
Abstract
Cytoskeletal proteins associate with specific cell adhesion complexes and membrane proteins and influence the structural and functional organization of polarized epithelial cells in the kidney. Among such proteins that have been studied in cultured cell lines and in animals are the tight junction complex (ZO-1 and occludin), the adherens cell-cell adhesion complex (α-, β-catenin and plakoglobin), and Na+,K+-ATPase, with its associated membrane skeleton proteins ankyrin and fodrin. Although abnormal distribution of these proteins has been implicated in the pathogenesis of various renal diseases, the relevance of these findings to corresponding disease of the human kidney remains to be established. As a first step towards elucidating a role for such proteins in human kidney disease, we undertook a histochemical analysis of the distribution of these proteins in biopsy specimens of human kidney taken from healthy kidney transplant donors. We found each protein to have a characteristic subcellular localization and an intensity of staining that varied among different segments of the nephron in a manner that is consistent with discrete, segmental nephron function.
E
For proper function of renal epithelial cells, ion channels, exchangers, and transporters must be localized to functionally and structurally distinct plasma membrane domains, termed apical and basolateral, that face the nephron lumen and interstitial space, respectively (Almers and Stirling 1984; Fish and Molitoris 1994). Studies have shown that both cell-cell and cell-substrate contacts are required to initiate formation of these plasma membrane domains (Drubin and Nelson 1996). Subsequently, targeting to and retention of proteins in these domains establishes and maintains the functional polarity of these cells (Rodriguez-Boulan and Powell 1992; Drubin and Nelson 1996). Cytoskeletal proteins play essential roles in the development and maintenance of structural and functional organization of polarized epithelial cells. Significantly, recent studies of several kidney diseases show changes in the polarized distribution of membrane and cytoskeletal proteins (Fish and Molitoris 1994).
The tight junction complex is composed of several cytoplasmic and membrane proteins, including zonula occludens-1 (ZO-1) and occludin, and is localized to the boundary of the apical and basolateral plasma membrane domains of polarized transporting epithelial cells (Mitic and Anderson 1998). The tight junction complex forms a primary barrier to the passage of solutes through the intercelluar space (gate function), and the segregation of membrane proteins to apical and basolateral membrane domains (fence function). It has been reported that ATP depletion of cultured kidney cells results in a loss of tight junction complexes at their apical intercellular sites and apical junctional actin cytoskeleton, leading to a loss of gate function (Mandel et al. 1993; Bacallao et al. 1994). Ischemia-induced loss of epithelial cell polarity (impaired gate and fence function) has also been described in rats (Molitoris et al. 1989).
The E-cadherin/catenin adherens junction complex plays an important role in cell-cell adhesion, signal transduction, and the initiation and maintenance of structural and functional organization of cells (Drubin and Nelson 1996; Gumbiner 1996). The assembly of cell-cell adhesion complexes has been studied extensively in cultured kidney cell lines (Cowin et al. 1986; Nagafuchi and Takeichi 1988; Ozawa et al. 1989; Piepenhagen and Nelson 1993; Hinck et al. 1994; Näthke et al. 1994; Marrs et al. 1995; Adams et al. 1996; Gumbiner 1996; Rajasekaran et al. 1996). Defects in the expression or interactions of catenins and E-cadherin have been postulated to contribute to tumor formation and progression (Munemitsu et al. 1995; Wakatsuki et al. 1996). Cell-extracellular matrix adhesion, especially integrin interaction with matrix proteins, regulates cell proliferation and matrix turnover during cell injury (Adler 1994). Recently, a potential role for integrins in acute renal failure has been suggested (Goligorsky et al. 1993; Simon 1994; Lieberthal et al. 1997). Detachment of viable tubule epithelial cells from the basement membrane has been documented after acute ischemic injury to the kidney (Racusen et al. 1991). Such detachment would lead to backleak of glomerular filtrate and could also be important in the production of cellular casts, as suggested in cultured kidney cell lines and rat (Gailit et al. 1993; Goligorsky and DiBona 1993).
Na+,K+-ATPase and its associated cytoskeletal proteins ankyrin and fodrin play a vital role in ion and solute reabsorption and the maintenance of electrolyte homeostasis (Almers and Stirling 1984; Fish and Molitoris 1994). The subcelluar distribution and interaction of these proteins have been studied mostly in cultured cell lines and in tissues from rodents (Katz et al. 1979; Molitoris 1991; Molitoris et al. 1992; Doctor et al. 1993). However, maldistribution of these proteins has recently been demonstrated after ischemia/reperfusion injury to human renal allografts (Alejandro et al. 1995).
The distribution of the aforementioned cell membrane-associated proteins has been described fully only in the mouse kidney (Piepenhagen and Nelson 1995; Piepenhagen et al. 1995). The purpose of the present study was to undertake a comparable analysis in the human kidney. To do this, we performed an immunohistochemical analysis of biopsy tissues of human kidney taken from living transplant donors during nephrectomy.
Materials and Methods
Subjects
The subjects of our study were nine healthy donors of a kidney for transplantation into a close relative with end-stage renal failure. Each donor and recipient of the kidney transplant gave informed consent for the performance of a renal biopsy during donor nephrectomy. The procedure was approved previously by the Committee for Research in Human Subjects at Stanford University. The donors ranged in age from 23 to 54 years. They had been diagnosed as having healthy kidneys by virtue of normal levels of creatinine clearance (109 ± 7 ml/min/1.73 m2) and protein excretion (66 ± 12 mg/24 hr). The predonation workup also included a medical history, physical examination, routine urinalysis, blood chemistry, and renal angiogram, all of which failed to reveal evidence of renal disease, diabetes, or hypertension. All recipients of these kidneys subsequently demonstrated prompt functional recovery of the allograft as judged by a rapid and progressive decline of serum creatinine to less than 1.4 mg/dl by the end of the first post-transplant week. To ensure the presence of medullary as well as cortical tissue, the biopsy was performed with a needle biopsy rather than by surgical removal of a wedge of tissue. The biopsy, using an 18-gauge biopsy needle, was performed immediately before application of the vascular clamp to the renal artery of the donor at the time of donor nephrectomy.
Immunohistochemistry
Antibodies. An affinity-purified rabbit antibody against human ZO-1 was raised against a 69-kD fusion protein corresponding to amino acids 463-1109 of ZO-1. An affinitypurified rabbit polyclonal antibody against human occludin was raised against a fusion protein consisting of the 150 amino acid C-terminal of occludin. Both antibodies were obtained from Zymed Laboratories (So. San Francisco, CA) and were used at dilutions of 1:10,000 and 1:200, respectively (see Jou et al. in press). An affinity-purified rabbit polyclonal antibody against α-catenin was raised against the 15 amino acid C-terminal of α-catenin and was kindly provided by Dr. Inke Näthke (Stanford University). It was used at a dilution of 1:200 (see Näthke et al. 1994). Rabbit polyclonal anti-human β-catenin antibody (YR7) raised against a recombinant N-terminal half of human β-catenin was a gift from Dr. David L. Rimm (Yale University) and was used at a dilution of 1:1000 (see Rimm et al. 1995). Rabbit polyclonal antibodies against the α-subunit of Na+,K+-ATPase, ankyrin-3, and the α-subunit of fodrin have been described previously (Piepenhagen et al. 1995), and were used at a dilution of 1:100. Double immunofluorescence staining with antibodies against either aquaporin 1 or 2 (Maunsbach et al. 1997), cytokeratin 8, and Tamm-Horsfall protein was used to identify proximal tubules, collecting ducts, and distal straight tubules (or thick ascending limbs of the loops of Henle) (for details see Piepenhagen and Nelson 1995; Kwon et al. 1998). The pattern of cytokeratin 8 distribution permitted proximal segments to be distinguished from distal convoluted segments, collecting ducts, and distal straight tubules. Anti-human Tamm-Horsfall protein monoclonal antibody was used to identify distal straight tubules, and was purchased from Accurate Chemical & Scientific (Westbury, NY). The antibody was used at a dilution of 1:200. (For the distributions of these marker proteins of nephron segments, see Piepenhagen and Nelson 1995: Kwon et al. 1998.)
Immunofluorescence Staining. Frozen tissue blocks were sectioned at -35 to -40C using a cryostat (2800 Frigocut N; Reichert-Jung, Nussloch, Germany). Sixm m-thick sections were transferred onto subbed glass slides coated with 0.1% gelatin and 0.01% chromium potassium sulfate. Frozen sections were allowed to warm to room temperature (RT) and were then extracted for 10 min at RT with cytoskeleton buffer [50 mM NaCl, 300 mM sucrose, 10 mM piperazine-N,N'-bis 2-ethane-sulfonic acid (pH 6.8), 3 mM MgCl, 0.5% Triton X-100, and 1 mM phenylmethylsulfonyl fluoride (PMSF)]. We have found that this brief incubation in Triton X-100 buffer increases the accessibility of antibodies into the tissue section (see Piepenhagen and Nelson in press). Slides were washed twice in PBS at RT for 10 min. Slides were incubated in blocking solution for 2 hr at RT in a humidified chamber. The blocking solution consisted of PBS with 20% normal goat serum, 0.2% bovine serum albumin, 50 mM NH4 Cl, 25 mM glycine, and 25 mM lysine. Slides were washed twice in PBS and 0.2% bovine serum albumin for 10 min at RT. They were then incubated with primary antibody overnight at 4C in a humidified chamber. Primary antibodies were diluted in PBS containing 20% normal goat serum and 0.2% bovine serum albumin. For double labeling experiments, sections were incubated with both primary antibodies. The following day, slides were again washed twice with PBS and 0.2% bovine serum albumin for 10 min at RT. Slides were incubated with the appropriate secondary antibody solutions for 2 hr at RT in a humidified chamber. For double labeling experiments, sections were co-incubated with both secondary antibodies at the same time. Rhodamine-, fluorescein-, and AMCA-conjugated secondary antibodies were diluted 1:200 in PBS containing 20% normal goat serum and 0.2% bovine serum albumin. After the secondary antibody incubation, slides were washed twice in PBS and 0.2% bovine serum albumin as above and then mounted with glass coverslips in PBS containing 16.7% Mowiol (Calbiochem; La Jolla, CA), 33% glycerol, and 0.1% paraphenylene diamine. Slides were viewed and photographed using a Zeiss Axioplan epifluorescence microscope equipped with differential interference contrast optics. Photographic slides (Kodak Ektachrome ASA 400; Eastman Kodak, Rochester, NY) were used to assess the distribution of each protein. Photographs were taken from at least three different fields for each protein at x 40, x 63, or x 100 magnification.
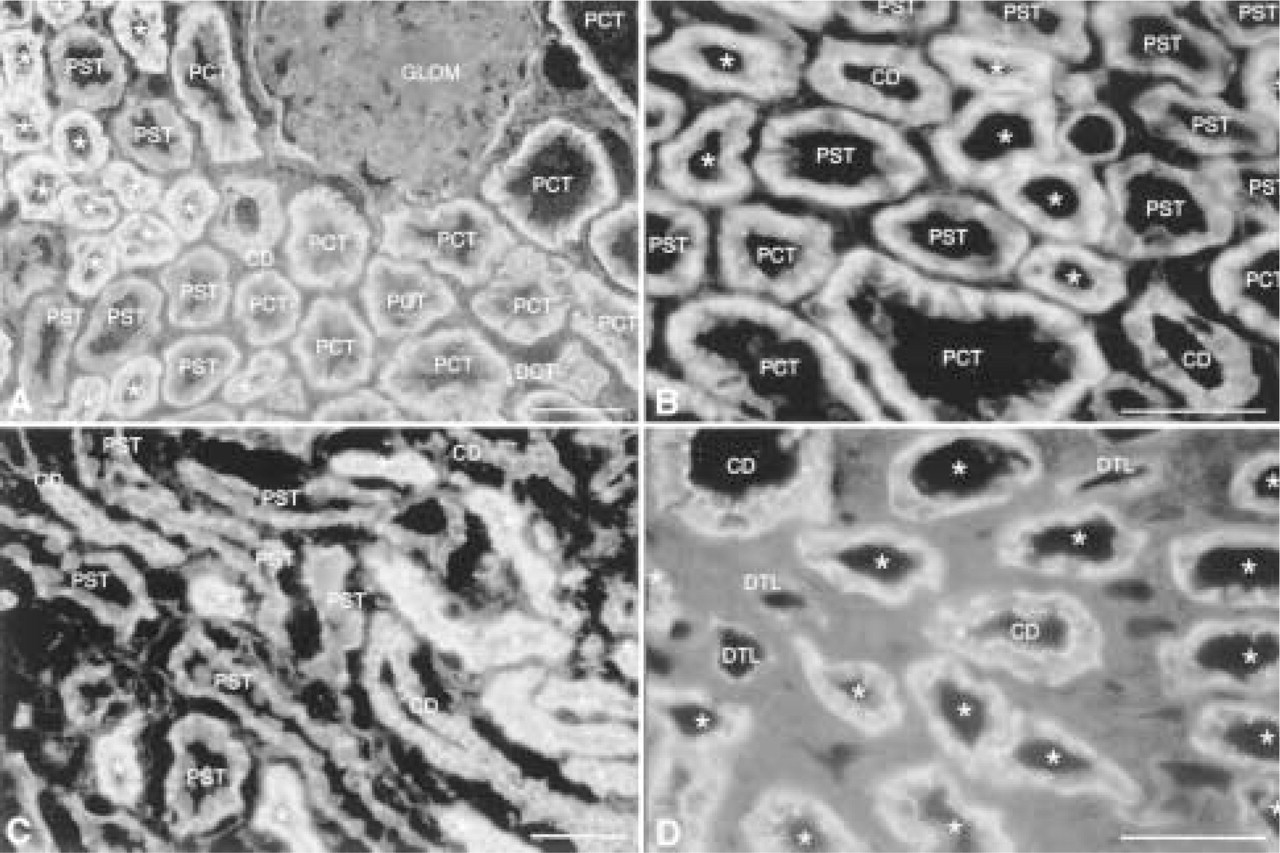
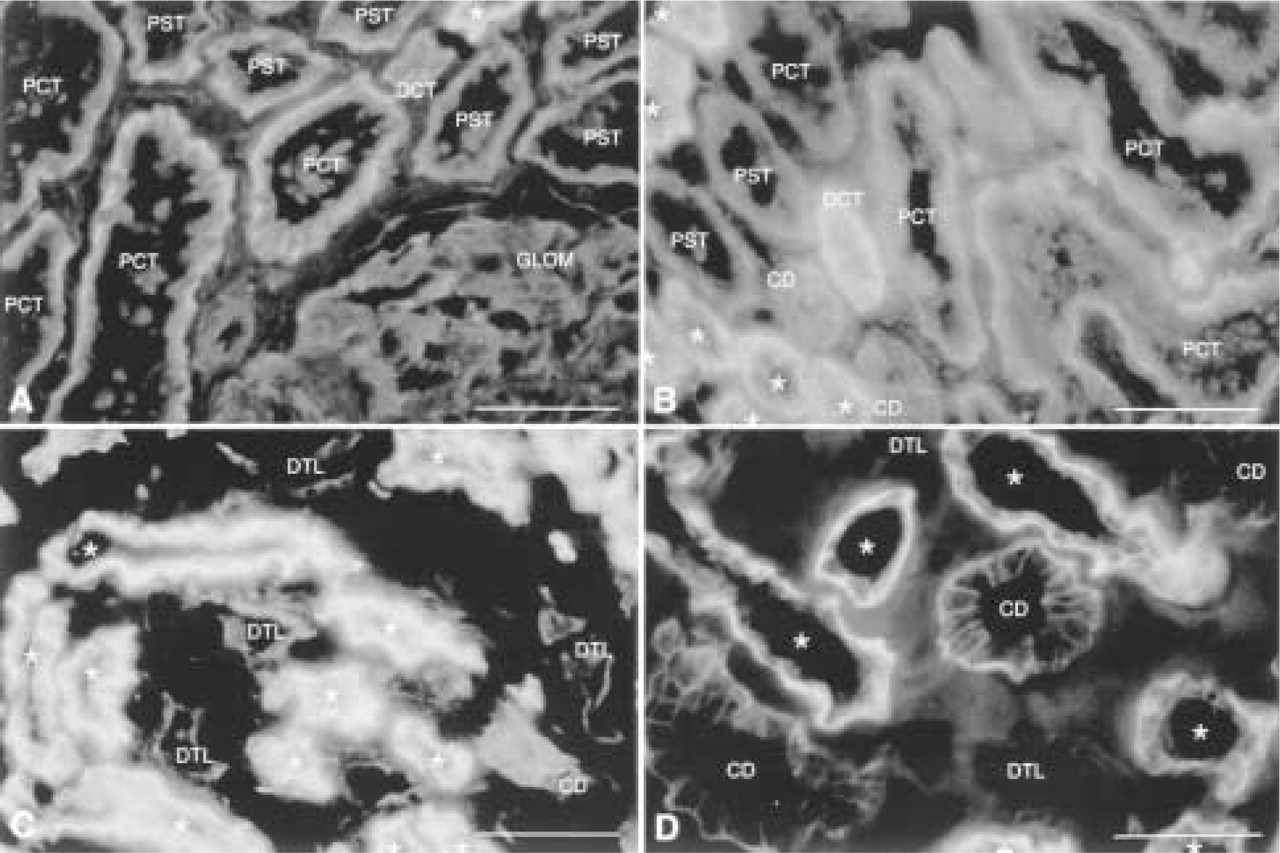
Expression of occludin in outer cortex (
Results
The needle biopsy cores contained cortex and outer medulla. Representative photomicrographs of the staining for each protein are organized in each figure (A-D) from the superficial cortex to inner stripe of outer medulla. The kidney tissue from each subject showed a similar and invariable pattern of protein distributions for each segment of the nephron.
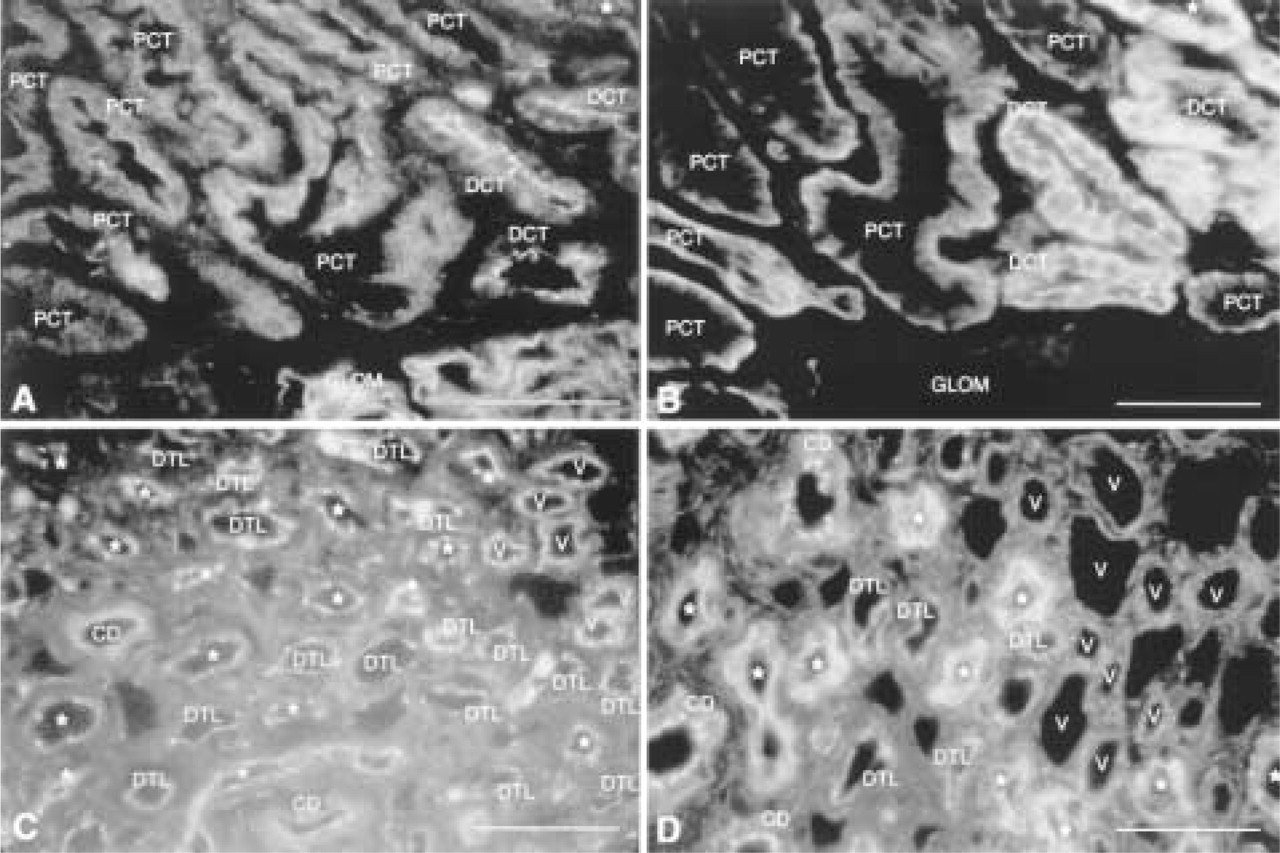
Comparison of ZO-1 and occludin distribution in sequential tissue sections from the same subject. (
Identification of Nephron Segments
We used antibodies to specific protein markers of different segments of nephrons, including CHIP-28 (AQP-1) (Denker et al. 1988; Sabolic et al. 1992; Fushimi et al. 1993; Nielson et al. 1993a,b, 1995), cytokeratin 8, Tamm-Horsfall protein, and AQP-2 (see Kwon et al. 1998; Piepenhagen et al. 1995). The results of our survey are summarized here (see also Figure 8), and the distributions of these proteins were found to be closely similar to their distributions previously reported by us (see Piepenhagen et al. 1995; Kwon et al. 1998). As we have described previously, we detected strong staining for AQP-1 along the entire proximal tubule and descending thin limb of Henle, weak staining in the mesangial region of glomeruli, and occasional staining of peritubular capillaries in the medulla. Cytokeratin 8 staining was strong in all tubule segments except for the ascending thin limb of Henle, glomeruli, and blood vessels. Tamm-Horsfall protein stained cells of distal straight tubules (thick ascending limbs of Henle) but all other nephron segments stained weakly. Staining of AQP-2 was restricted to collecting duct cells that are probably principal cells in which the ADH-sensitive water channel is known to exist. However, staining becomes less intense in the cells of medullary collecting ducts.
Distribution of Tight Junction Proteins
Occludin staining (Figures 1 and 2) is detected in parietal epithelium of Bowman's space, and cells of all tubule segments except for the ascending thin limb of Henle. Cells of proximal convoluted and straight tubules, distal straight tubule, and distal convoluted tubule show distinctly basolateral membrane distribution of the staining for occludin. In cells of the proximal straight tubule, lateral membrane staining is more pronounced: Note that distinctive punctate staining at the luminal surface, compared to that of ZO-1 (see Figure 2), is not detected. Staining in cells of the descending thin limb is apparent at intercellular junctions and basal membranes and is less intense compared with that in other tubule cells. Basolateral membrane and intercellular staining for occludin at the apical junctional complex is apparent in cells of distal straight and convoluted tubule, and intercellular staining is pronounced in cells of collecting ducts. Basolateral membrane staining in collecting duct does not exceed the level of background staining. Blood vessels are stained for occludin at intercellular junctions between endothelial cells but less intensely than that for ZO-1 (see Figure 2). Glomeruli and peritubular capillaries do not stain for occludin. ZO-1 is localized to the apical intercellular junctional complex in all renal tubular epithelial cells, although some diffuse cytoplasmic staining is also detected (Figure 2). Proximal tubule cells show a fine punctate staining along the luminal surface. Within the same section, the intensity of staining is weak in cells of the descending thin limb of Henle, stronger in distal straight tubules, and strongest in distal convoluted tubules and collecting ducts. Distal straight tubule, distal convoluted tubule, and collecting duct show strong, widely spaced punctate staining along the luminal surface, consistent with the flattened morphology of the cells. Staining in the loop of Henle is sparse. Glomeruli demonstrate strong curvilinear staining consistent with a localization of ZO-1 to the junction of podocyte foot processes and slit diaphragms.
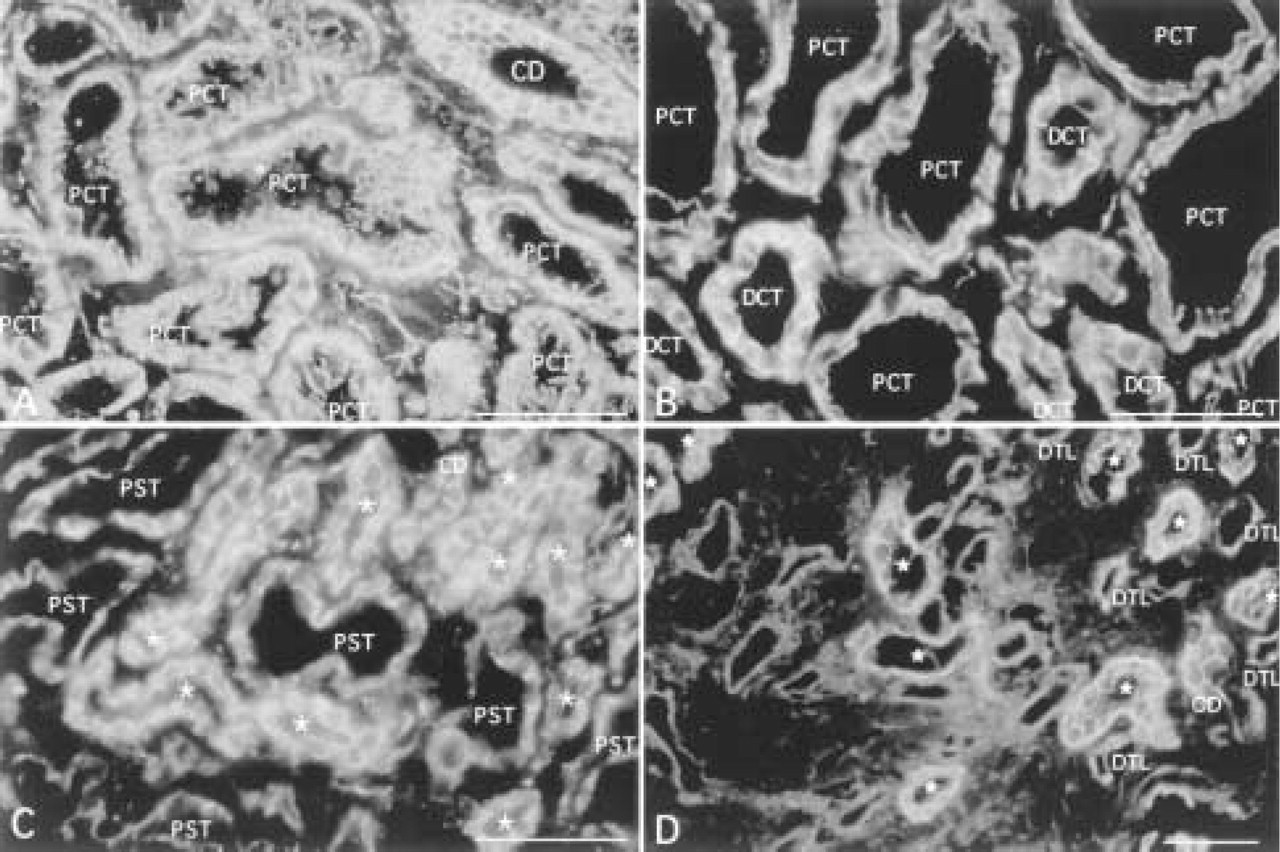
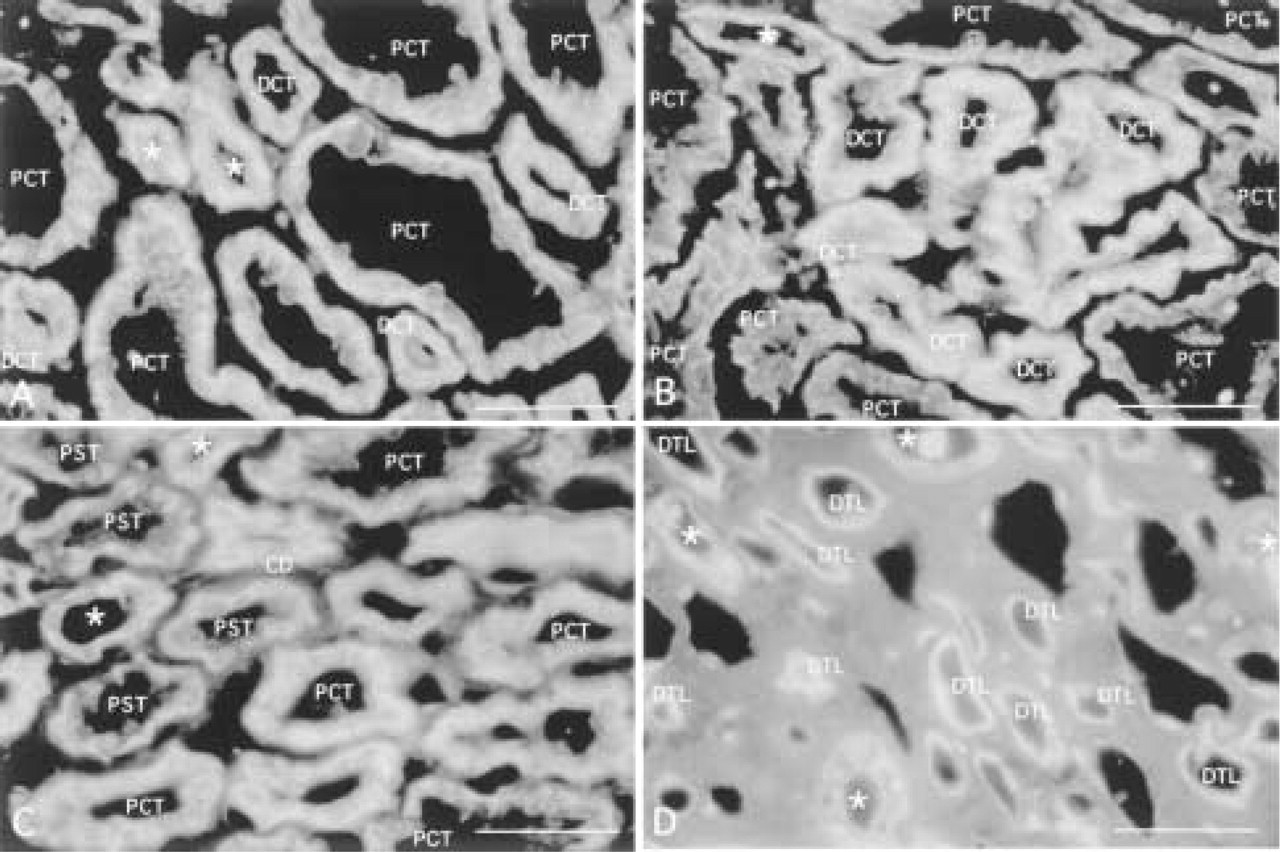
Expression of α-catenin in outer cortex (
Distribution of Cytoplasmic Proteins (Catenins) of Adherens Junctions
Cadherin cell-cell adhesion proteins are linked to the actin cytoskeleton through two cytoplasmic proteins termed β-catenin, which binds to cadherin cytoplasmic domain and α-catenin, which binds to β-catenin and the actin cytoskeleton (for review see Gumbiner 1996). Staining for α-catenin (Figure 3) and β-catenin (Figure 4) is detected in all tubule cells, endothelial cells of glomeruli, blood vessels, and peritubular capillaries. Staining for both proteins is localized to the ba-solateral membrane of cells of all tubule segments except for the ascending thin limb of Henle. The pattern of staining in proximal tubule cells frequently appears as a stringy or serpiginous pattern which corresponds to a tangential view of lateral membrane interdigitations. Lateral membrane staining is accentuated towards the luminal side at the apical junctional complex. In proximal straight tubules, descending thin limb of Henle, and distal straight tubules, staining is distributed more strongly to lateral membranes than to basal membranes, although lateral membrane staining in proximal straight tubules is more widely spaced than that in distal tubules. Cells of distal convoluted tubules and collecting ducts demonstrate strong basolateral membrane staining. Occasionally, two parallel lines of staining are observed in distal convoluted tubules and collecting ducts that correspond to lateral membranes of adjacent cells. Parallel staining of tubules for plakoglobin (γ-catenin) revealed cell type and subcellular distributions similar to those of α- and β-catenin (data not shown).
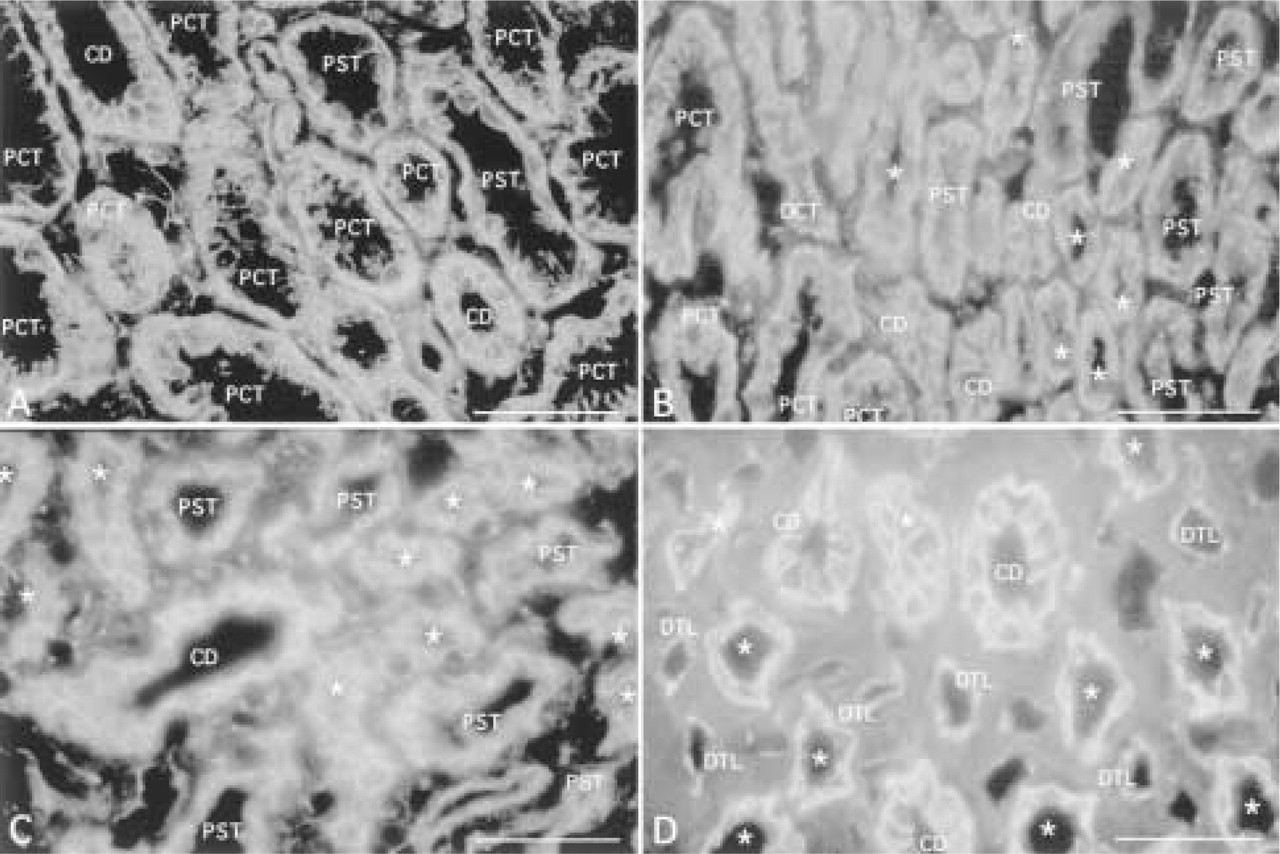
Expression of β-catenin in outer cortex (
Distribution of Na+,K+-ATPase and Associated Membrane Cytoskeletal Proteins
Previous studies have demonstrated that Na+,K+-ATPase is tightly bound to a membrane-cytoskeletal complex composed of ankyrin and the actin binding protein fodrin (spectrin). Na+,K+-ATPase (Figure 5), ankyrin (Figure 6), and fodrin (Figure 7) staining is detected in proximal convoluted, proximal straight, distal straight and distal convoluted tubules, collecting ducts, and glomeruli. The intensity of staining is variable, with the highest intensity in distal straight tubules, followed by distal convoluted tubules, then proximal convoluted tubules, proximal straight tubules, and finally collecting ducts. The intensity of the staining is weakest in cells of proximal straight tubule and collecting duct, although it becomes stronger on the basolateral membrane of collecting duct cells in inner stripes of the outer medulla. Staining is localized to the basolateral membrane with more abundance in the basal membrane in cells of the proximal convoluted and distal straight tubules. Little or no staining above background is detected in the thin loop of Henle. Staining is occasionally present in endothelial cells of glomeruli. We note that cell type and subcellular distributions of Na+,K+-ATPase, ankyrin, and fodrin were similar.
Expression of Na+,K+-ATPase in outer cortex (
Expression of ankyrin in outer cortex (
Discussion
Differential distribution of Na+,K+-ATPase, ankyrin, fodrin, E-cadherin, ZO-1, α- and β-catenin, plakoglobin, desmoplakin, laminin A, and a6-integrin along the kidney nephron has been previously described in the mouse (Piepenhagen and Nelson 1995; Piepenhagen et al. 1995). However, characterization of the distribution of these proteins has not been described in detail in normal human kidney tissues. Here we present a comprehensive analysis of these proteins to establish a foundation to identify abnormalities in human kidney (summarized in Figure 8).
Tight Junction Complex Proteins
On the basis of previous physiological measurements of tight junction function, tight junction proteins would be expected to be more strongly expressed in distal nephron, where paracellular permeability is low, compared to proximal tubule segments, where permeability is high (Claude and Goodenough 1973). Expression of ZO-1 in human kidney was detected at the site of apical intercellular junction in cells of all segments of nephron, with the highest intensity in collecting duct and distal convoluted tubule, followed by distal straight tubule, proximal tubule, and finally the thin limb of Henle (Figure 8). The distribution of ZO-1 followed the topology of cells of each segment. Therefore, ZO-1 staining in flattened cells of thin limbs appeared more widely spaced than that in columnar cells of the proximal convoluted tubule. Previous immunohistochemical analysis of ZO-1 in mouse kidney showed distinct ZO-1 expression at the apical intercelluar junction throughout the distal nephron, weak staining for ZO-1 in cells of the proximal nephron, and no staining in cells of the descending thin limb of Henle (Piepenhagen et al. 1995). It is possible that these minor differences reflect subtle differences in tight junction organization in human and mouse kidney.
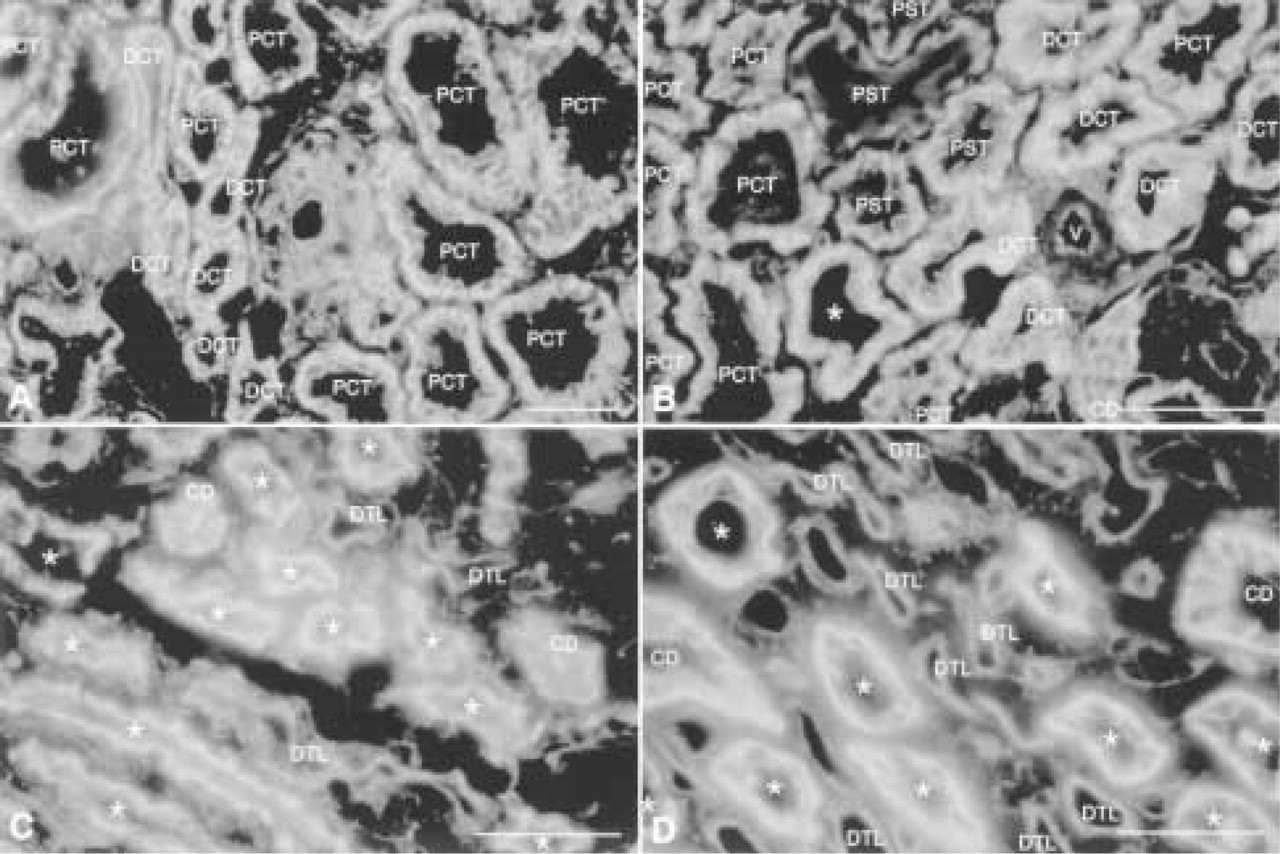
Expression of fodrin in outer cortex (
Recently, occludin has been identified as an integral membrane protein of the tight junctions (Anderson et al. 1993; Furuse et al. 1993,1994; Tsukita et al. 1996; Wong and Gumbiner 1997). Characterization of the distribution of this protein in kidney has not been reported. To our surprise, we observed a distinctive basolateral distribution in proximal tubule cells; apical junctional complex staining was distinct in cells of the distal nephron. Although ZO-1 and occludin form a complex and co-localize to the apical junctional complex in most cells (Furuse et al. 1994), the distribution of these proteins is clearly different in proximal tubule cells. Although we do not know why ZO-1 and occludin had different subcellular distributions in proximal tubule cells, it is interesting that these cells have high paracellular permeability. It is possible that the diffuse localization of occludin along the lateral membrane and not specifically to the apical junctional complex could contribute to the high permeability of proximal tubule cells.
Cell-Cell Adhesion Proteins
Immunohistochemical staining of mouse kidney showed that α- and β-catenin are distributed to the basal membrane and apical junctional complex in proximal tubule cells, and to the basolateral membrane with additional cytoplasmic staining in cells of the distal nephron (Piepenhagen and Nelson 1995). In general, the distributions of α- and β-catenin in human kidney were similar to those in mouse (see Figure 8). However, cells of the human nephron exhibited more distinct lateral staining for α- and β-catenin throughout the nephron. A stringy or serpiginous pattern of staining, corresponding to convoluted lateral membrane interdigitation of proximal tubule cells, was frequently noted in human kidney.

Summary of distribution of marker proteins, AQP 1 (CHIP 28), cytokeratin K8, Tamm-Horsfall protein (THP), and AQP 2; tight junction complex protein, ZO-1 and occludin; cell- cell adhesion protein, α-, β-catenin, and plakoglobin; cell-substrate adhesion protein, α6-integrin, laminin; and Na+,K+-ATPase and membrane-associated cytoskeleton proteins, ankyrin and fodrin. The distributions of α6-integrin and laminin have been reported by us previously (Kwon et al. 1998) and were reproduced in the present study (data not shown). Abbreviations as in Figure 1, except that DST(TAL) is substituted for ∗ and ATL indicates the ascending thin limb of loop of Henle. The thickness of bars indicates relative intensity of staining of a protein in a given nephron segment based on analyses of staining in the same section.
Na+,K+-ATPase, Ankyrin, and Fodrin
The cellular distribution and staining intensity of Na+,K+-ATPase in mouse kidney correlated well with the description of Na+,K+-ATPase enzymatic activity along the nephron (Katz et al. 1979; Garg et al. 1981). Our analysis of Na+,K+-ATPase distribution in human kidney (see Figure 8) is compatible with these findings in mouse kidney, with the following exceptions. Note that the polyclonal antiserum specific for the α1-subunit of the Na+,K+-ATPase, and the fixation/ staining protocol used to detect the distribution of Na+,K+-ATPase in human kidney were the same as those used in the mouse kidney (Piepenhagen et al. 1995). First, the relative intensity of Na+,K+-ATPase staining in cells of the proximal convoluted tubule was weaker than that in mouse, in which proximal tubule staining is comparable to that in other nephron segments. Second, in distal convoluted tubule adjacent to distal straight tubule, the intensity of Na+,K+-ATPase staining was similar to that of the distal straight tubule. However, cells in the rest of the distal convoluted tubule showed weaker staining intensity than that in distal straight tubule. Third, cells of the collecting duct in the inner stripe of the outer medulla were stained stronger for Na+,K+-ATPase than those of the cortex and outer stripe of the outer medulla. Fourth, the distribution of Na+,K+-ATPase was more basal and cytoplasmic in cells of the proximal tubule and distal straight tubule, whereas the cells of other tubules showed a basolateral membrane distribution similar to that in mouse kidney. The highest intensity of Na+,K+-ATPase staining was in the distal straight tubule and the adjacent part of the distal convoluted tubule. The lowest intensity was in the proximal straight tubule and collecting duct, and little or no staining was detected in thin loops of Henle. Differences in the staining intensity of Na+,K+-ATPase along the nephron correlate well with Na+,K+-ATPase activity along the nephron (Katz et al. 1979; Garg et al. 1981). The presence of some cytoplasmic staining in cells of the proximal and distal straight tubule might suggest mild injury to this ischemia-sensitive region of the kidney owing to vasoconstrictor influences during surgical manipulation.
Previous studies have demonstrated that Na+,K+-ATPase, ankyrin, and fodrin bind with high affinity to form a complex, and it has been proposed that the interaction between Na+,K+-ATPase and the membrane-associated cytoskeleton is important for restricting the distribution of Na+,K+-ATPase to a specific domain of polarized epithelial cells (Bennett 1990; Nelson 1992; Nelson et al. 1990; Doctor et al. 1993; Leiser and Molitoris 1993). In human kidney, the intensity of the staining of ankyrin and fodrin along the nephron was less variable, especially of fodrin, than that of Na+,K+-ATPase, although the general trend of staining in different nephron segments for all three proteins was similar (Figure 8). The subcellular distributions of ankyrin and especially fodrin were distinctly more restricted to the basolateral membrane domain than that of Na+,K+-ATPase.
There are several examples of studies indicating abnormal distribution of proteins in human renal diseases (eg. Ivanyi et al. 1993; Mount et al. 1993). For example, an abnormal distribution of Na+,K+-ATPase and distribution of different aquaporins in cyst epithelium of hereditary polycystic kidney disease have been described (Wilson et al. 1991; Bachinsky et al. 1995). In addition, renal ischemia-induced degradation of ankyrin and fodrin with altered surface membrane polarity, loss of E-cadherin correlated with invasiveness of transitional cell cancers, and a defect in the regulation of β-catenin levels in carcinomas have been described (Bennett 1990; Nelson et al. 1990; Nelson 1992; Doctor et al. 1993; Leiser and Molitoris 1993; Munemitsu et al. 1995; Wakatsuki et al. 1996). Finally, the importance of laminin-a6-integrin interaction for epithelial polarization in developing tubules and a contributory role of redistribution of α3- and β1-integrin to the apical membrane in the pathophysiology of ischemic injury to the kidney has been proposed (Klein et al. 1988; Sorokin et al. 1990; Gailit et al. 1993; Lieberthal et al. 1997). Our detailed analysis of segmental and subcellular distributions of classes of proteins along the nephron provides a strong foundation for the analysis of abnormalities in protein distribution induced by structural damage to the kidney and the ensuing effects on organ function.
Footnotes
Acknowledgments
Supported by grants R01 DK50712 and M01-RR00070 from the National Institutes of Health. Dr Kwon's postdoctoral fellowship was supported by the Satellite Dialysis Centers Fund in Nephrology.