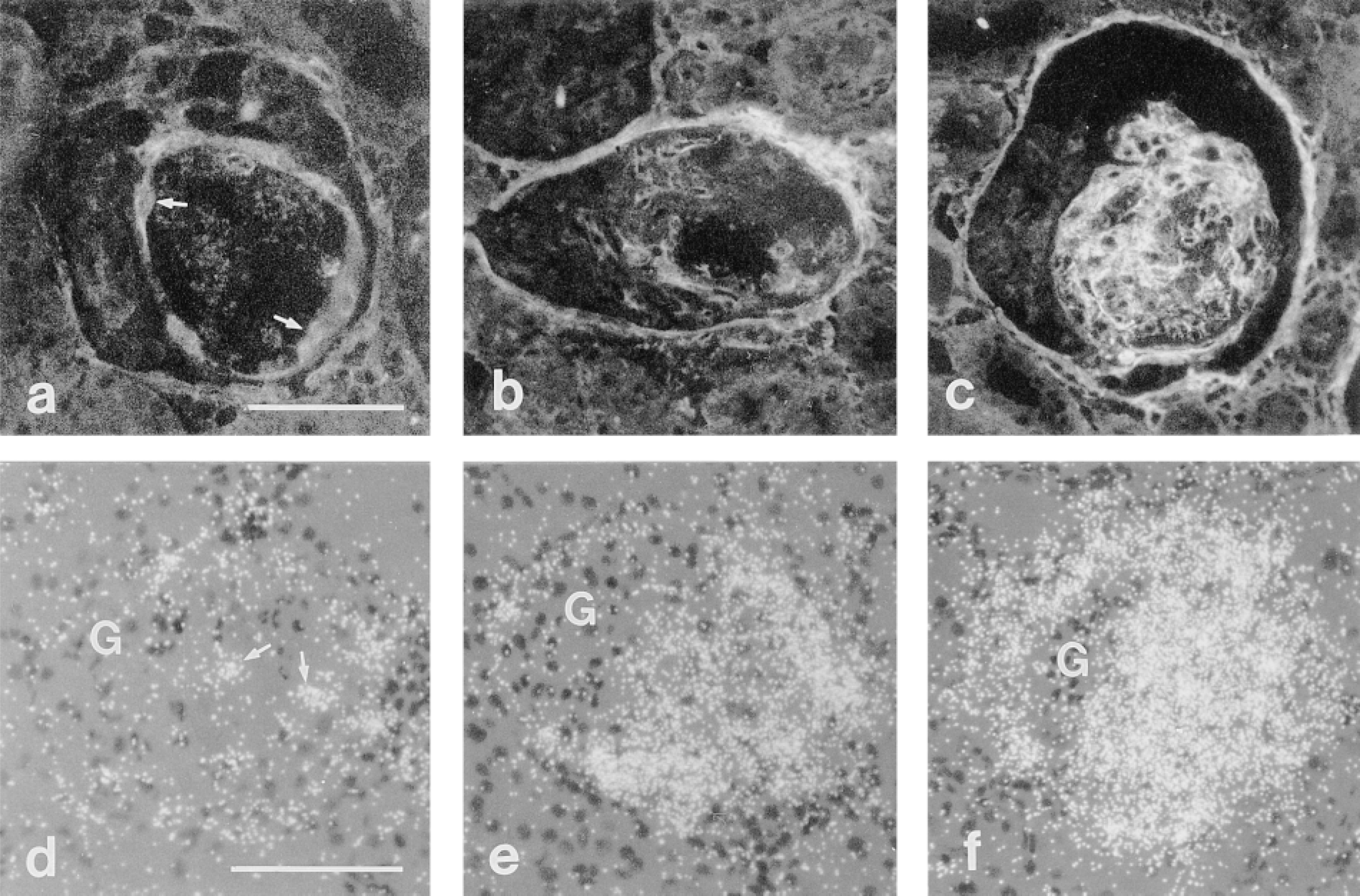
Glomerular lesions at 24 hr after HV showed negligible staining for markers of cell proliferation (α-smooth muscle actin protein or PDGFβ receptor) and little uptake of [3H]-thymidine by autoradiography. Cell proliferation in glomerular lesions peaked at 48 hr as assessed by PCNA (Figure 5a) and cell incorporation of [3H]-thymidine (Figure 5d), as described previously (Barnes and Abboud 1993; Barnes et al. 1994b). Beginning at 48 hr and continuing through 72 hr after HV injection, expression of α-smooth muscle actin (Figure 5b), and PDGFβ receptor mRNA increased (Figure 5c).
Extracellular Matrix Synthesis
Expression of mRNA and protein for additional ECM components, Type IV collagen and laminin, were most abundant and predominately associated with mesangial cells in micronodules at 72 hr after HV (Figures 6a and 6b). Expression of ECM was associated with the maximal expression of TGF-β1 mRNA and protein at 72 hr after HV by in situ hybridization and immunofluorescence microscopy, respectively (Figures 6c and 6d). Expression of TGF-β was negligible at earlier times.
Glomerular localization of TSP-1 protein (a-c) and mRNA (d-f) by immunofluorescence and in situ hybridization, respectively. TSP-1 protein localizes to platelet aggregates in the central aspects and at the margins (arrows) of lesions at 24 hr (a), peaks in intensity at 48 hr (b), and diminishes at 72 hr (c) after HV. TSP-1 mRNA is expressed in marginating mesangial cells (arrows) but not in macrophages located in microaneurysms 24 hr after HV. Intensity of mRNA message corresponds to TSP-1 protein, peaking at 48 hr and waning at 72 hr after HV (d-f). G, intact glomerular capillary tuft. Bars = 50 μm.
Discussion
Previous studies in our laboratory have characterized a sequence of events involving mesangial cell migration, proliferation, and ECM synthesis during remodeling in HV-induced glomerular injury (Barnes and Abboud 1993; Barnes et al. 1994b,1995a). We have also shown that an alternatively spliced fibronectin isoform, Fn-EIIIA, is differentially expressed by platelets, macrophages, and mesangial cells throughout the course of this disease (Barnes et al. 1994a,1995b). In this study, the temporal and spatial expression of TSP-1 was compared to that of Fn-EIIIA during the course of HV-induced glomerular disease and also compared to cellular events involved in remodeling using markers for proliferation and ECM synthesis. This study shows that despite the fact that TSP-1 and Fn-EIIIA are synthesized and/or secreted by similar cell types, the cellular sources and patterns of peak expression of each protein follow a different course, corresponding to particular cellular events associated with remodeling, and indicate that TSP-1 and Fn-EIIIA are differentially regulated during the progression of glomerular disease.
The roles of TSP-1 and Fn-EIIIA during remodeling in vivo are not clear. However, the onset, peak expression, cellular sources, and known effects of these proteins in vitro suggest functional relevance for both adhesive proteins in glomerular disease. For example, platelet-derived TSP-1 and Fn-EIIIA in early lesions appear to be associated with mesangial cell migration. Mesangial cell expression of TSP-1 appears to be associated with migration and proliferation but not ECM accumulation, whereas mesangial cell expression of Fn-EIIIA is associated with proliferation and ECM accumulation.
Glomerular localization of Fn-EIIIA protein (a-c) and mRNA (d-f) by immunofluorescence and in situ hybridization, respectively. Fn-EIIIA protein localizes to platelet aggregates and at the margins of lesions (arrows) at 24 hr (a) and progressively increases in intensity at 48 hr (b) and 72 hr (c) after HV. Fn-EIIIA is most intense in micronodules at 72 hr after HV. By in situ hybridization, Fn-EIIIA mRNA is expressed in macrophages located in the central aspects of microaneurysms (arrows), but not in marginating mesangial cells 24 hr after HV (d). Intensity of message increases over time and corresponds to Fn-EIIIA protein at 48 and 72 hr after HV (e,f). G, intact glomerular capillary tuft. Bars = 50 μm.
This and previous studies have characterized discrete cellular events involving cell migration, proliferation, and ECM synthesis over the course of HV-induced glomerular disease. Glomerular lesions in this model are characterized by early microaneurysms, constituting a highly localized milieu replete with platelets and macrophages and their biologically active secretory products (Barnes 1989; Barnes and Abboud 1993; Barnes et al. 1994a,b). The early lesions are devoid of resident glomerular cells and are separated from intact glomerular structures. Over the course of 3 days after injection of HV, mesangial cells migrate to the margins of lesions and proliferate, ultimately forming micronodules composed of a confluent mass of mesangial cells. Mesangial cell migration occurred before proliferation (Barnes et al. 1994b) and peak cell proliferation occurred before maximal ECM synthesis (Barnes and Abboud 1993; Barnes et al. 1994a,1995b).
The exact roles of TSP-1 and Fn-EIIIA in glomerular injury and repair have not been defined. However, the above data are consistent with known functions of these adhesive proteins in vitro and in vivo. For example, TSP-1 and fibronectin are chemotactic for a variety of cell types, including leukocytes, fibroblasts, smooth muscle cells, and epithelial and endothelial cells (Hynes 1990; Sage and Bornstein 1991). Similarly, we have shown that glomerular mesangial cells migrate in response to platelet fibronectin (Barnes and Hevey 1991), which has a high content of cellular (Fn-EIIIA) isoform (Paul et al. 1986; Peters et al. 1995). TSP-1 is also chemotactic to mesangial cells in vitro (Taraboletti et al. 1992), and both TSP-1 and Fn-EIIIA are enhanced around migratory tracts and areas of cell proliferation during embryogenesis (ffrench-Constant and Hynes 1988,1989; Hynes 1990; O'Shea et al. 1990) and wound repair (Raugi et al. 1987; ffrench-Constant et al. 1989; Brown et al. 1993; Reed et al. 1993; DiPietro et al. 1996). We identified a partitioning of TSP-1 and Fn-EIIIA protein at the margins of glomerular lesions at 24 hr after HV, specifically where mesangial cells migrate in early lesions, suggesting that TSP-1 and Fn-EIIIA may provide a provisional substratum and modulate cell adhesion and migration during the early stages of glomerular injury. Our results also indicate that mesangial cells at the margins of lesions synthesize TSP-1 but do not express Fn-EIIIA mRNA, suggesting that migrating mesangial cells rely on their own synthesis of TSP-1 but not Fn-EIIIA. In addition, platelets appear to be important sources of both adhesive proteins in early lesions and may have a role in cellular repopulation of glomerular lesions before mesangial cell expression of these proteins. Macrophages appear to provide an exogenous source of Fn-EIIIA but not TSP-1 in glomerular lesions, unlike that described for wound healing, in which both adhesive proteins are synthesized by macrophages (Brown et al. 1993; DiPietro et al. 1996).
TSP-1 may also play an indirect role in remodeling by modulating extracellular proteolysis. Plasminogen and plasminogen activators (PAs) and their inhibitor, plasminogen activator inhibitor-1 (PAI-1), are associated with cells actively involved in degradation of ECM required for motility and proliferation (Vassalli et al. 1991), and components of the plasmin/plasminogen system have been used as histochemical markers for cell migration or active remodeling (Feinberg et al. 1989; Sappino et al. 1989; Pepper et al. 1992; Barnes et al. 1995a). Because TSP-1 binds PAs (Harpel et al. 1990), the effect of TSP-1 on cell migration may be related to its ability to act as a focus for protease generation (Dixit 1992) or inhibition (Hogg et al. 1992) during remodeling. In support of this hypothesis is the observation that TSP-1 localizes at the margins of early glomerular lesions in the same areas in which we previously reported mesangial expression and localization of PAI-1 (Barnes et al. 1995a).
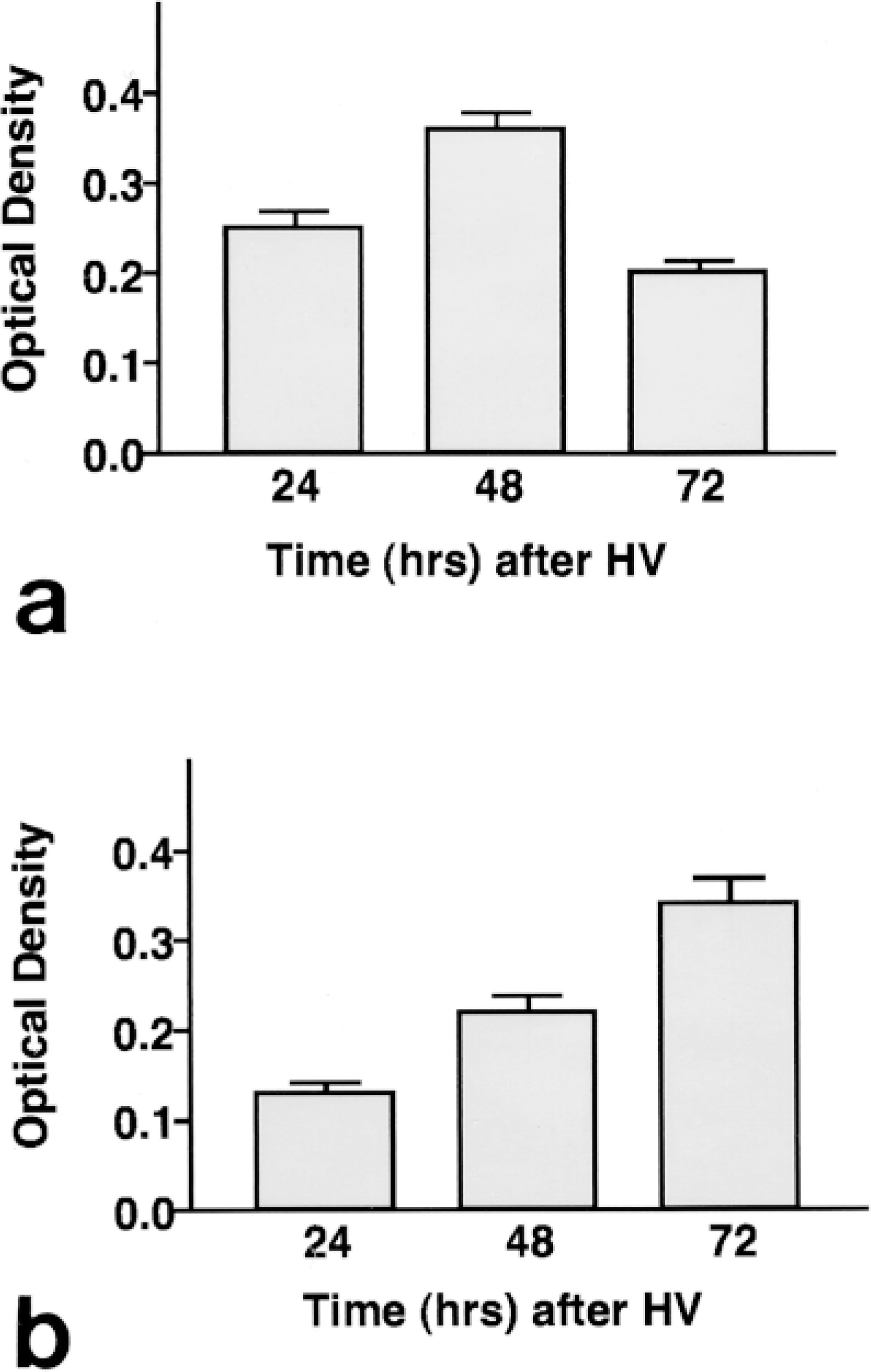
Intensity of immunoperoxidase staining for TSP-1 and Fn-EIIIA protein measured by image analysis. Staining (optical density) of reaction product (diaminobenzidine) for TSP-1 (a) peaks at 48 hr after HV, whereas staining for Fn-EIIIA (b) progressively increases, reaching its highest intensity at 72 hr after HV.
TSP-1 and Fn-EIIIA have also been implicated in cell proliferation during cell remodeling after injury (Hynes 1990; Barnes et al. 1995b; Hugo et al. 1995). TSP-1 and Fn are known mitogens for mesangial cells in culture (Simonson et al. 1989; Kobayashi and Yamamoto 1991; Marinides et al. 1994; Hugo et al. 1995), and a recent report correlates TSP-1 with proliferation and PDGF expression in vitro and in vivo during the proliferative phase of the anti-Thy-1 model of proliferative glomerulonephritis (Hugo et al. 1995). Our results also show that TSP-1 expression is associated with cell proliferation. Localization of TSP-1 protein and expression of mRNA by in situ hybridization were maximal at 48 hr after HV, at a time when cell proliferation, as measured by [3H]-thymidine incorporation, was determined to peak. Mesangial cells acquired expression of α-smooth muscle actin and PDGFβ receptor at 48 hr after HV, both of which have also been linked to cell proliferation in several models and clinical settings of glomerular remodeling (Johnson et al. 1991; Alpers et al. 1992).
Fn-EIIIA expression by mesangial cells was identified at 48 hr after HV, but maximal expression was not observed until 72 hr after HV, when expression of collagen, laminin, and TGF-β were maximal, suggesting that expression of this adhesive protein by mesangial cells may function in cell proliferation as discussed above. However, peak abundance is more closely associated with ECM accumulation (McDonald 1988; Border and Ruoslahti 1990; Morla et al. 1994). Fn may provide a foundation for subsequent matrix assembly because of its different binding domains for collagen, laminin, heparan sulfate, and other Fn molecules (McDonald 1988; Hynes 1990; Morla et al. 1994). TGF-β regulates extracellular matrix accumulation (Ignotz and Massague 1986; Roberts et al. 1990) and preferentially stimulates fibroblast synthesis of Fn-EIIIA (Balza et al. 1988). Moreover, expression of Fn-EIIIA protein has been associated with TGF-β1 mRNA in several glomerular diseases involving matrix expansion (Border and Ruoslahti 1990; Kagami et al. 1993; Yamamoto et al. 1993,1994).
Conversely, mesangial cell expression of TSP-1 protein and mRNA peaked at 48 hr but waned by 72 hr after HV. Our observations of a decrease in TSP-1 expression at the time of maximal mesangial expression of TGF-β1 and extracellular matrix proteins suggest that TSP-1 is dissociated from matrix accumulation in this model.
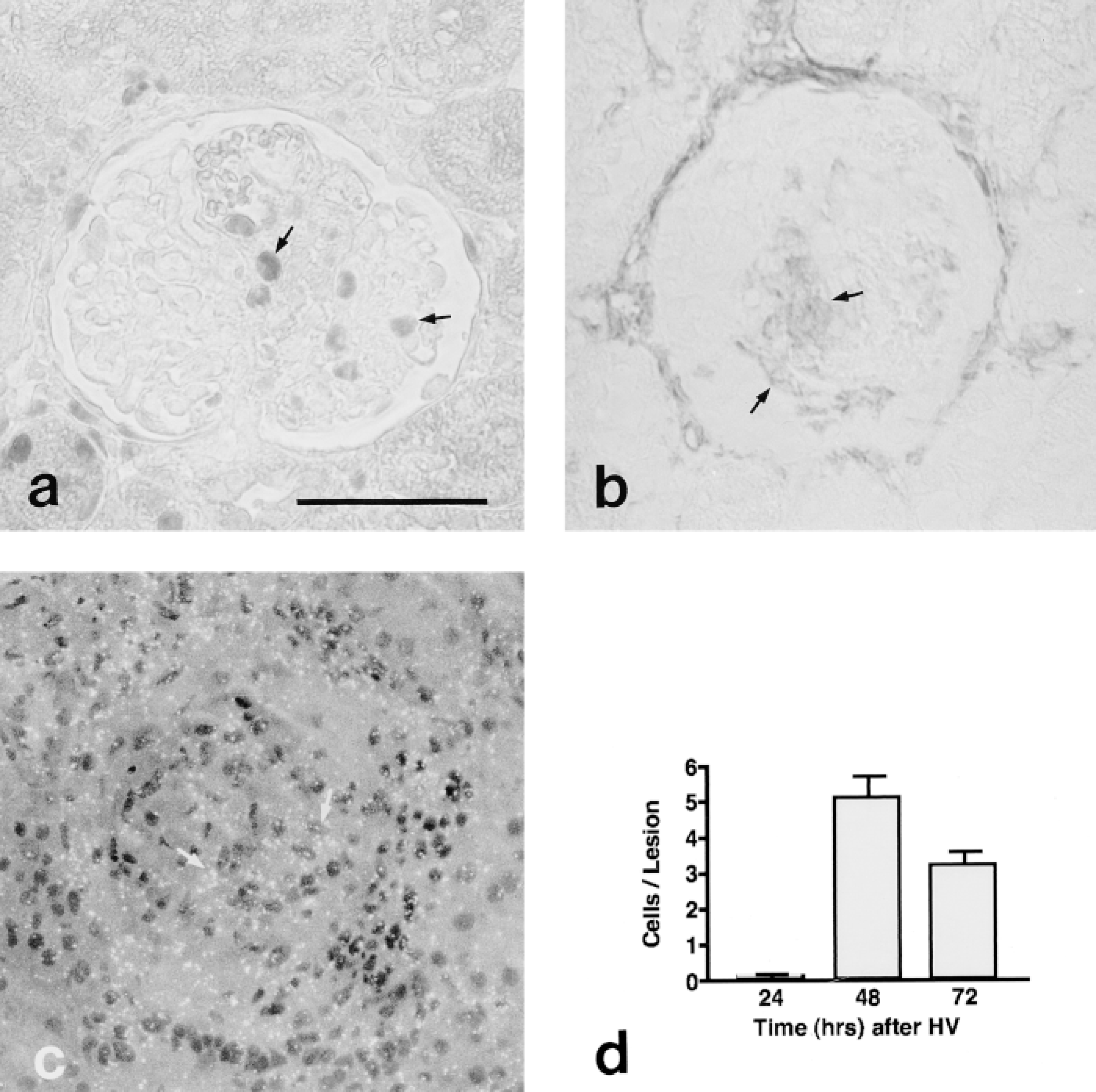
Assessment of proliferation in glomerular lesions. PCNA (a)-positive cells, α-SMA (b) (stained by immunoperoxidase histochemistry) and PDGFβ receptor (c) mRNA expression (in situ hybridization) were first observed in mesangial cells in the central aspects of glomerular lesions (arrows) 48 hr after HV. [3H]-Thymidine incorporation (d) peaked at 48 hr after HV (measured in tissue sections by autoradiography). Bar = 50 μm.
This study associates the temporal and spatial expression of TSP-1 and Fn-EIIIA with distinctive mesangial cell behaviors during remodeling over the course of experimental glomerulonephritis. However, specific functional roles for TSP-1 and Fn-EIIIA throughout the progression of experimental glomerular disease and their relevance to clinical forms of glomerulonephritis remain to be determined. Mesangiolysis can be found in a variety of human glomerulopathies, including malignant nephrosclerosis, renal transplant rejection, thrombotic thrombocytopenic purpura, hemolytic uremic syndrome, IgA nephropathy, snake bite-induced nephropathy, diabetic nephropathy, and several forms of glomerulonephritis (Morita and Churg 1983; Chugh 1989; Lee et al. 1989; Stout et al. 1993). The pathological mechanisms or evolution of mesangiolytic lesions in these diseases have not been defined. However, they may represent early stages of a continuum of mesangial proliferative and glomerulosclerotic lesions. A role for TSP-1 and Fn-EIIIA in progression of mesangiolytic lesions in humans remains to be determined.
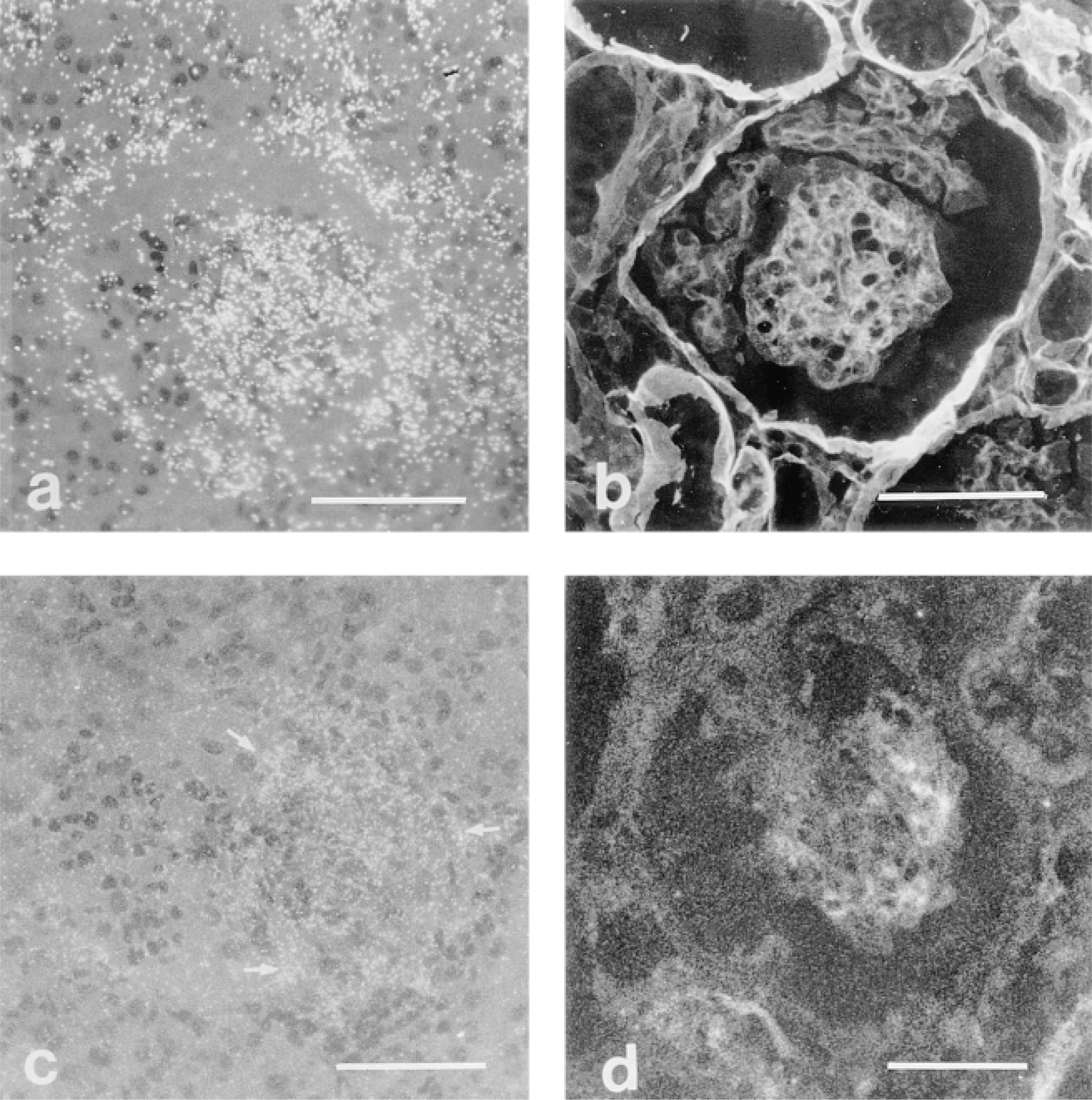
Assessment of ECM synthesis in glomerular lesions. Extracellular matrix mRNA and protein were most abundant at 72 hr after HV, as illustrated by in situ hybridization of collagen Type IV mRNA (a) and immunofluorescence of laminin protein (b). Extracellular matrix synthesis in micronodules was associated with expression of TGF β mRNA (arrows) by in situ hybridization (c) and protein by immunofluorescence microscopy (d) at this time. Bars = 50 μm.
Footnotes
Acknowledgements
Supported by NIH grants DK38758 from the National Institutes of Health (NIDDK) and by the Office of Research and Development, Medical Research Service, Department of Veterans Affairs.