
Abstract
The process of active nuclear protein transport is mediated by the nuclear localization signal (NLS). An NLS-containing karyophile forms a stable complex, termed the nuclear pore-targeting complex, to target nuclear pores. The α-subunit of the complex (importin-α) binds to the NLS and the β-subunit (importin-β-) carries the α-subunit, bound to the NLS substrate, into the nucleus. To date, five mouse α-subunits have been identified and classified into three subfamilies (α-P, α-Q, and α-S). The expression of these α-subunits and the β-subunit in various adult mouse tissues was examined by immunoblotting and immunohistochemistry using antibodies specific for each subfamily of the α-subunit or the β-subunit. The β-subunit was found to be ubiquitously expressed, whereas each subfamily of the α-subunit showed a unique expression pattern in various tissues, especially in brain and testis. In brain, the expression of α-P was not observed, whereas α-S was significantly expressed in Purkinje cells, and pyramidal cells of the hippocampus and cerebral cortex. In testis, α-P was expressed predominantly in primary spermatocytes, whereas α-Q was found mainly in Leydig cells. Expression of α-S was detected in almost all cells in convoluted seminiferous tubules and Leydig cells to a similar extent. These results suggest that nuclear protein import may be controlled in a tissue-specific manner by α-subunit family proteins.
Keywords
I
A number of α-subunit-related proteins from a variety of species have been identified using various biological screening techniques. Initially, we identified the 58-kD component of the PTAC from an extract of mouse Ehrlich ascites tumor cells, which was originally referred to as PTAC58 (Imamoto et al. 1995b,c). The α-subunit acts as an NLS receptor. There are two common structural features among this family of proteins. (a) They possess eight tandem repeats of 42-43 amino acids, which are rich in hydrophobic residues and are referred to as arm motifs. These repeats were originally identified in the product of a Drosophila segment polarity gene and were referred to as the armadillo protein (Peifer et al. 1994). (b) They contain about 40 amino acids which are rich in basic residues at the amino-terminal portion. These residues are required for binding to the β-subunit (Görlich et al. 1996; Weis et al. 1996). In human cells, three α-subunit family proteins have been identified, which were originally termed Rchl (Cortes et al. 1994; Cuomo et al. 1994), NPI1 (O'Neill and Palese 1995), and Qip1 (Seki et al. 1997), and which show about 50% amino acid identity with one another. However, in mouse cells, five α-subunit family cDNAs have been recently isolated from mouse Ehrlich ascites tumor cells (Tsuji et al. 1997). These are referred to as α-P (the human Rch1 counterpart and identical to PTAC58), α-S1 (the human NPI1 counterpart), α-S2, α-Q1 (the human Qip1 counterpart), and α-Q2. These proteins can be classified into three subfamilies, on the basis of their amino acid identities; α-P, α-Q, and α-S. These three subfamilies of the α-subunit show about 50% amino acid identity with one another, and individuals within the same subfamily show more than an 80% amino acid identity.
The β-subunit has been also identified in various species: Kap95 in yeast (Enenkel et al. 1995; Moroianu et al. 1995), PTAC97 in mouse (Imamoto et al. 1995a), and importin-β in human (Chi et al. 1995; Görlich et al. 1995). These β-subunit homologues bind directly to the α-subunit, NPC components, and small GTPase Ran (Gsp1/Gsp2 in yeast) (Chi et al. 1997; Kose et al. 1997; Kutay et al. 1997). It is known that the function of the β-subunit in nuclear import is to anchor the α-subunit, which is bound to the NLS substrate, to the NPC and to transport them into the nucleus.
Furthermore, it has been demonstrated that three distinct classes of α-subunits differentially recognize the NLSs (Miyamoto et al. 1997), which raises a new question as to why such a divergence of NLS receptors exists in cells. In addition, previous studies, using Northern blot analysis, have shown that the expression of various NLS receptors is controlled in both tissue-specific and cell type-specific manners at the mRNA level (Prieve et al. 1996; Nadler et al. 1997; Tsuji et al. 1997). However, the expression patterns of each α-subfamily protein have not yet been studied at the protein level, nor are any histochemical data available on these proteins. This article reports an examination of expression patterns of these three subfamilies of α-subunits in various adult mouse tissues by using specific antibodies. The study concludes that, whereas the β-subunit is ubiquitously expressed, each subfamily of the α-subunit shows a unique expression pattern in various tissues. These results support the hypothesis that the β-subunit plays a crucial role in nuclear protein transport and suggest that nuclear protein transport might be controlled in a tissue-specific manner by α-subunit proteins.
Materials and Methods
Animals and Cells
C57BL/6 strain mice, 4 weeks of age, were purchased from Nihon-SLC (Shizuoka, Japan). Mouse Ehrlich ascites tumor cells were passaged in mouse abdominal cavity and harvested using methods which have been described previously (Imamoto et al. 1995c). HeLa cells were cultured in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum.
Antibodies
Recombinant GST β-subunit, GST α-P, and GST α-Q1 proteins were prepared as described previously (Imamoto et al. 1995c). Instead of GST α-S1, a sufficient amount of which was not obtained, GST-NPI1, which is a human counterpart of mouse α-S1 with 97% amino acid identity (Tsuji et al. 1997), was prepared. Immunization of each recombinant protein was performed as described previously (Imamoto et al. 1995a,c). Specific antibodies were affinity-purified from antisera from each immunized rabbit by using a recombinant protein-conjugated Sepharose column for each antibody. In addition, anti-β-subunit, anti-α-P, and anti-α-Q antibodies were passed through a GST affinity column to deplete antibodies to the GST portion. Immunoblotting analysis confirmed that anti-α-S (anti-NPI1) or anti-α-Q antibody efficiently crossreacted with α-S1 and α-S2, or with α-Q1 and α-Q2, respectively, but not with other subfamily proteins. Anti-α-P antibody, which reacted strongly with α-P, crossreacted very slightly with α-S1 and α-S2.
Preparation of Cell Lysate for Immunoblotting
Animals were perfused through the left cardiac ventricle with PBS under pentobarbital sodium (0.1 g/kg body weight) anesthesia. Organs were then removed and homogenized with lysis buffer [5 mM HEPES (pH 7.3), 10 mM potassium acetate, 2 mM magnesium acetate, 2 mM dithiothreitol (DTT), 1 mM (p-amidinophenyl) methanesulfonyl fluoride hydrochloride (p-APMSF), and 1 μg/ml each of aprotinin, leupeptin, and pepstatin]. The lysates were centrifuged at 10,000 X g for 10 min at 4C and the supernatants were then collected as the cytosolic fractions. Total cytosol from Ehrlich ascites tumor cells was prepared as described previously (Imamoto et al. 1995c).
Immunoblotting
Sample proteins were denatured by boiling with sample buffer containing 2% sodium dodesylsulfate (SDS) and 100 mM DTT, separated by 10% SDS-PAGE, and then transferred electrophoretically to nitrocellulose membranes. The filters were incubated with 1.5 μg/ml of rabbit anti-β-sub-unit, anti-α-P, anti-α-S, and anti-α-Q affinity-purified antibodies in 20 mM Tris-HCl (pH 7.5) buffer containing 0.3 M NaCl and 3% skim milk for 12 hr at 4C after blocking with PBS containing 3% skim milk for 1 hr at room temperature (RT). Rabbit antibodies were detected with alkaline phosphatase-labeled goat anti-rabbit IgG antibody (Bio-Rad Laboratories; Hercules, CA) using standard protocols.
Immunocytochemistry
HeLa cells plated on the coverslips were washed with warm (37C) PBS and fixed with 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4) (PFA-PB) at 37C for 10 min. After washing, the cells were permeabilized with 0.5% Triton X-100 in PBS for 5 min at RT and blocked with 3% skim milk in PBS. Then the cells were incubated with affinity-purified antibodies (15-30 μg/ml) for 1 hr at RT. After extensive washing with PBS, the cells were incubated with FITC-conjugated goat antibodies to rabbit IgG (Biosource International; Camarillo, CA) for 1 hr at RT. The samples were examined with an Axiophot 2 microscope (Carl Zeiss Jena GmbH; Jena, Germany).
Immunohistochemistry
Animals were perfused through the left cardiac ventricle with 4% PFA at RT for 10 min under pentobarbital sodium (0.1 g/kg body weight) anesthesia. Tissues were postfixed in 4% PFA-PB for 12 hr or longer at 4C and washed successively with 2, 5, 10, 15, and 20% sucrose in PBS for 6 hr or longer. The samples were embedded with OTC compound (Miles; Elkhart, IN), frozen, and then sectioned in 10-μm-thick sections using a cryostat, Leica CM 1900 (Leica Microsystems Nussloch GmbH; Nussloch, Germany). The sections were then mounted on poly-L-lysine-coated slide glasses. After air-drying, the cryosections were blocked with 1% skim milk and 1% normal horse serum in PBS and the sections were then incubated with 15-30 μg/ml of affinity-purified antibodies or normal rabbit serum (as a negative control) independently for 12 hr at 4C. After extensively washing with PBS, the sections were treated with FITC-conjugated goat antibodies to rabbit IgG in PBS for 1 hr at RT. After washing with PBS, the sections were observed by a fluorescent microscope, Axiophot 2 (Carl Zeiss). The antibodies preabsorbed with respective immunogens (GST fusions) were used as controls. Each of the preabsorbed antibodies showed no specific staining patterns.
Results
Expression of α-subunit Subfamily Proteins and β-subunit in Various Tissues
Although it has been previously reported that α-subunit mRNA is expressed in a tissue-specific or celltype specific manner (Prieve et al. 1996; Nadler et al. 1997; Tsuji et al. 1997), very little is known about the tissue distribution of the polypeptides of these transport factors and their histological distribution in normal adult tissues. Therefore, we initially carried out immunoblotting analysis for three distinct classes of α-subunits and β-subunit. Affinity-purified antibodies against α-P, α-Q, α-S, and β-subunit were used for this purpose. The specificity of each antibody was tested by immunoblotting of the GST-cleaved recombinant proteins. As shown in Figure 1A, anti-α-P antibody reacted efficiently with recombinant α-P protein and crossreacted very slightly only with α-S1 and α-S2 proteins. This crossreactivity appeared to be negligible because the crossreactive bands were barely detectable, as shown in Figure 2B, Lane s. As shown in Figures 1B and 1C, anti-α-Q and anti-α-S antibodies recognized the corresponding subfamily proteins but not any of the other subfamily proteins.
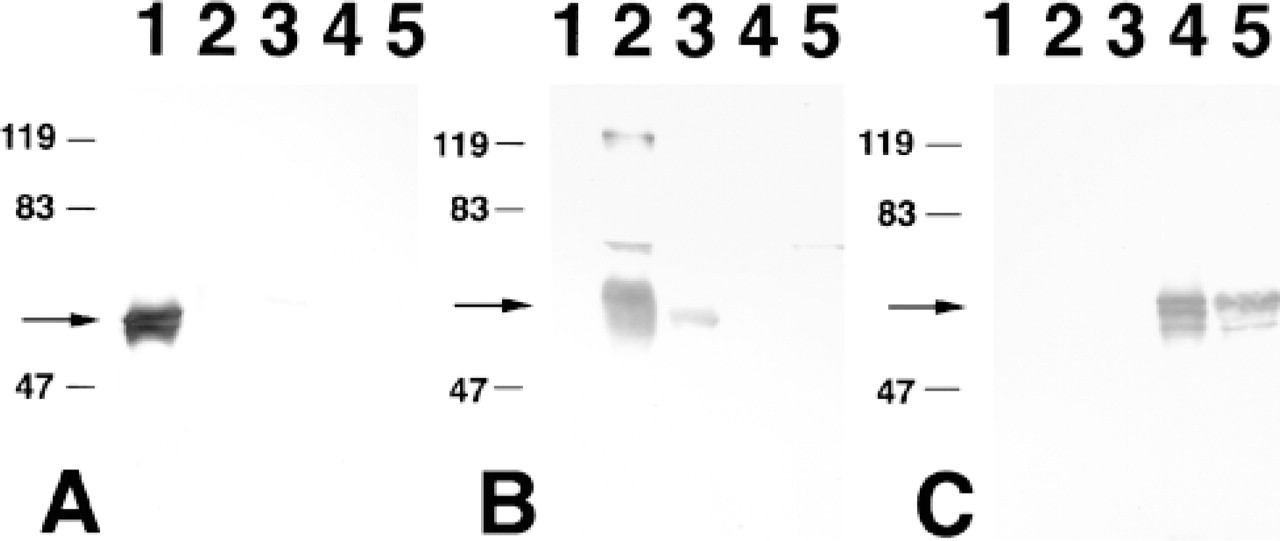
Specificity of antibodies raised for three subfamilies of importin-α subunits. Immunoblotting of recombinant α-P (Lane 1), α-S1 (Lane 2), α-S2 (Lane 3), α-Q1 (Lane 4), and α-Q2 (Lane 5) proteins with affinity-purified anti-α-P (A), anti-α-S (B), and anti-α-Q (C) antibodies was performed. One hundred ng of recombinant protein was applied to each lane. Crossreactive bands observed in Lanes 1 and 5 of B appear to be residual amounts of noncleaved GST α-subunit proteins which reacted with residual anti-GST antibodies in anti-α-S antibody. Arrows indicate the positions of each α-subunit. Molecular mass markers are indicated at left of each panel.
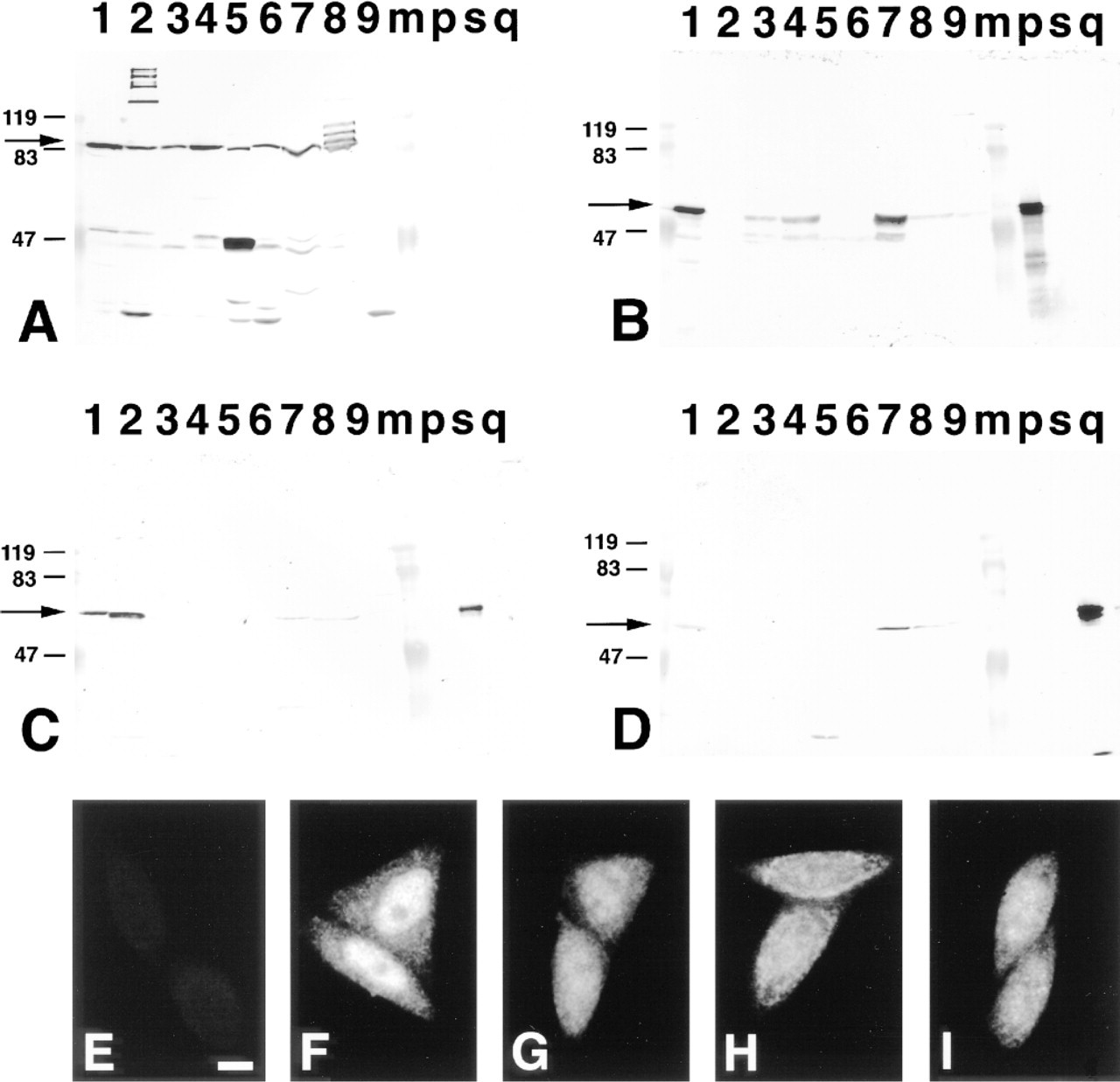
Immunoblotting of β-subunit and α-subunit subfamily proteins in various mouse tissues and their immunostaining patterns in HeLa cells. Ten μg of total cell lysates of mouse Ehrlich ascites tumor cells (Lane 1), brain (Lane 2), thymus (Lane 3), spleen (Lane 4), liver (Lane 5), kidney (Lane 6), testis (Lane 7), lung (Lane 8), and heart (Lane 9), and 100 ng of recombinant α-P (Lane p), α-S1 (Lane s) and α-Q1 (Lane q) proteins were applied to SDS-PAGE and transferred to nitrocellulose filters. The filters were immunoblotted by affinity-purified anti-β-subunit (A), anti-α-P (B), anti-α-S (C), and anti-α-Q (D) antibodies. Arrows indicate the positions of the β-subunit and each α-subunit. Molecular mass markers were applied to Lane m and appear at left of each panel (bold lines). HeLa cells were processed for immunofluorescence staining with affinity-purified anti-β-subunit (F), anti-α-P (G), anti-α-S (H), and anti-α-Q (I) antibodies, and normal rabbit serum as a negative control (E), as described in Materials and Methods. Bar = 10 μm.
By using these specific antibodies, it was possible to explore the expression of α-subunits and β-subunit in various tissues of the normal adult mouse. As shown in Figure 2, we found that the β-subunit is located in all tissues examined, whereas each α-subunit polypeptide exhibited a distribution pattern that was quite distinct. For example, α-P was not detected at all in brain, whereas α-S was significantly expressed and α-Q weakly expressed. In testis, all the α-subunit subfamilies were strongly expressed compared to other tissues. However, in liver and kidney, little or no expression of each α-subunit polypeptide was detected by the antibodies used here.
To identify the cell types in each tissue that express each α-subunit subfamily molecule and the β-subunit, we carried out an immunohistochemical analysis. First of all, to confirm that each antibody can detect the antigen immunocytochemically, we stained HeLa cells with these antibodies because it was previously reported that HeLa cells express three classes of the α-subunit family (Miyamoto et al. 1997). As shown in Figures 2E-2I, the β-subunit and all the classes of the α-subunit family were located not only in the cytoplasm but also in the nucleus, which is consistent with the previous reports (Imamoto et al. 1995a; Kose et al. 1997). As described above, the characteristic expression profiles of the α-subunit family were observed in the immunoblotting of the lysates from brain and testis. As a result, we focused on the brain and the testis for detailed analysis.
Expression of α-subunits and β-subunit Polypeptides in the Brain
In the cerebellum (Figure 3), the β-subunit protein was found in the cortex including the molecular layer, the Purkinje cell layer, and the granular layer, but not in the white matter (Figures 3C and 3D). As shown in Figure 2A, Lane 2, this anti-β-subunit antibody crossreacted with some extra bands in the brain extract. However, another lot of anti-β-subunit antibody, which cross-reacted with some extra bands different from those seen in Figure 2, also showed almost the same staining patterns as those seen in Figure 3 (not shown). On the basis of these results, we conclude that the signals obtained in Figure 3 indicate the expression of β-subunit in the brain. The α-P protein was not detected in any layer (Figures 3E and 3F), as expected, as evidenced by immunoblotting analysis. In contrast, the α-S protein was located predominantly in the cortex, especially in Purkinje cells (Figures 3G and 3H). Although the distributions of β-subunit and α-S were similar, α-S was prominently expressed only in Purkinje cells, which is a characteristic of α-S. In addition, we found that the α-S protein is located not only in the Purkinje cell soma but also in the dendrites. On the other hand, α-Q was only faintly detected and was diffusely distributed throughout the brain (Figures 3I and 3J).

Expression of β-subunit and α-subunit subfamilies in the cerebellum. Sections of the cerebellum were immunostained with affinity-purified anti-β-subunit (C,D), anti-α-P (E,F), anti-α-S (G,H), anti-α-Q (I,J) antibodies, and normal rabbit serum as a negative control (A,B). m, molecular layer; pu, Purkinje cell layer; g, granular layer; w, white matter. B, D, F, H, and J show higher-magnification views. Bars = 25 μm.
In the hippocampus (Figure 4), both the β-subunit protein (Figures 4C and 4D) and α-S protein (Figures 4G and 4H) were located in all regions. However, α-S protein was detected at much higher levels in the pyramidal cells than in other cells in this area. Moreover, the pyramidal cell neurites, which extend into the stratum radiatum, also showed positive staining, similar to their somata (Figures 4D and 4H). The expression patterns of α-P (Figures 4E and 4F) and α-Q (Figures 4I and 4J) in this area were similar to those in the cerebellum.
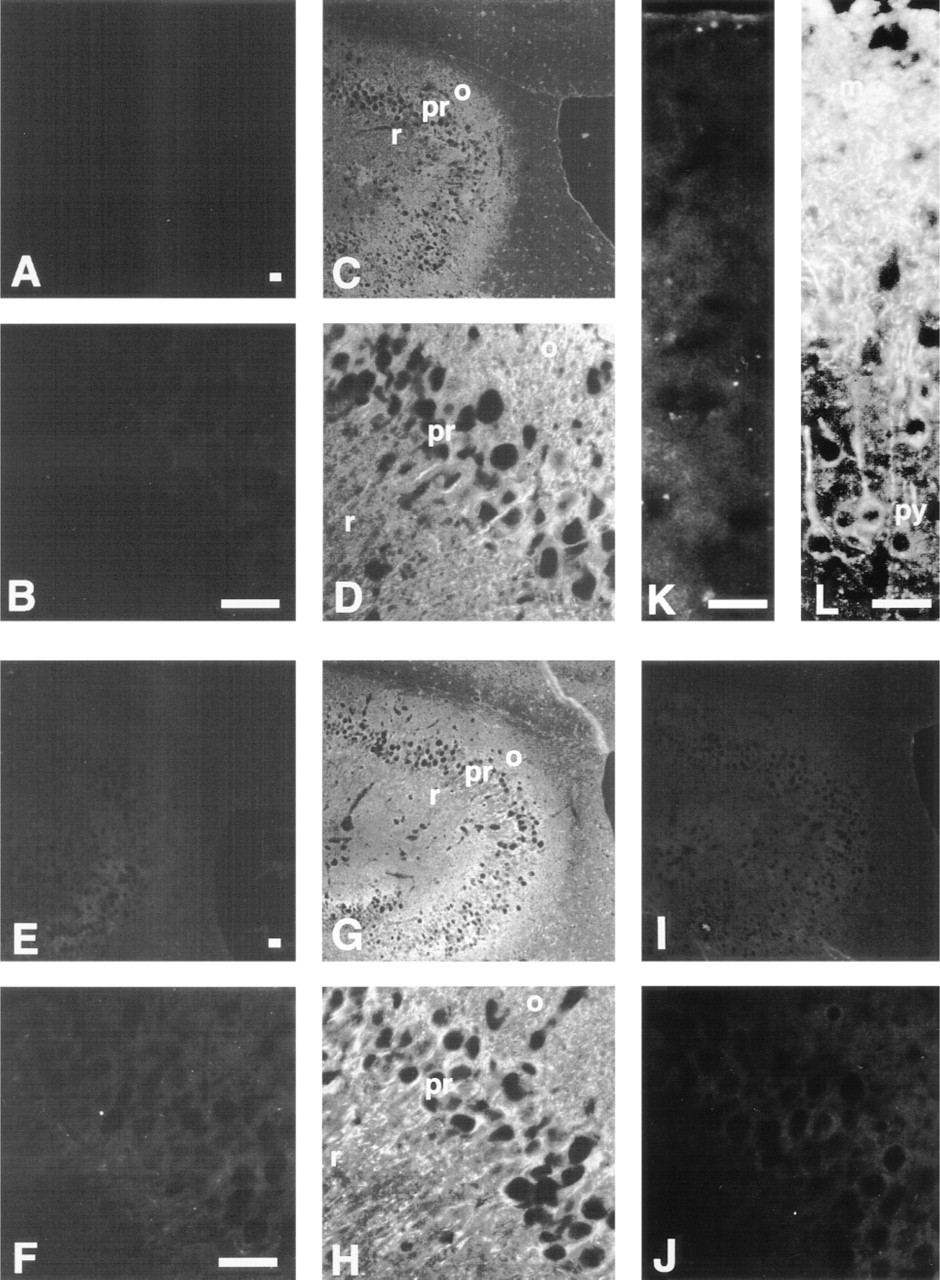
Expression of β-subunit and α-subunit subfamilies in the hippocampus (A-H) and in the cerebral cortex (K,L). Sections were immunostained with affinity-purified anti-β-subunit (C,D), anti-α-P (E,F), anti-α-S (G,H,L), anti-α-Q (I,J) antibodies, and normal rabbit serum as a negative control (A,B,K). r, stratum radiatum; o, oriens layer; py, pyramidal cell layer; m, molecular layer. B, D, F, H, and J-L show higher-magnification views. Bars = 25 μm.
In other areas of the brain, the localization of these α-subunit proteins was similar to that in the cerebellum and the hippocampus. For example, in the cerebral cortex, whereas α-P protein was not detected, the pyramidal cells and their neurites were strongly stained by the anti-α-S antibody (Figure 4L).
Expression of α-subunits and β-subunit Polypeptides in the Testis
The β-subunit protein was detected in all cells of the convoluted seminiferous tubules and in the interstitum (Figure 5B). In contrast, the α-P protein was detected predominantly in the primary spermatocytes and only slightly in other cells in the convoluted seminiferous tubules and the Leydig cells at the interstitum (Figure 5C). The α-S protein was located in all cells in the convoluted seminiferous tubules as well as in Leydig cells, similar to the β-subunit protein (Figure 5D). The α-Q protein was detected predominantly in the Leydig cells (Figure 5E). It is noteworthy that each of the α-subunit family molecules was strongly expressed in the testis but that the cells expressing them were different from one another.
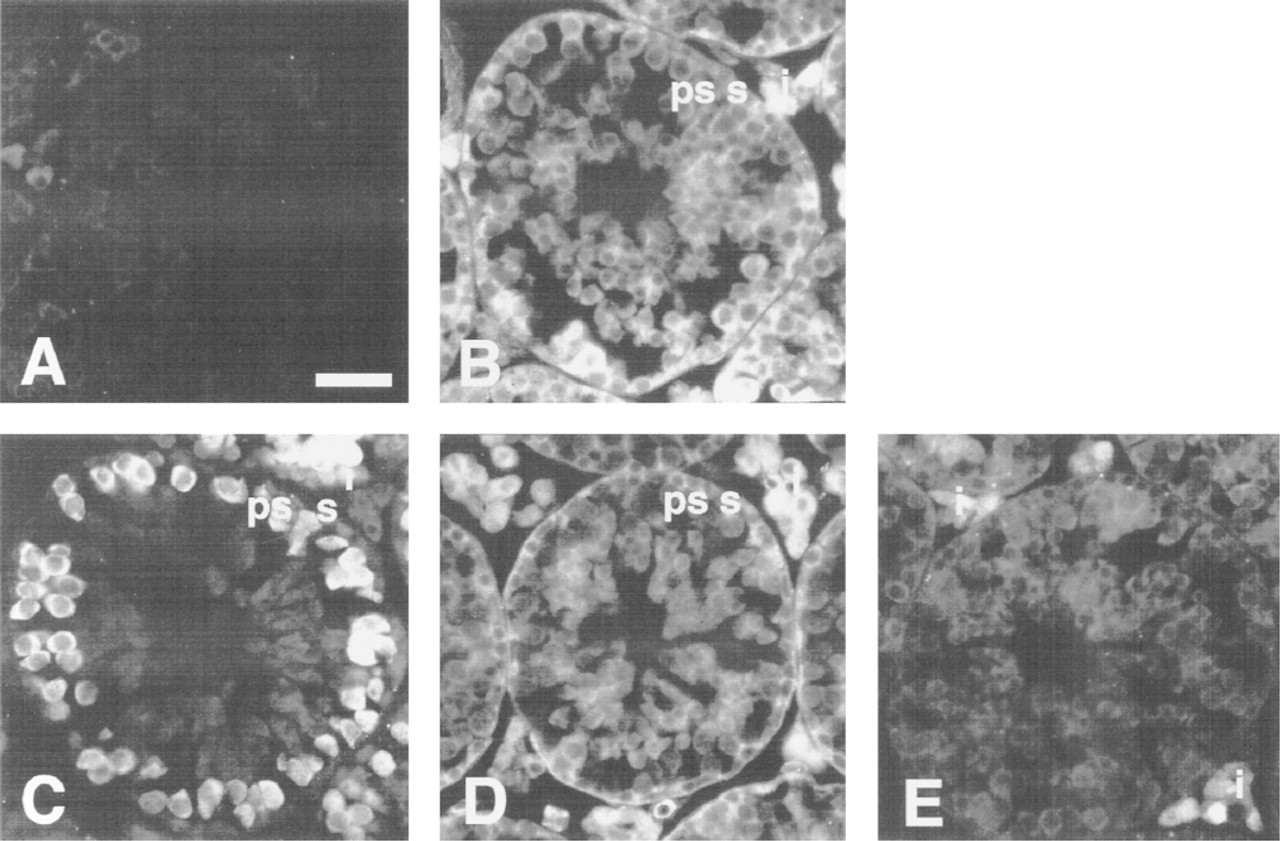
Expression of β-subunit and α-subunit subfamilies in the testis. Sections of testis were immunostained with affinity-purified anti-β-subunit (B), anti-α-P (C), anti-α-S (D), anti-α-Q (E), and normal rabbit serum as a negative control (A). ps, primary spermatocytes; s, spermatogonia; i, interstitial cells. Bar = 25 μm.
Discussion
The yeast Saccharomyces cerevisiae contains a single importin-α homologue, SRP1, which is an essential gene (Yano et al. 1992). In contrast, the α-subunit has been thus far identified in humans as three proteins and in mice as five. On the basis of sequence similarities and functional properties, these proteins can be classified into three distinct subfamilies. Why such a divergence of receptors exists in higher eukaryotes remains to be determined. On the other hand, the issue of whether nuclear protein transport is modulated in a tissue- or cell type-specific manner also remains unclear. Therefore, it is important to understand how the transport factors, importin-α family molecules and importin-β, are expressed in normal adult tissues. The present study approached this issue by examining the expression pattern of these polypeptides in normal mouse adult tissues via the use of specific antibodies.
We failed to produce five individual independent antibodies that react uniquely with individual α-subunit family molecules, probably because two members of each subfamily have a high degree of amino acid identity (more than 80%). However, we were successful in obtaining three distinct antibodies that recognize one subfamily of the α-subunit but not other subfamily members. By using these three specific antibodies, we were able to analyze the subfamily-specific expression pattern of the α-subunit.
Immunoblotting and immunohistochemical analyses indicate that all the subfamilies of the α-subunit were differentially expressed in a tissue-specific manner. The α-P protein was not detected in the brain nor in the highly concentrated cytosol and insoluble fraction of the brain homogenate (not shown), whereas a high expression of α-P protein was detected in the testis. These results are consistent with previous studies, based on Northern blotting of mRNAs from various tissues (Tsuji et al. 1997). In addition, the findings clearly show that α-P protein is located predominantly in the primary spermatocytes in the testis. Furthermore, the α-P protein was highly expressed in Ehrlich ascites tumor cells and in all the established tissue culture cells examined (not shown). Note that the primary spermatocyte is derived from the spermatogonium and proliferates actively before the start of meiosis. On the basis of these findings, we speculate that the α-P protein may be highly expressed predominantly in proliferating cells, suggesting that it may be mainly involved in the import of karyophilic proteins related to growth or cell division.
In contrast, the expression of the α-S subfamily proteins showed a characteristic pattern in the brain. The α-S proteins were strongly expressed in the cell somata and dendrites of Purkinje cells, in the pyramidal cells and their neurites of the hippocampus, and in the pyramidal cells and their neurites of the cerebral cortex. In this respect, it is noteworthy that Purkinje cells and the pyramidal cells receive input signals at their neurites from the molecular layer and transmit output signals to other regions along their axons. In a previous study, it was demonstrated that a transcription factor Stat1 (signal transducer and activator of transcription 1) which preexists in the cytoplasm in the resting state, is transported into the nucleus as the result of nuclear pore-targeting complex formation with NPI1, the human counterpart of mouse importin-α-S1, but not by Rch1, the human counterpart of mouse importin-α-P, when cells are stimulated by interferon-γ (Sekimoto et al. 1997). These findings suggest that although intracellular signal transduction pathways and their machinery in neurons are poorly understood, it is entirely possible that the α-S subfamily molecules are related to the signal transduction in neurons. If so, it would be interesting to determine which molecules are transported into the nucleus by the α-S proteins in individual neurons.
On the other hand, as shown in immunoblotting experiments, α-Q family proteins were expressed in many tissues, whereas their expression in the testis was observed predominantly in the Leydig cells. Human Qip1 was originally isolated as a binding molecule to DNA helicase Q1 by yeast two-hybrid method (Seki et al. 1997). Later, it was revealed that Qip1 efficiently recognizes the NLS of DNA helicase Q1 but not the NLSs of SV40 T-antigen and cap-binding protein 80 (Miyamoto et al. 1997). These results suggest that Qip1 may act as a novel NLS receptor, which has high recognition specificity for helicase Q1-NLS, although the possibility remains that Qip1 carries other untested karyophilic proteins into the nucleus. Therefore, it is possible that in tissues expressing α-Q proteins, a limited number of unique karyophilic proteins, such as helicase Q1, may be transported efficiently to the nucleus by α-Q proteins. It would be interesting to identify karyophilic proteins that are recognized by α-Q proteins in individual tissues.
By using several classes of α-subunit proteins which have different binding affinities for distinct types of karyophiles, we found that individual cells appear to be capable of importing various nuclear proteins. It is probable that individual cells transport nuclear proteins that are specific for their own function, as well as housekeeping karyophiles, which are essential for the life of every cell, into the nuclei by modulating a combination of α-subunit expression. Hence, it is likely that individual cells regulate the expression level of each α-subunit protein so as to efficiently and precisely transport the karyophilic proteins required for their own homeostasis into the nuclei.
On the other hand, importin-β polypeptides were found to be expressed more or less in all the tissues and in most of the cells in each tissue examined, although the expression level in the heart was rather low. The β-subunit is essential for nuclear pore-targeting complex formation and is able to bind to all classes of α-subunit proteins. Although several importin-β-related molecules, such as transportin (Pollard et al. 1996) and karyopherin-β3 (Yaseen and Blobel 1997), have been identified, it is only importin-β that carries α-subunit bound to a karyophile into the nucleus. Therefore, it is reasonable to suspect that the β-subunit protein is expressed more or less in all cells that express any of the α-subunits, which is consistent with our observations in this study.
This is the first report that shows the expression pattern of the nuclear import factors, importin-α family and importin-β, in normal adult tissues at the polypeptide level as well as histochemically. Such findings will aid in our understanding of whether different types of cells have unique modes for nuclear protein transport and raise an interesting question as to the existence of cell type-specific nuclear protein transport machinery that contributes to cell differentiation or the maintenance of differentiated phenotypes that are specific for each cell.
Footnotes
Acknowledgements
Supported by a Grant-in-Aid for Scientific Research on Priority Areas (No. 07282103) and a Grant-in-Aid for COE Research (No. 07CE2006) from the Japanese Ministry of Education, Science, Sports and Culture.
We wish to thank Naoko Imamoto for valuable discussion, and Tadanobu Ban, Toshihiro Sekimoto, Yoichi Miyamoto, and Jiyoong Kim for helpful advice.