
Abstract
Antithrombin is a serine protease inhibitor that is critical in maintaining a thromboresistant vasculature. The association between low serum antithrombin concentration and renal disease suggests that the kidney plays a role in the conservation of plasma antithrombin. We used immunohistochemical techniques to determine the spatial distribution, heparin binding characteristics, and intracellular and intercellular localization of antithrombin in biopsy specimens (n = 53) of human donor kidneys obtained at the time of transplantation. In the renal cortex, double antibody techniques demonstrated the presence of intracellular antithrombin in proximal tubule epithelial cells. The reactivity was granular and was co-localized with vesicle-like structures. Distal and collecting tubules did not demonstrate intraepithelial antithrombin reactivity. No tubule structures in the medullary region demonstrated intracellular antithrombin, but all these structures showed intense basement membrane antithrombin reactivity. Double antibody techniques also demonstrated that the heparin binding domain of intraepithelial antithrombin was occupied. Semiquantitative scores for intraepithelial antithrombin were significantly decreased in renal biopsy specimens obtained 30 min after anastomosis compared with biopsies from the same organ obtained before anastomosis. These findings suggest that antithrombin, probably in association with heparin or heparan sulfate, is internalized by renal proximal epithelial cells. Although the ultimate fate of intraepithelial antithrombin is not known, this may represent a mechanism by which the kidney helps to maintain plasma antithrombin concentrations.
A
We studied renal biopsy specimens taken at the time of transplantation to characterize the spatial distribution, heparin binding characteristics, and intracellular location of antithrombin in tubule epithelial cells of human kidneys. Our findings show that antithrombin, in association with heparin/heparan sulfate, is selectively localized in proximal tubule epithelial cells. This suggests that antithrombin is internalized by these cells and that this may represent a reabsorption mechanism to maintain plasma antithrombin concentrations by preventing urinary loss of filtered antithrombin.
Materials and Methods
Patients and Tissue Samples
Needle biopsies (n = 53) were obtained at the time of transplantation in patients (n = 44) who received renal allografts between 1991 and 1996. Biopsy material consisted of cortical (n = 41) and medullary (n = 12) specimens and all biopsies were obtained from transplanted kidneys within 30 min after anastomosis. In nine of the 44 patients, we received a biopsy taken from the donor kidney before reperfusion with recipient blood and another biopsy was taken 15-30 min after anastomosis was completed.
All patients were immunosuppressed with cyclosporin (10 mg/kg/day and tapered to 3-4 mg/kg/day) begun after renal function was established (creatinine < 4.0 mg/dl), azathioprine (3 mg/kg/day and tapered to 1.5 mg/kg/day), and prednisone (500 mg/day and tapered to 100 mg/day by Day 4). Maintenance prednisone was 10-25 mg/day. Unless contraindicated, all recipients of cadaveric allografts also received OKT3 induction therapy (5-10 mg/day) for 5-14 days. Patient demographics, HLA mismatches, total ischemic time, type of donor organ (cadaveric vs living related), organ preservation method (ice storage vs pump perfused), and pretransplant creatinine values were recorded.
Immunohistological Techniques
Antibodies and Control Experiments
The sources, specificities, and dilutions of all antibodies used are given in Table 1. Antibodies were centrifuged (100,000 × gat 4C) monthly to remove complexes and immunoglobulin aggregates. The dilution for each antibody was two serial dilutions before endpoint titration (Faulk and Johnson 1977). Negative controls included exposure of cryostat sections to PBS, primary or conjugate antibody only, or irrelevant primary antibody. Positive controls were obtained by reacting tissue sections with monoclonal antibody (MAb) to α-smooth muscle cell actin or MAb to platelet endothelial cell adhesion molecule-1 (PECAM-1).
Definition and characteristics of antibodies used in this study a

PCT, proximal convoluted tubules; DCT, distal convoluted tubules; CT, collecting tubules; AT, antithrombin; FITC, fluorescein isothiocyanate; RITC, rhodamine isothiocyanate.
Biopsies were embedded in optimal cutting temperature medium (Miles; Elkhart, IN), snap-frozen in liquid nitrogen, and stored at —20C. Cryostat sections (4 μm) were removed from the cryostat blade by flash condensation onto glass microscope slides and air-dried overnight at room temperature (RT). For single antibody experiments, the sections were washed in 10 mM PBS, pH 7.2, for 30 min, incubated for 20 min at RT with primary antibody, washed three times (10 min each) in PBS, and incubated for 20 min with fluorescein isothiocyanate (FITC)-conjugated second antibody.
To further characterize tubule antithrombin, double antibody experiments were performed using primary antibodies from different species and appropriately matched second antibodies (Table 1). Sections were incubated with primary mouse MAb to various different antigens followed by rhodamine (RITC)-conjugated goat anti-mouse IgG second antibody. The sections were then washed in PBS, incubated with rabbit polyclonal antibody to antithrombin, and reacted with FluoroBlue-conjugated goat anti-rabbit IgG. The polyclonal antibody against antithrombin produces a single line of identity by double immunodiffusion analysis with plasma or purified antithrombin (McKay 1980,1981). We have used this antibody extensively to assess changes in vascular antithrombin in cardiac and renal allografts (Absher et al. 1992; Bukovsky et al. 1992; Labarrere et al. 1992; Faulk and Labarrere 1993,1994; Torry et al. 1994; Torry et al. 1998) as well as antithrombin binding to endothelial cells in vitro (Justus et al. 1995).
To identify the spatial distribution of tubule antithrombin, a mouse MAb (URO3; clone F23) to an apical 140-kD membrane-associated protein of human proximal convoluted tubules (PCTs) and a mouse MAb (URO5; clone T16) to a 42-48-kD membrane protein of human distal convoluted tubules (DCTs) and collecting tubules (CTs) were used in double antibody experiments with rabbit antibody to antithrombin. The marker of proximal tubules (clone F23) gave the same reactivity patterns as an MAb to adenosine deaminase binding protein, known to be present on PCTs (Schrader and Pollara 1978; Tolkoff-Rubin et al. 1987). Specificity of the marker for distal and collecting tubules (clone T16) was confirmed by histochemical techniques on adjacent sections.
To determine if the heparin binding site of tubule anti-thrombin was occupied, mouse MAbs to the heparin binding domain of human antithrombin (MAb 2331) and to an epitope distant from the heparin binding domain (MAb 2333) were used. The MAb to the heparin binding domain of human antithrombin (MAb 2331) recognizes free anti-thrombin in human myocardial cells after infarction (unpublished observations). It also recognizes free antithrombin in solution by Western blot assay and ELISA. An antibody to megalin (gp330) was used to determine if this receptor complex co-localized with intracellular antithrombin. MAb to human Golgi zone apparatus was used in double antibody studies with antithrombin antibody to provide information regarding the intracellular distribution of tubule antithrombin.
Immunohistochemistry for T-cell, macrophage, and neutrophil infiltrates as well as vascular antithrombin was performed as described (Torry et al. 1994). All sections were coverslipped with aqueous mounting medium and the sections were studied by epi-illumination in a Leitz Orthoplan microscope fitted with interference optics. The microscope contained an HBO-100 mercury arc lamp and an epi-illuminator with I-2-type FITC, N2.1-type RITC, and FluroBlue filters. Photomicrographs were taken by using a Leitz camera and 35-mm Ektachrome film (ASA 200). To study the localization and distribution of antithrombin in double antibody experiments, multiple photomicrographs of the same area were taken by using different filters sequentially. Immunohistochemical reactivity patterns were evaluated from coded slides of equivalent magnification projected from the same distance in a darkened room. The antibody reactivity patterns were evaluated by three investigators experienced in renal histology and immunopathology.
Scoring of Antibody Reactivity
Semiquantitative scores for intracellular tubule antithrombin were established by using a 1 through 3 scoring system as described (Torry et al. 1998). Briefly, biopsies with 50-100% of PCTs demonstrating granular antithrombin immunoreactivity were assigned a score of 3.0; biopsies with 1-49% of tubules demonstrating granular antithrombin were given a score of 2.0; and biopsies devoid of granular tubule antithrombin immunoreactivity were given a score of 1.0. Semiquantitative scores for vascular antithrombin and cellular infiltrates were assigned as described (Torry et al. 1994). All scores were assigned by two experienced renal histologists/immunopathologists blinded to the origin of the biopsy specimens and subsequent graft function. Antibody reactivity scores were analyzed by the nonparametric Wilcoxon Rank Sum test. Statistically significant differences between groups were indicated if p < 0.05. All data are reported as the mean ± SEM.
Results
Control Experiments
No immunofluorescence was detected in sections exposed only to PBS, primary antibodies, or conjugated second antibodies alone, or to irrelevant isotype-matched primary antibodies. Normal reactivity patterns were detected in positive control experiments that used MAbs to either α-smooth muscle actin or PECAM-1 to label vascular smooth muscle cells and endothelial cells, respectively.
Antithrombin in the Renal Cortex (Table 2)
Tubule Antithrombin
Antithrombin was identified in epithelial cells of tubules that morphologically resembled PCTs as well as in the basement membrane of tubules that morphologically resembled DCTs. The intraepithelial immunoreactivity was granular. The precise localization of granular intraepithelial antithrombin to PCTs was determined by using double antibody techniques with an MAb to an epitope on PCTs. These experiments demonstrated that granular intraepithelial antithrombin reactivity was confined to PCT epithelium and that these tubules lacked basement membrane antithrombin reactivity (Figure 1). With double antibody techniques with an MAb to an epitope on distal and collecting tubules, epithelial cells of these structures lacked granular intraepithelial antithrombin immunoreactivity. However, many of these tubules demonstrated intense basement membrane antithrombin immunoreactivity (Figure 2).
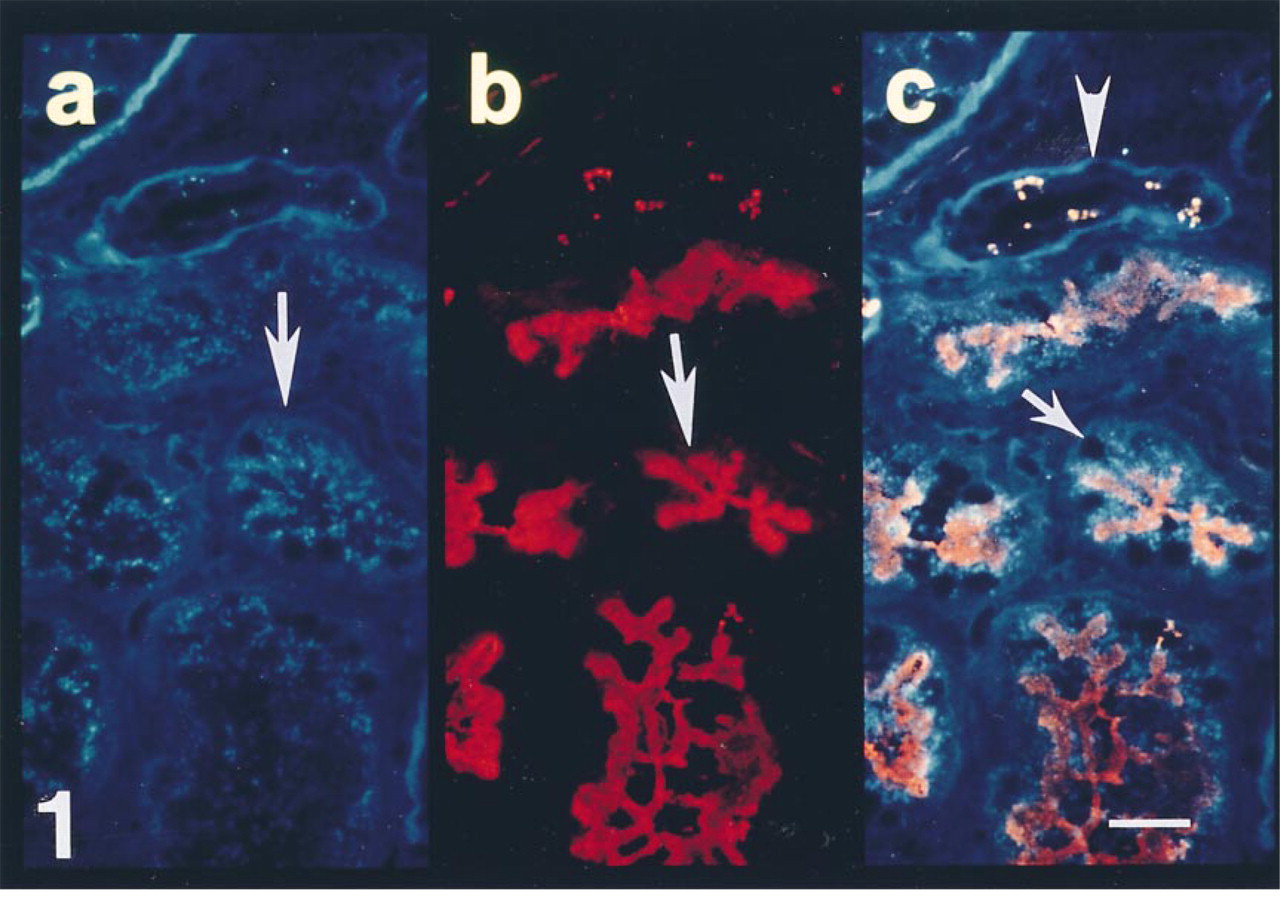
Section from a biopsy of a cadaveric renal allograft taken 30 min after reperfusion with recipient's blood. (a) Polyclonal antibody to antithrombin demonstrates granular immunoreactivity in the epithelial cells of some tubules (arrow). (b) MAb to proximal convoluted tubules labels the apical aspects of the epithelium (arrow). (c) Double exposure photomicroscopy demonstrates a high density of intracellular antithrombin in all epithelial cells of proximal tubules (arrow). Note the lack of intraepithelial antithrombin reactivity in adjacent tubules that are not labeled by the MAb to proximal tubules (arrowhead). Bar = 10 μm.
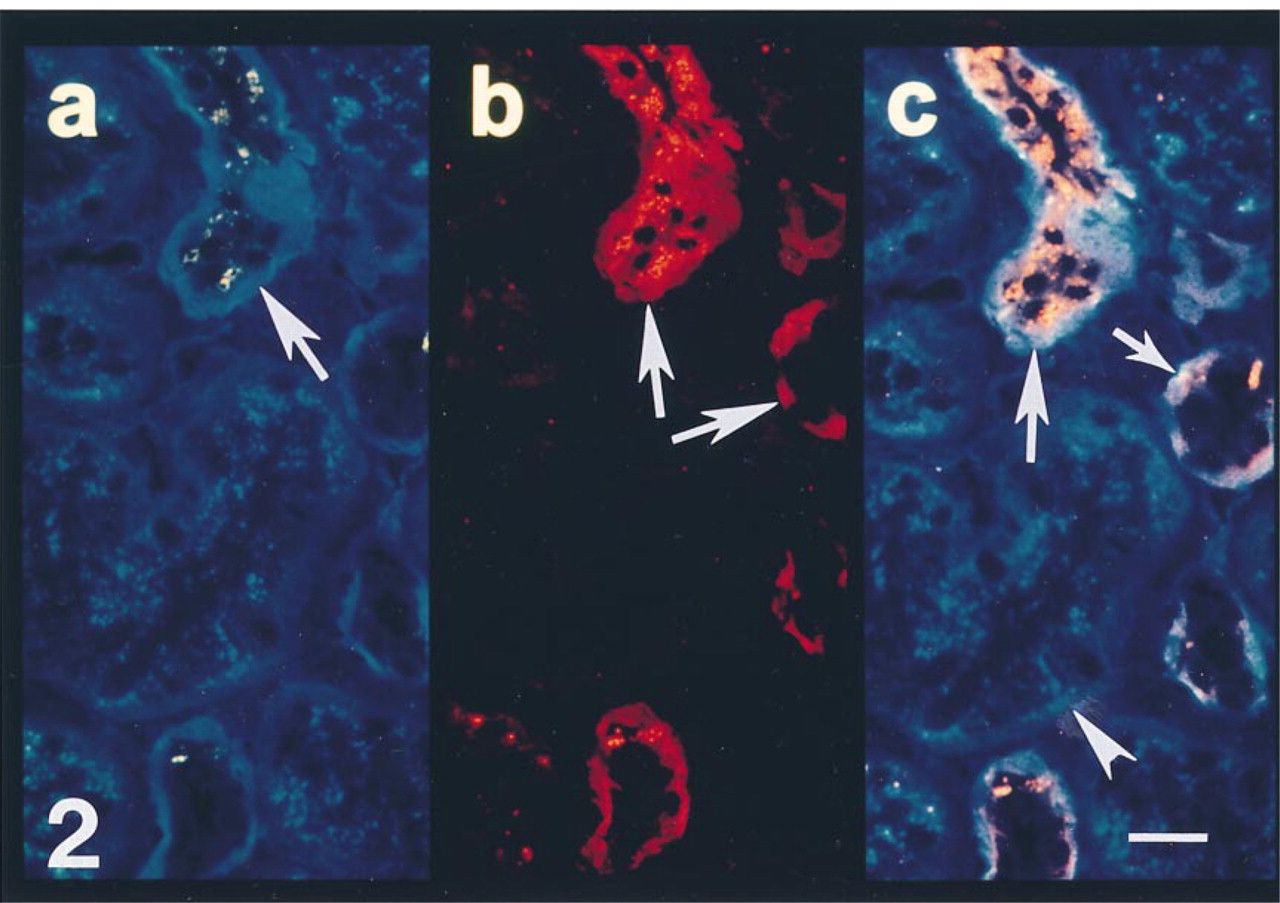
Section from the same biopsy shown in Figure 1. (a) Polyclonal antibody to antithrombin demonstrates lack of granular immunoreactivity in the epithelium of some tubules (arrow). (b) MAb to an epitope on distal and collecting tubules labels the epithelial cells of some tubules (arrow). (c) Double exposure photomicroscopy demonstrates the lack of intraepithelial antithrombin reactivity in distal/collecting tubules (arrow) but the presence of intracellular antithrombin in epithelial cells of adjacent tubules (arrowhead). Bar = 10 μm.
Summary of antithrombin distribution in human kidneys a

PCT, proximal convoluted tubules; DCT, distal convoluted tubules; CT, collecting tubules.
Vascular Antithrombin
Renal sections reacted with a polyclonal antibody to antithrombin produced characteristic reactivities in smooth muscle cells of arteries and arterioles, arterial intima, and with venous endothelium, as previously reported (Absher et al. 1992; Torry et al. 1994). Capillary antithrombin reactivity was generally present only in microvasculature bordering distal tubules that lacked intraepithelial antithrombin reactivity, whereas capillaries surrounding PCTs that contained granular intraepithelial antithrombin were consistently antithrombin-negative.
Antithrombin in the Renal Medulla (Table 2)
Tubule Antithrombin
Unlike proximal tubules, the medullary loops of Henle and collecting tubules did not demonstrate granular intraepithelial antithrombin reactivity. However, they all demonstrated intense basement membrane antithrombin reactivity (not shown).
Vascular Antithrombin
All the capillaries of the medulla demonstrated antithrombin reactivity. Similar to the cortex, smooth muscle cells of arteries and arterioles, arterial intima, and venous endothelium in the medulla also demonstrated antithrombin immunoreactivity (not shown).
Characterization of Proximal Tubule Antithrombin
To characterize the heparin binding characteristics of intraepithelial antithrombin, double antibody experiments were performed with MAbs that recognize either the heparin binding site or an epitope distant from the heparin binding site of human antithrombin. Double antibody techniques using the polyclonal antibody and monoclonal antibody to the non-heparin binding site of antithrombin showed co-localization on the intraepithelial granules of PCTs, basement membrane of DCT/CTs, and on the microvasculature (Figure 3). In contrast, there was no detectable immunoreactivity in any of these structures with the MAb against the heparin binding site of antithrombin (Figure 4). Results from the use of this antibody, which recognizes free antithrombin in solution by Western blot analysis and ELISA (not shown), suggests that the heparin binding site of antithrombin in PCTs, tubule basement membranes, and microvasculature might be occupied.
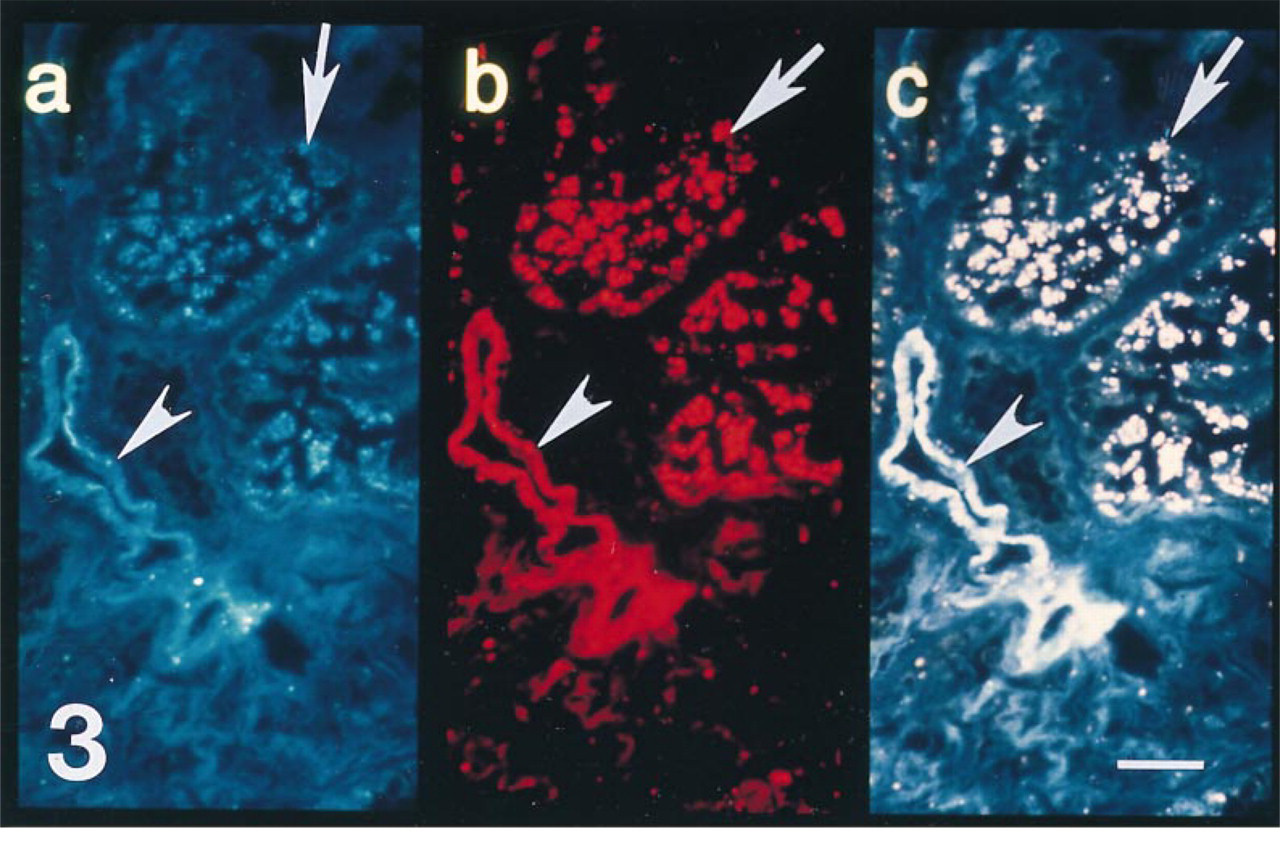
Section from a biopsy of a cadaveric allograft taken 30 min after reperfusion with recipient blood. (a) Polyclonal antibody to antithrombin labels tubule (arrow) and vascular (arrowhead) structures. (b) MAb that recognizes an epitope on human antithrombin that is distant from the heparin binding site of antithrombin also labels tubule (arrow) and vascular (arrowhead) structures. (c) Double exposure photomicroscopy clearly demonstrates that both antibodies co-localize (producing a pink signal) on intraepithelial antithrombin (arrow) and antithrombin associated with the microvasculature (arrowhead). Bar = 10 μm.
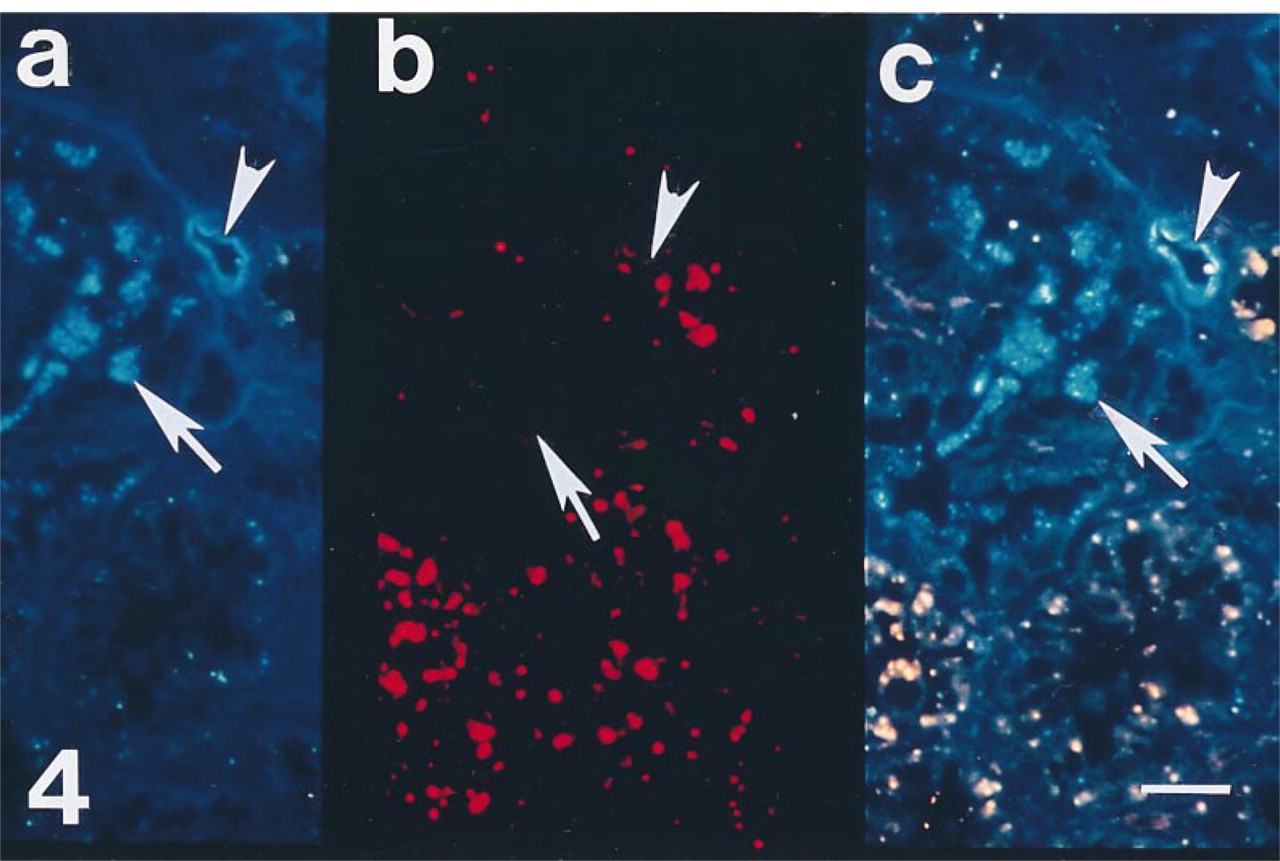
Section from a biopsy of a cadaveric allograft taken 30 min after reperfusion with recipient blood. (a) Polyclonal antibody to antithrombin labels tubule (arrow) and vascular structures (arrowhead). (b) MAb that recognizes the heparin binding site on human antithrombin fails to demonstrate reactivity in tubule (arrow) or vascular structures (arrowhead). (c) Double exposure photomicroscopy demonstrates only immunoreactivity of the polyclonal antibody in proximal tubules (arrow) and microvasculature (arrowhead), indicating that the heparin binding site of intraepithelial and vascular antithrombin may be occupied. Bar = 10 μm.
The intraepithelial distribution of antithrombin was further assessed by double antibody techniques with an MAb to human Golgi zone apparatus. The intracellular antithrombin immunoreactivity in PCTs co-localized with structures recognized by the Golgi zone MAbs, and no antithrombin immunoreactivity was associated with similar structures of DCTs or CTs (Figure 5).
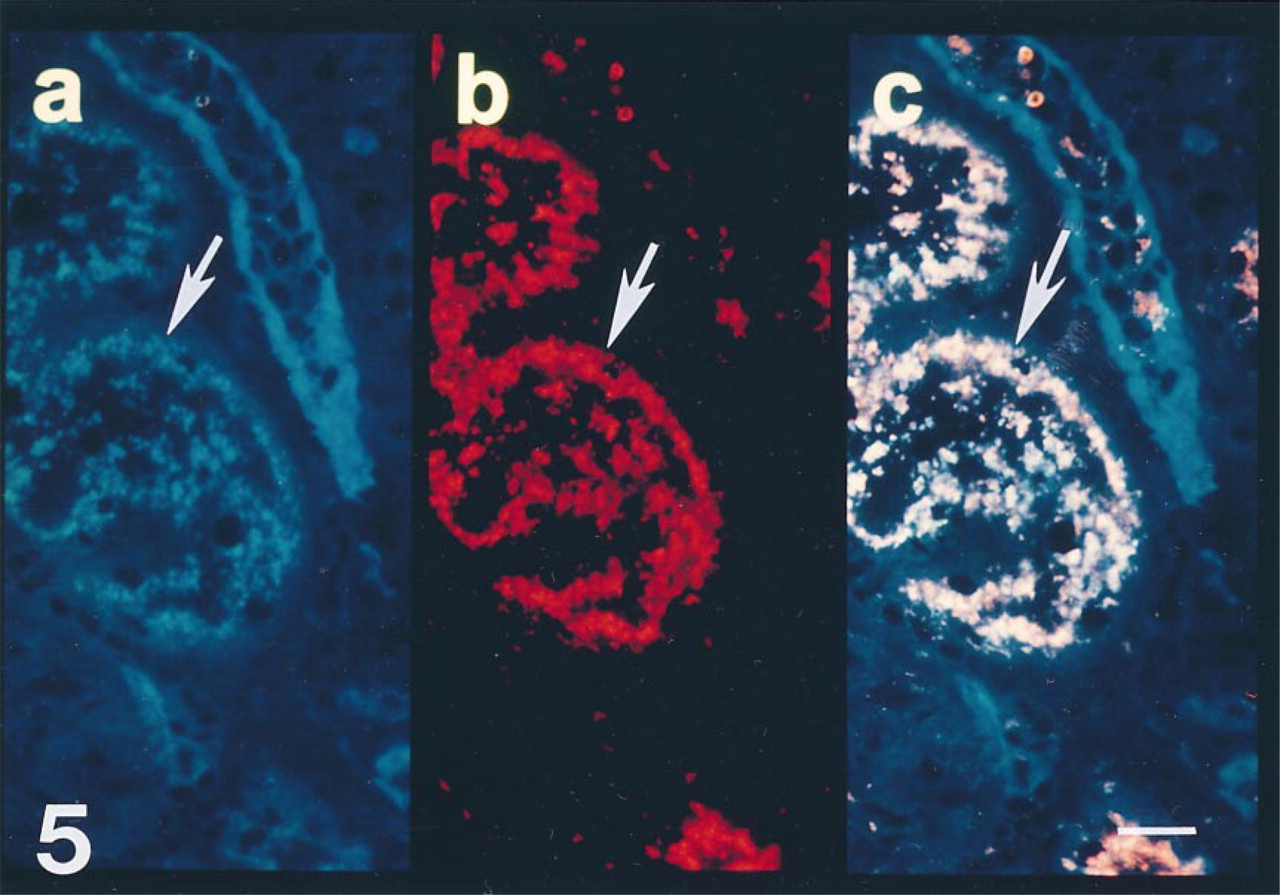
Section from a biopsy of a cadaveric allograft taken 30 min after reperfusion with recipient blood. (a) Polyclonal antibody to antithrombin demonstrates reactivity in epithelial cells (arrow). (b) MAb that recognizes human Golgi zone apparatus also demonstrates intense intraepithelial reactivity (arrow). (c) At this resolution, double exposure photomicroscopy demonstrates that both antibodies co-localize (producing a pink signal) on intraepithelial structures (arrow). Bar = 10 μm.
The potential mechanism for antithrombin entrance into vesicular structures of proximal tubules was assessed using double antibody techniques with antibody to gp330. This receptor complex is confined to proximal tubule brush borders and internalizes other protease inhibitors (Christensen et al. 1992; Moestrup et al. 1993). However, we could not demonstrate an association between gp330 reactivity and antithrombin reactivity in proximal tubules (data not shown).
Paired biopsies from nine patients provided the opportunity to assess temporal changes in tubule anti-thrombin before and after graft anastomosis (Table 3). Semiquantitative scores for tubule antithrombin were decreased in biopsies taken within 30 min of anastomosis compared to biopsies taken before reperfusion. These changes were not due to hyperacute rejection because there were no differences in cellular infiltrates between pretransplant and immediate posttransplant biopsies (Table 3).
Differences in semiquantitative antibody scores between biopsies from donor kidneys before reperfusion and from the same allograft within 15-30 min of reperfusion
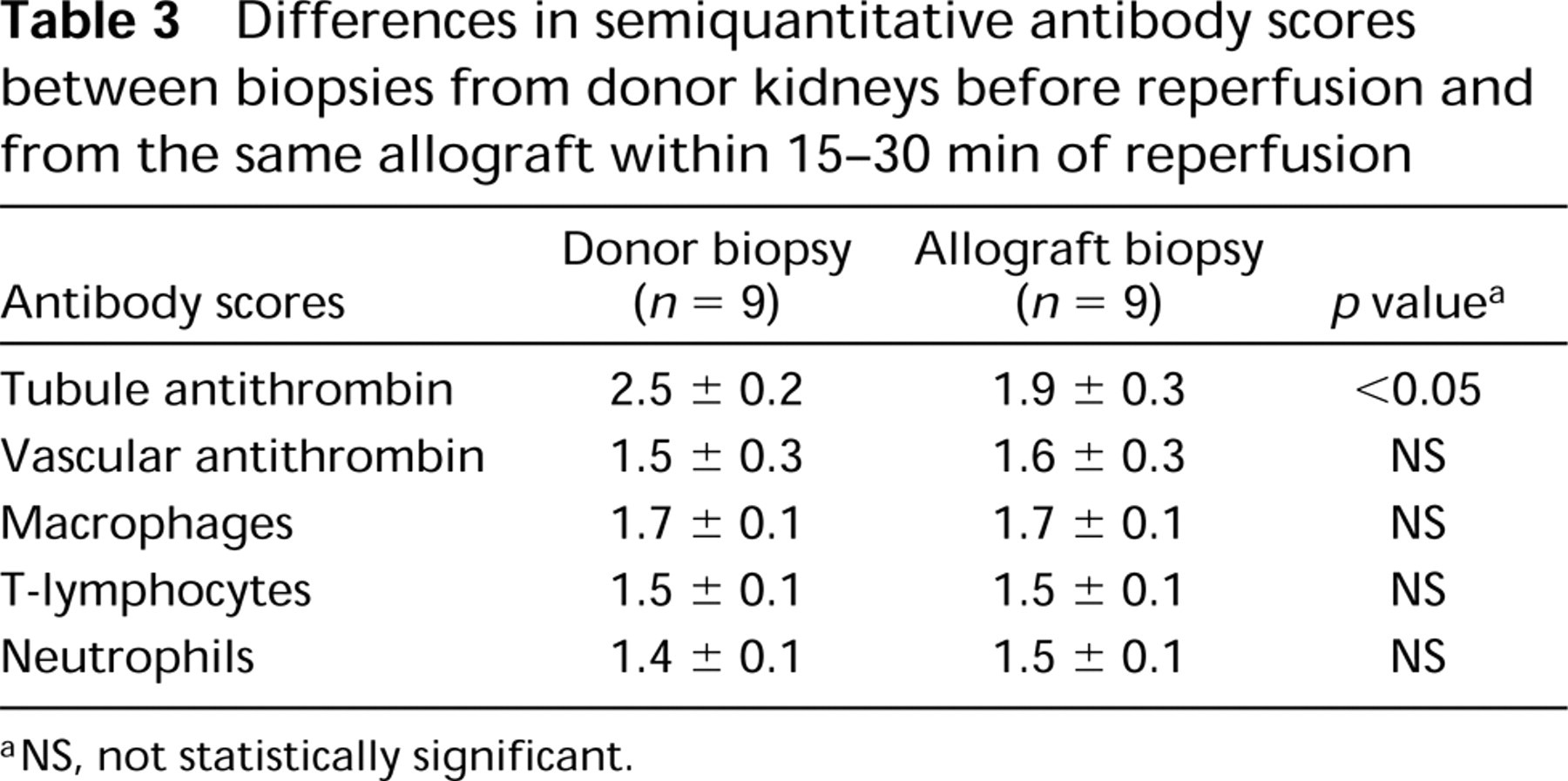
NS, not statistically significant.
Discussion
This study demonstrates the presence of intracellular antithrombin in renal epithelial cells of human kidneys. Intraepithelial antithrombin is confined to cells of proximal tubules, the heparin binding domain of antithrombin is occupied, and intraepithelial anti-thrombin is associated with intracellular structures. To our knowledge, this is the first study to describe the spatial distribution and characteristics of intracellular tubule antithrombin in human kidneys.
The spatial compartmentalization of tubule anti-thrombin in normal human kidneys was consistent. Proximal tubules with prominent intraepithelial anti-thrombin reactivity always lacked basement membrane antithrombin reactivity, and the capillaries surrounding these tubules lacked antithrombin immunoreactivity. However, distal/collecting tubules and all tubule structures in the medulla lacked intraepithelial anti-thrombin, and the vast majority demonstrated intense basement membrane antithrombin reactivity. In addition, capillaries surrounding distal and collecting tubules were usually antithrombin-reactive.
The biochemical mechanisms and physiological significance of these differences in the spatial distribution of tubule antithrombin in human kidneys are not known. However, we have shown differences in the spatial distribution of vascular antithrombin in the microvasculature of human hearts (Labarrere et al. 1992; Faulk and Labarrere 1993,1994) and kidneys (Bukovsky et al. 1992; Absher et al. 1992; Torry et al. 1994), and these alterations are predictive of graft failure (Faulk and Labarrere 1993).
Our present findings show that the heparin binding site of intraepithelial antithrombin is occupied, presumably by heparin and/or heparan sulfate. In contrast, only a fraction of antithrombin in plasma is complexed with heparin/heparan, because measurements of clotting times are significantly prolonged by exogenous heparin (Hirsh 1991). The source of endogenous heparin/heparan complexed with intraepithelial antithrombin is not presently known, but two possibilities are that filtered antithrombin is internalized as an already complexed molecule or that it becomes complexed with heparin/heparan produced by proximal tubule epithelial cells. Because urinary loss of heparan sulfate proteoglycan correlates with proteinuria immediately after renal transplantation (Stefanidis et al. 1996), complexed antithrombin may be internalized from the glomerular filtrate by a receptor-mediated process. Proximal tubule epithelial cells express megalin (gp330) at the base of the microvilli (Christensen et al. 1992), and this receptor mediates endocytosis of many proteins, including other complexed protease inhibitors (Moestrup et al. 1993). However, we could not demonstrate an association of this receptor with intraepithelial antithrombin. The association of tubule antithrombin with other proteins shown to be involved in proximal tubule reabsorption, such as gp280 (Verroust et al. 1996), needs to be investigated.
Internalization of tubule antithrombin appears to be an active process because our previous data show that allografts with delayed function were characterized by a reduction in intraepithelial tubule antithrombin at the time of transplantation (Torry et al. 1998). In the present study, we found a slight but statistically significant reduction in tubule antithrombin immediately after reperfusion. These findings are specific for antithrombin because earlier immunohistochemical studies did not identify other serine protease inhibitors such as α2-plasmin inhibitor (Torry et al. 1994) or α1-anti-trypsin (unpublished observations) in renal epithelial cells at the time of transplantation. We have found similar antithrombin reactivity patterns in renal tissue taken from rats and immediately processed for immunohistochemistry (not shown), suggesting that the presence of intraepithelial antithrombin is not due to cell damage secondary to donor organ harvest, preservation, and reperfusion associated with transplantation. Others have demonstrated similar vascular patterns of antithrombin reactivity in rat kidneys (Xu and Slayter 1994).
Certainly, other explanations for intracellular antithrombin, such as proximal epithelial synthesis of the molecule, are possible. However, by Northern blot analysis, rabbit kidneys do not express antithrombin transcripts (Sheffield et al. 1992). Nevertheless, cells from a monkey kidney epithelial cell line synthesize a molecule capable of binding and neutralizing thrombin (Morganstern et al. 1993), and other intraepithelial serine protease inhibitors have been reported in different cell types (Kopitar et al. 1985; Remold-O'Donnell et al. 1989). Their physiological significance is not known.
In kidneys, filtered antithrombin may help to prevent clot formation in tubule lumens, because we occasionally observe luminal fibrin in dysfunctional renal allografts (unpublished observations). Intraepithelial antithrombin may be trafficked via a heparin/heparan-mediated process through epithelial cells of proximal tubules as a recycling mechanism for filtered antithrombin, or it may be brought into the cells to serve as an intracellular protease inhibitor. Although the exact subcellular mechanism is not known, the apparent co-localization of intraepithelial antithrombin and vesicle-like structures of the Golgi apparatus imply an orderly trafficking of intracellular antithrombin. Indeed, endocytic trafficking of substances to the Golgi apparatus in various cell types has recently been described (Pavelka et al. 1998).
Regardless of the mechanism(s) involved, intraepithelial antithrombin apparently is tightly bound. Results of displacement/replacement and enzymatic experiments using heparin, heparan sulfate, or heparinase showed that vascular, but not intraepithelial, antithrombin, can be removed from sections of human kidney (Absher et al. 1992).
Although the fate of intraepithelial antithrombin in proximal tubules is not known, the physiological importance of tubule antithrombin is supported by significant reductions in plasma antithrombin concentrations in patients with end-stage renal disease (Vaziri et al. 1994) and by low serum antithrombin concentrations in patients with nephrotic syndrome (Vaziri et al. 1984; Farkas et al. 1993) and in animal models of induced nephrotic syndrome (Cruz et al. 1994). Indeed, the relatively short antithrombin half-life of 1.86 days (Hatton et al. 1997) suggests that the kidneys may help to conserve high plasma antithrombin concentrations. Others have shown that increased plasma antithrombin activity during development is not regulated at the transcriptional level in liver (Niessen et al. 1996), suggesting that other mechanisms in addition to liver synthesis are critical in regulating plasma antithrombin activity. Proximal tubule involvement in conserving other abundant plasma protein concentrations is well known. Indeed, relatively high quantities of filtered albumin are reabsorbed almost exclusively in proximal tubules (Gekle 1998), thus preventing urinary loss of albumin. Our present immunohistochemical findings, coupled with our clinical associations (Torry et al. 1998), support the hypothesis that proximal tubule reabsorption of antithrombin may provide a specific mechanism to prevent urinary loss of antithrombin.
Footnotes
Acknowledgements
Supported by The National Kidney Foundation of Indiana (RJT), Indiana Affiliate of The American Heart Association (RJT) and Novo Nordisk Pharmaceuticals (Copenhagen, Denmark).
We thank Denise Miller for technical support, and Dr Charles Carter, Dr Brian Haag, Nancy Duncan, RN, Christy Null, RN, and Kim Fox, RN of the Methodist Transplant Department for their assistance. The anti-megalin antibody was a generous gift from Dr Dennis Brown (Massachusetts General Hospital and Harvard Medical School, Boston, MA).