
Abstract
Ultrastructural localization of a P29 protein of Toxoplasma gondii was examined on thin sections by an immunogold technique using a P29 antigen-specific monoclonal antibody (5-241-178). Immunolocalization of the P29 protein in extracellular tachyzoites demonstrated that this antigen was present in the dense granules. Thus, we have identified this P29 antigen as the seventh protein (GRA7) to be localized to the dense granules of T. gondii. P29 immunolocalization in intracellular tachyzoites demonstrated association of this antigen with the parasite membrane complex, tubular elements of the intravacuolar network, and with the parasitophorous vacuolar membrane. Our immunolabeling data suggest trafficking of the P29 (GRA7) antigen from the dense granule via the intravacuolar network to the parasitophorous vacuolar membrane on invasion of the tachyzoite into the host cell.
Toxoplasma gondii, a coccidian protozoan, is an obligate intracellular parasite of vertebrates and an important opportunistic pathogen in humans. T. gondii has a powerful capacity to infect a wide range of cell types, both phagocytic and nonphagocytic cells. Host cell invasion is an active process that leads to parasite internalization in a new membrane-bounded intracellular compartment termed the parasitophorous vacuole (PV). This specialized vacuole prevents acidification (Sibley et al. 1985) and fusion with other endocytic vesicles, including both early and late endosomes and lysosomes (Joiner et al. 1990; Sibley 1993). In addition, the parasite forms a network of membranous tubules within the lumen of the vacuole and connecting with the vacuolar membrane.
T. gondii has two main secretory organelles that are involved in the invasion process, the rhoptries and the dense granules (Cesbron-Delauw et al. 1989; Charif et al. 1990; Achbarou et al. 1991; Leriche and Dubremetz 1990,1991; Saffer et al. 1992; Dubremetz et al. 1993). In the early stages of invasion of the host cell by Toxoplasma, the apically located rhoptries discharge components that perturb the host cell membrane and therefore facilitate the internalization of the parasite. Subsequently, the dense granules release their contents into the vacuolar space after invasion is complete. After exocytosis, the proteins GRA1, GRA2, GRA4, and GRA6 localize to the intravacuolar network, whereas GRA3 and GRA5 localize primarily to the parasitophorous vacuole membrane (PVM) (Cesbron- De lauw 1994). The trafficking of the dense granule proteins to the PV and PVM after invasion presumably plays an important role for multiplication of the parasite within the host cell and evasion of the immune response.
Dense granule proteins of T. gondii have been described as highly immunogenic molecules that may be involved in humoral immunity (Darcy et al. 1988; Decoster et al. 1988; Mevelec et al. 1992). In this work we describe the cloning of a new gene encoding a novel Toxoplasma antigen. This gene was cloned by screening a phage library with pooled plasma obtained from individuals with acute toxoplasmosis. Subsequent isolation of a mouse monoclonal antibody directed against this new antigen permitted the identification of the product of this new gene as the P29 antigen. We report the quantitative immunolocalization of the P29 antigen to the dense granules of T. gondii and propose the name GRA7 according to the uniform T. gondii nomenclature (Sibley et al. 1991).
Materials and Methods
Parasite Culture
Tachyzoites of T. gondii RH strain were maintained by twice-weekly passages of peritoneal fluid into female Swiss mice carrying TG sarcoma cells. Tachyzoites were recovered from the peritoneal cavity 3 days after infection, washed with PBS, pH 7.2, collected by centrifugation, and resuspended (1200 x g, 4 min) at the desired cell density. The tachyzoite population was on the order of 109/ml.
Isolation of T. gondii DNA, RNA, and Protein, and Synthesis of cDNA
A 10-liter secondary suspension culture of HeLa cells infected with the RH strain of T. gondii was grown to a tachyzoite density of approximately 1 x 107/ml and filtered through a 10-μm Millipore (Bedford, MA) Polygard cartridge filter to remove HeLa cells from the tachyzoites. The tachyzoite filtrate obtained contained less than 1% HeLa cells. The tachyzoites were then concentrated by centrifugation, washed, and resuspended in Hank's buffer. The tachyzoite concentrate was then pipetted dropwise into liquid nitrogen and the frozen tachyzoite pellets were recovered and stored at -80C until further use. The tachyzoite pellets were converted to tachyzoite powder by grinding the pellets to a fine powder using a mortar and pestle chilled with dry ice and liquid nitrogen. The tachyzoite powder was subsequently used for the isolation of tachyzoite nucleic acid as described below.
T. gondii DNA was isolated from the tachyzoite powder using the Stratagene DNA extraction kit (La Jolla, CA). The tachyzoite powder was dissolved in Solution 2 and total DNA was isolated according to the kit's protocol. After ethanol precipitation and resuspension of the DNA in TE buffer, undissolved DNA and contaminating polysaccharides were removed by centrifugation at 200,000 x g for 1 hr.
T. gondii RNA was isolated from the tachyzoite powder using the Stratagene RNA isolation kit. The tachyzoite powder was dissolved in Solution D and total RNA was isolated according to the kit's protocol. After ethanol precipitation and resuspension of the RNA in diethyl pyrocarbonate (DEPC)-treated water, polyA+ RNA was selected with an oligo-dT column using a Pharmacia mRNA isolation kit (Piscataway, NJ). The purified mRNA was concentrated by ethanol precipitation and stored in DEPC-water at -80C until further use.
T. gondii protein was isolated from filtered and concentrated tachyzoites. Approximately 2.4 x 108 tachyzoites were dissolved in 1 ml sample buffer (1% glycerol, 1.5% SDS, 125 mM Tris-HCl, pH 6.8, 0.05% bromophenol blue) and boiled for 5 min. The final protein concentration was approximately 1 mg/ml. The protein preparation was stored at -20C until further use.
Purified T. gondii mRNA was used as a template for the synthesis of cDNA using the ZAP-cDNA Synthesis kit (Stratagene). The cDNA was then ethanol-precipitated, resuspended in water, and stored at -20C until further use as a template for construction of a Toxoplasma cDNA library.
Cloning and Expression of the Gene Encoding the Novel Antigen
Standard methods were used for isolation and analysis of plasmid DNA, DNA sequence analysis, and Southern analysis (Sambrook et al. 1989). DNA and amino acid sequences were analyzed using the Geneworks software package (Intelligenetics; Mountain View, CA). A T. gondii cDNA library with 660,000 members was constructed in the UNIZAP XR vector using T. gondii cDNA and the ZAP-cDNA Gigapack II Gold cloning Kit (Stratagene). The cDNA library was screened with pooled plasma from patients with acute toxoplasmosis (Antibody Systems; Bedford, TX) using Duralose UV membranes and the picoBLUE Immunoscreening Kit (Stratagene) and a goat anti-human IgM horseradish peroxidase-labeled conjugate (Kirkegaard & Perry Laboratories; Gaithersburg, MD). Approximately 130,000 phage plaques were screened resulting in the isolation of four positive clones. These four phage clones were converted to plasmid clones (pGM610, pGM611, pGM612, and pGM613) using the subcloning cloning protocol from the ZAP-cDNA Synthesis Kit, and were found by restriction enzyme analysis to contain DNA inserts in the range from 0.7 to 1.3
The 1.3-
Cloning and Expression of Other Toxoplasma Antigens
Using the published DNA sequences, PCR primers were generated to clone the genes encoding the following Toxoplasma antigens previously described in the literature into the CKS expression vector (Bolling and Mandecki 1990) by PCR amplification: P22 (SAG2) (Prince et al. 1990); P24 (GRA1) (Cesbron-Delauw et al. 1989); P25 (Johnson and Illana 1991); P28 (GRA2) (Prince et al. 1989); P30 (SAG1) (Burg et al. 1988); P35 (Knapp et al. 1989); P41 (GRA4) (Mevelec et al. 1992); P54 (ROP2) (Saavedra et al. 1991); P66 (ROP1) (Knapp et al. 1989; Ossorio et al. 1992); and P68 (Knapp et al. 1989). The cloned DNA sequences were confirmed by DNA sequence analysis. Expression of these Toxoplasma recombinant proteins in E. coli was performed as described for the CKS-novel antigen fusion protein.
Generation of the 5-241-178 Mouse Monoclonal Antibody Directed Against the Novel Antigen
Swiss mice (Charles River Laboratories; Wilmington, MA) were infected intraperitonally with 2.5 x 107 tachyzoites of T. gondii strain TS4 (American Type Tissue Collection; Rockville, MD). Five days later the mice were treated orally with 10 mg pyrimethamine and 200 mg sulfamethoxazole per kg daily for 10 days. After 12 additional weeks, the mice were injected i.v. with 1.2 x 107 sonicated tachyzoites 3 days before fusion. Resulting hybrids from the PEG-mediated fusion of splenocytes and the SP2/0 myeloma cells were screened with sonicated tachyzoites, sonicated E. coli lysates containing the CKS-novel antigen, CKS-P22, P24, P25, P28, P30, P35, P41, P54, P66, and P68 CKS fusion proteins and unfused CKS using standard procedures (Köhler and Milstein 1975, 1976; Goding 1986). Putative clones were cloned out by limiting dilution and retested. One highly reactive monoclonal antibody was isolated which was designated 5-241-178, which reacted very strongly with sonicated tachyzoites and with the CKS-novel antigen but showed no reactivity to the other Toxoplasma recombinant fusion proteins or to unfused CKS. This monoclonal antibody clone was found to produce IgG type antibodies as determined using a mouse monoclonal antibody isotyping kit (Sigma Chemical; St Louis, MO).
Gel Electrophoresis and Western Blot Analysis
Approximately 10 mg of Toxoplasma lysate and 10 mg E. coli extract containing the CKS-novel antigen fusion protein were loaded on a 4-20% gradient SDS-polyacrylamide gel in the presence of β-mercaptoethanol using a Daiichi precast gel (Integrated Separation Systems; Natick, MA). Prestained protein molecular weight markers (106-18.5 kD) were included on the gel for molecular weight determination (Bio-Rad; Richmond, CA). After electrophoresis, the proteins were transferred to nitrocellulose (Schleicher & Schuell; Keene, NH) using the ISS SemiDry Electroblotter and reagents (Integrated Separation Systems). After transfer, the membrane was saturated overnight with membrane blocking solution [1% bovine serum albumin (BSA), 1% casein acid hydrolysate, 0.05% Tween-20 in Tris-buffered saline (TBS; 20 mM Tris, 0.5 M NaCl, pH 7.5)] at room temperature (RT). The next day the membrane was briefly rinsed with water and immersed in monoclonal antibody diluent (100 mM Tris, 135 mM NaCl, 10 mM EDTA, 0.2% Tween-20, 0.01% thimerosal, 4% bovine calf serum, pH 7.5). The 5-241-178 monoclonal antibody was diluted into monoclonal antibody diluent, added to the membrane, and then incubated for 1 hr at RT. The membrane was washed once with water and twice with TBS and then immersed in conjugate diluent (100 mM Tris, 135 mM NaCl, 0.01% thimerosal, 10% bovine calf serum, pH 7.5). The goat anti-mouse IgG- horseradish peroxidase conjugate (Kirkegaard & Perry Laboratories) was diluted into conjugate diluent (1:1,000), added to the membrane, and then incubated for 1 hr at RT. The membrane was washed with water and twice with TBS-Tween-20 and then developed using BioRad HRP Color Development Reagent until the desired band intensity was achieved. Color development was stopped by rinsing the membrane with water.
Immunocytochemistry
After two washes with PBS, pH 7.2, freshly isolated tachyzoites were fixed for 2 hr with 2% paraformaldehyde, 0.5% glutaraldehyde in PBS (pH 7.2) at 4C. Tachyzoites were washed with PBS, then pre-embedded in BSA (diluted 1:4) and glutaraldehyde (25%) in the ratio 4:1, dehydrated with increasing concentrations of ethanol, and embedded in LR White resin. Ultrathin sections (thickness 80 nm) placed on gold grids were immunolabeled as follows. They were incubated first with PBS-1% BSA, pH 7.2, for 30 min at RT and then with the 5-241-178 monoclonal antibody, diluted 1:10,000 in PBS-1% BSA for 1 hr at RT. After three washes with PBS-1% BSA, thin sections were incubated for 1 hr with goat anti-mouse IgG-gold (10 nm) (Biocell; Tebu, France) diluted 1:50 for 1 hr at RT. After two washes in PBS and distilled water, the sections were contrasted with uranyl acetate. Controls without monoclonal antibody were prepared and observed.
Quantitative Immunolabeling Analysis
Conditions for the preparation of samples were designed at the same time to preserve the ultrastructure, to identify labeled organelles, and to retain immunoreactivity. The monoclonal antibody was tested at increasing concentrations up to optimal immunolabeling response, i.e., when the ratio of total signal to nonspecific signal was the highest possible. Under these conditions, we could carry out immunolabeling assays quantitatively, the antibody being bound to almost all accessible epitopes of the antigen. Thus, we could compare the distribution of the antigens quantitatively by evaluating the immunolabeling density (number of gold particles/μm2).
Quantitation was performed with an image analyzer (Bio 500; Biocom, Les Ulis, France) using transmission electron micrographs of ultrathin sections and a semiautomatic quantitative method (Bonnet et al. 1990; Boulanger et al. 1991). After digitizing of images, analysis of labeling densities was performed in three steps. First was identification and selection of the biological compartments: membrane complex, rhoptries, dense granules, reticular network, and membrane of the parasitophorous vacuole. An image segmentation was applied and areas were computed. Tachyzoite membrane complex and parasitophorous vacuole membrane were materialized by two outlines which delimited these two compartments in the section. Therefore, the gold particle distribution was estimated in the area included between these outlines (thickness about 60 nm, which represents the membrane section). Second, extraction of dense markers (gold particles) was performed automatically by mathematical morphology (Serra 1982). Third, quantitation of markers was expressed as number of gold particles/μm2 area. As a reference, we used isolated markers and determined the mean area of one marker. In the case of coalescent particles, the count was estimated from the ratio of their area to the area occupied by the reference marker. Gold particles of uniform size (10 nm) were used and the binding capacity of each particle was assumed to be the same (Bendayan 1984). Because immunolabeling was restricted to the surface of the sections, only those binding sites exposed interact with the specific antibodies. Those sites can be revealed and permit quantitative and comparative evaluations (Bendayan et al. 1987).
The average number of gold particles/μm2 was calculated by subtracting the background determined in the resin (LR White) from the labeling density of the experimental specimens. Quantitation was determined on 13 negatives for extracellular tachyzoites and 20 negatives for intracellular tachyzoites on 50 observed negatives. Only those having a good contrast were taken into account.
State 1 and State 2 of the intracellular state of the tachyzoites are discriminated from the parasitophorous vacuole membrane (PVM) labeling, PVM being the second target after the reticular network of the dense granule exocytosis.
The quantitative evaluation of the immunolabeling is expressed by averages with the standard deviation (SD). To define the significant difference in the immunolabeling among the different states (extracellular, State 1, and State 2), among five different compartments (DG, dense granules; R, rhoptries; MC, membrane complex; RN, reticular network; PVM, parasitophorous vacuole membrane), and among the different compartments during the different states, we used the U-test of Mann and Whitney (1947) and the p value was considered significant when p<0.05.
Results
Identification of the Novel Antigen as the P29 Antigen of T. gondii
The 5-241-178 monoclonal antibody was used to probe a Western blot containing total Toxoplasma protein and the recombinant CKS-novel antigen fusion protein, which is shown in Figure 1. The 5-241-178 monoclonal antibody reacted specifically with a 29-kD protein present in Toxoplasma lysate (Figure 1, Lane 2), thus identifying the novel antigen as the P29 antigen of T. gondii. The monoclonal antibody also reacted with the recombinant fusion protein (Figure 1, Lane 3) which was used to isolate the monoclonal antibody during hybridoma screening. The 5-241-178 monoclonal antibody did not react with CKS recombinant proteins containing the SAG1, SAG2, GRA1, GRA2, GRA4, ROP1, or ROP2 proteins (data not shown). The possibility that this monoclonal antibody reacts with the one of the other three dense granule proteins not tested (GRA 3, GRA5, and GRA6) is extremely remote, for the following two reasons. First, the 5-241-178 monoclonal antibody reacts with a single protein of 29 kD. The dense granule proteins GRA3 (30 kD), GRA5 (21 kD), and GRA6 (32 kD) (Cesbron-Delauw 1994) are of different molecular weights, which could be resolved by the gradient gel used to generate this Western blot. Second, there is no amino acid sequence homology between the P29 antigen and other dense granule proteins of T. gondii.

Detection of the novel antigen by Western blot using the 5-241-178 mouse monoclonal antibody. Lane 1, Prestained protein molecular weight markers, kD indicated. Lane 2, Approximately 10 mg of T. gondii lysate prepared from the RH strain; Lane 3, Approximately 10 mg of the CKS-novel antigen lysate prepared from E. coli.
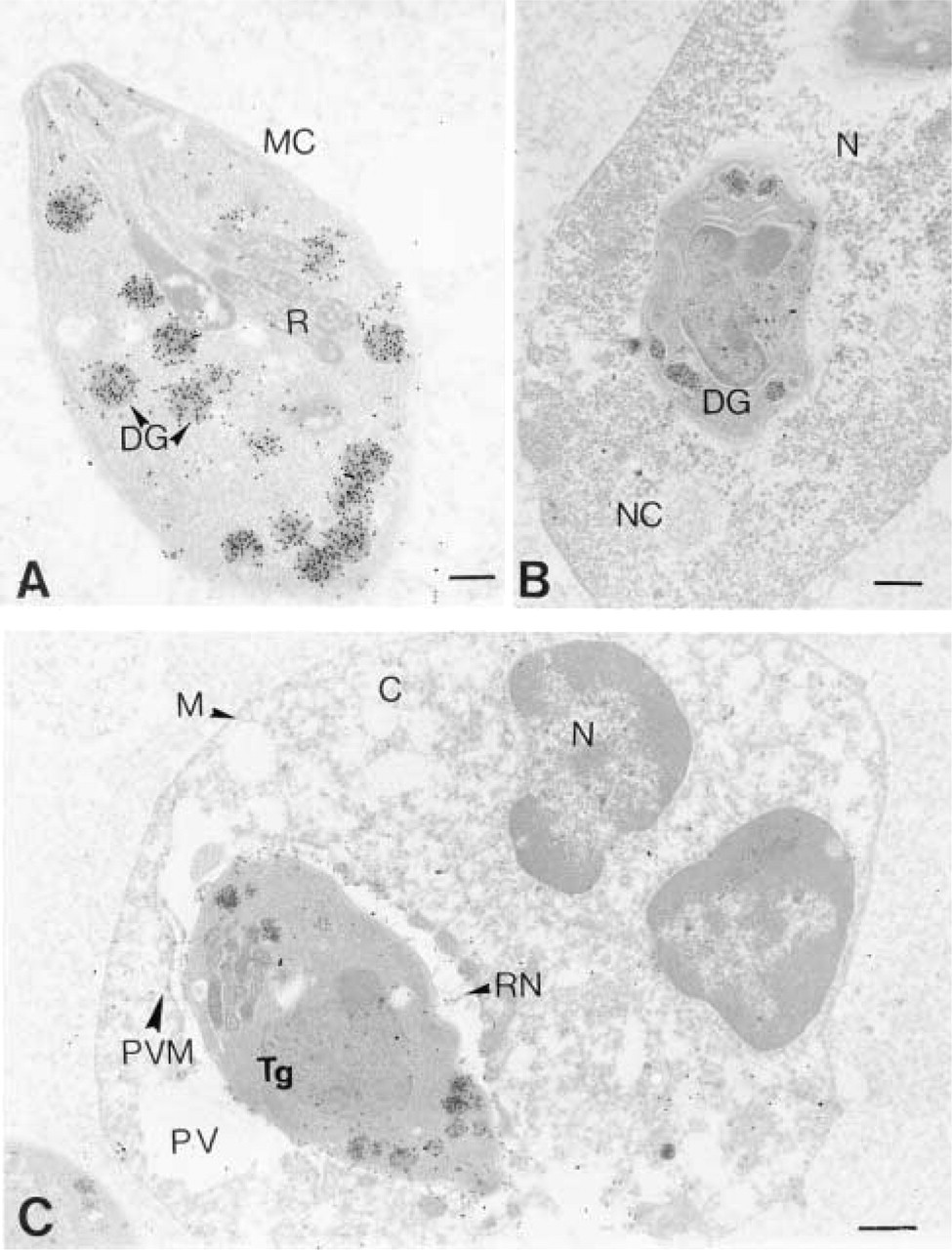
(A) Immuno-EM localization of P29 (GRA7) in extracellular T. gondii. MC, membrane complex; DG, dense granules; R, rhoptries. Bar = 0.2μm. (B) Intranuclear tachyzoite inside a necrosed cell. Immunolabeling is located in dense granules (
The distribution of the labeling density of the P29 antigen using the monoclonal antibody 5-241-178 examined in the five compartments-dense granules, membrane complex, rhoptries, reticular network of the parasitophorous vacuole, and the parasitophorous vacuole membrane-is shown in Figures 2-4. The results of the quantitative estimation of the immunolabeling are given in Table 1.
Immunolocalization of the P29 antigen in extracellular tachyzoites demonstrated that the P29 antigen was present inside the dense granules. The P29 antigen was found to be about 25 times more abundant in the dense granules than in either the rhoptries or membrane complex. With the nonparametric U-test of Mann and Whitney (1947), we observed that the immunolabeling differences between DG and MC and the DG and R were statistically significant (p<0.001). Careful examination of the nonlabeled areas in the cell and the LR White resin confirms the extremely low level of nonspecific background labeling when the monoclonal antibody was omitted from the labeling protocol [for extracellular parasites (1.5 ± 1.6) and for intracellular parasites (2.2 ± 2.4)].
Immunolocalization of the P29 antigen in intracellular tachyzoites as shown in Figures 3 and 4 and in Table 1 demonstrated that there was a differential distribution of the P29 antigen and, from all the negatives observed, we divided the intracellular state of the parasite into two states. In State 1, the P29 antigen present in the tachyzoite dense granules (DG), as in the extracellular state, was also redistributed principally in the membrane complex (MC) of the tachyzoite and in the reticular network (RN) inside the parasitophorous vacuole. In State 2, the P29 antigen distribution was extended and was found in association with the parasitophorous vacuole membrane (PVM). The quantitative P29 immunolabeling repartition is shown on Table 1. The dense granule labeling differences were not significant between the extracellular state and State 1 but were very significant between the extracellular state and State 2 (p<0.01) (the immunolabeling decreases from 1092 ± 280 to 494 ± 290) and were also significant between State 1 and State 2. The immunolabeling of the membrane complex increases from the extracellular state to State 1 and the observed labeling difference was significant (p<0.01). The immunolabeling of the PVM from State 1 to State 2 significantly increases from 23.8 ± 19.2 to 81.5 ± 47.5 (p<0.01), suggesting an inverse immunolabeling relationship between the DG and PVM from State 1 to State 2. There was probably a flux of P29 antigen secretion via the membrane complex from dense granules to the PVM once tachyzoites were inside the host cell. We observed a decrease in the immunolabeling of the membrane complex from State 1 to State 2 but the labeling difference was not significant. There were no significant immunolabeling differences in the RN between States 1 and 2. Outward extension of immunolabeling was observed from the parasitophorous vacuole into the host cell cytoplasm and plasma membrane (Figure 4). In necrosed cells (nucleus corpus observed was typical of dying cells), tachyzoites were seen inside the nucleus. Immunolabeling was always present in the dense granules and also in the nucleus (Figure 2B).
Discussion
Dense granule organelles of protozoan parasites Sarcocystis (Entzeroth et al. 1986), Plasmodium (Trager et al. 1992), and Toxoplasma (Leriche and Dubremetz 1990; Cesbron-Delauw 1994) are involved in the installation process within the host cell, presumably by contributing to the structural modifications of the parasitophorous vacuole. As the parasites grow and replicate within the cell, the parasitophorous vacuole is modified (Achbarou et al. 1991). At early times after invasion, the PVM is depleted of intramembranous particles (IPMs) relative to the host cell plasma membrane. IPMs progressively appear in the PVM as the vacuole matures (Porchet-Henneré and Torpier 1983). A tubuloreticular network appears within the vacuolar space (Sibley et al. 1986), progressively increasing as the vacuole enlarges. Although the intravacuolar network is clearly of parasite origin, the source of the IMPs in the PVM is not clear.
Dense granule secretion has all the hallmark features of regulated exocytosis, including storage of secretory proteins in specialized organelles, fusion with the plasma membrane, and release of their contents. This burst of exocytosis occurs soon after internalization. Dense granule antigens, termed GRA proteins, are differentially targeted to either the vacuolar space, the network, or the surrounding membrane of the parasitophorous vacuole.
Sequencing of six genes encoding dense granule proteins has shown typical hydrophobic signal sequences that target them in the secretory pathway. Except for the canonical signal peptide at the N-terminus, their structural features are different and may be related to their differential distribution in the parasit-ophorous vacuole (Cesbron-Delauw 1994). GRA1 (23 kD) is a calcium binding protein uniformly distributed throughout the lumen of the vacuole (Cesbron-Delauw et al. 1989) and is associated with the network through peripheral interactions (Sibley et al. 1995). GRA2 (28 kD), GRA4 (40 kD), and GRA6 (32 kD) are tightly associated with the tubular elements of the membranous network (Charif et al. 1990; Achbarou et al. 1991). Ultrastructural studies suggest that the intravacuolar network and the membrane complex of the parasite are contiguous in localized regions. The molecular structure of GRA4 suggests its association with the intravacuolar network membrane via a transmembrane region (Mevelec et al. 1992). GRA2 may be linked to lipids with two amphipathic helices present in the molecule (Mercier et al. 1993). GRA6 may be a transmembrane fibrous protein implicated in the structures of the membranous network (Lecordier et al. 1995). GRA3 (30 kD) and GRA5 (21 kD) are distributed on the membrane delimiting the PV (Achbarou et al. 1991; Dubremetz et al. 1993; Lecordier et al. 1993), and GRA3 is found both in the network and the PVM locations (Cesbron-Delauw 1994). In a similar manner to GRA5, oligomers of GRA3 associate stably with the vacuole membrane via hydrophobic interactions (Ossorio et al. 1994).
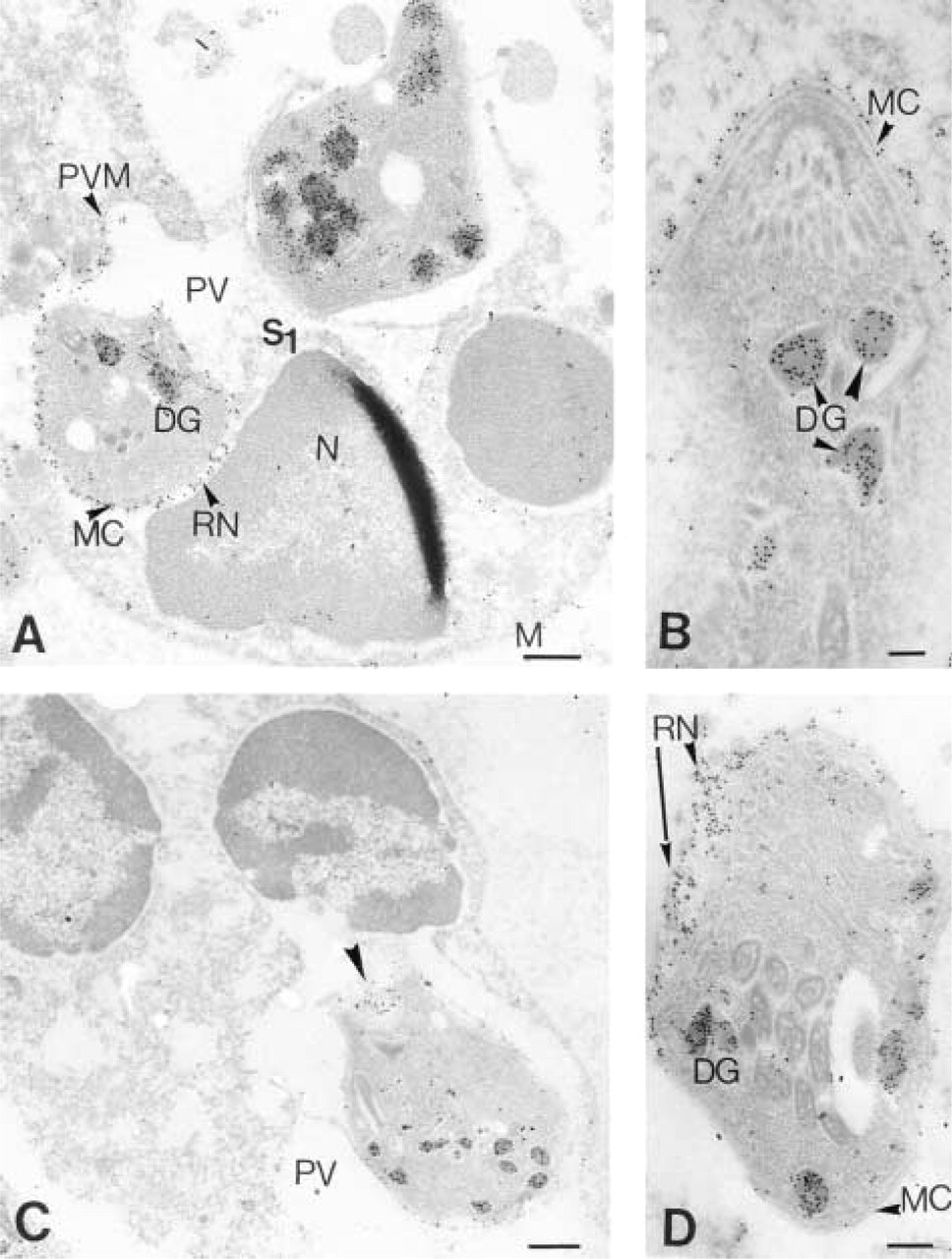
Immuno-EM localization of P29 (GRA7). (
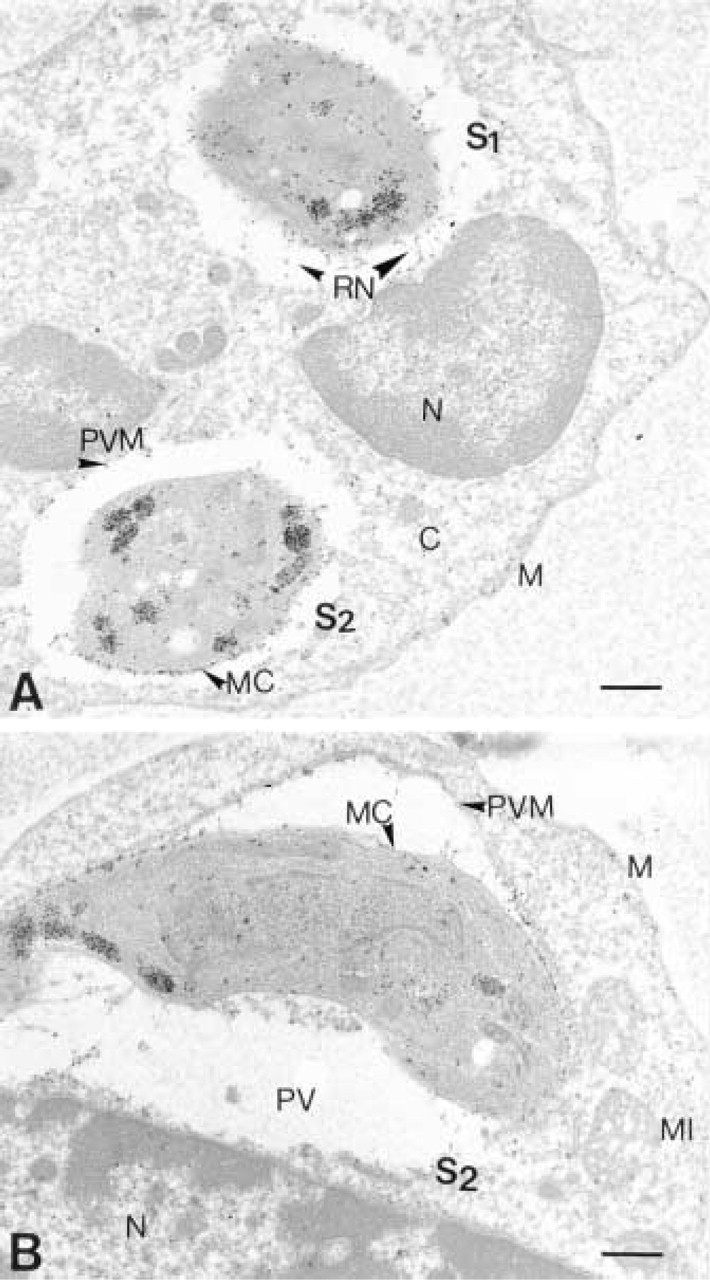
Immuno-EM localization of P29 (GRA7). (
Quantitative P29 immunolabeling analysis in extracellular and intracellular Toxoplasma gondii tachyzoites and in the parasitophorous vacuole (gold particles/μm2) a
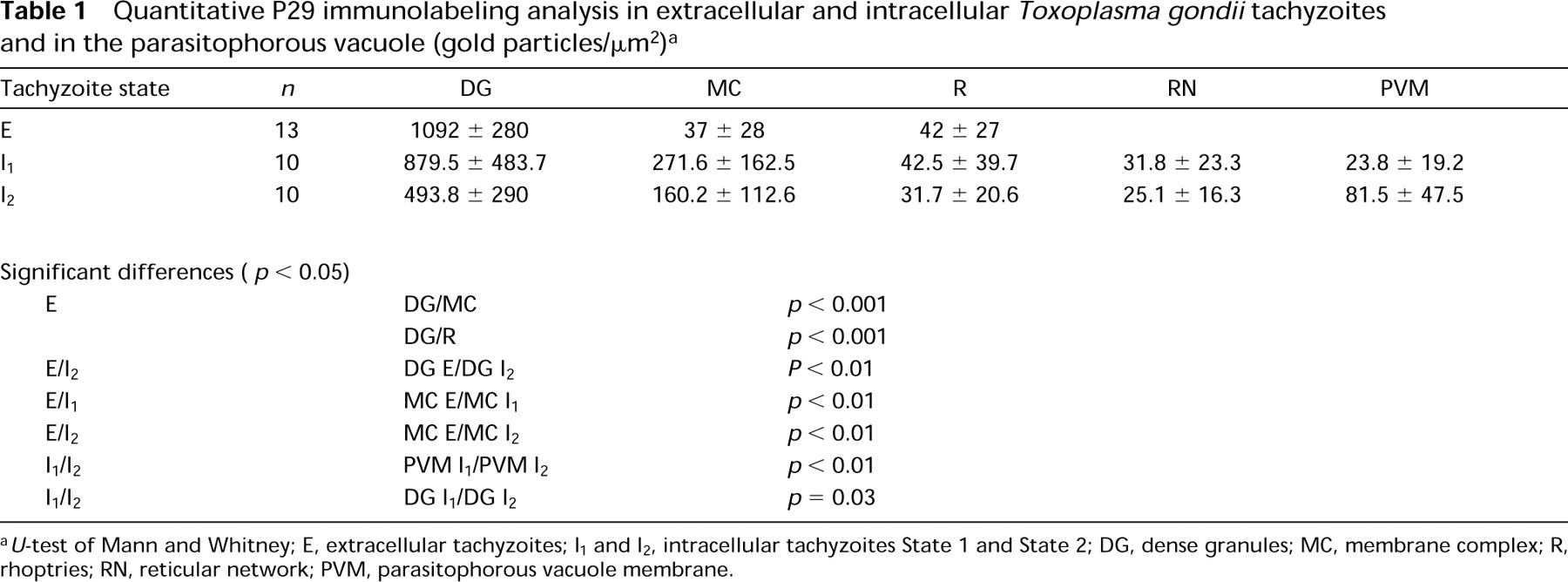
U-test of Mann and Whitney; E, extracellular tachyzoites; I1 and I2, intracellular tachyzoites State 1 and State 2; DG, dense granules; MC, membrane complex; R, rhoptries; RN, reticular network; PVM, parasitophorous vacuole membrane.
Immunolabeling of tachyzoites with the new monoclonal antibody 5-241-178, which binds specifically to the P29 antigen of T. gondii, intensely stained the electron-dense granules and characterized a new dense granule protein of 29 kD termed GRA7. Ultrastructural immunolocalization indicates that after invasion of the host cell by the tachyzoite, the P29 antigen (GRA7) was found associated with the parasite membrane complex and with the tubular elements of the intravacuolar network (State 1) (Figure 3; Table 1). Furthermore, this antigen was also incorporated into the membrane of the parasitophorous vacuole (State 2) (Figure 4). Electron micrographs suggest that the intravacuolar network of membranous tubules is connected in localized regions to the vacuolar membrane. Our immunolabeling data shows trafficking of the P29 antigen from the dense granules via the intravacuolar network to the PVM. We also observed an outward expansion of the P29 antigen from the vacuole into the host cell cytoplasm and into the cell plasma membrane.
Analysis of the primary amino acid sequence determined from the DNA sequence of the gene encoding the P29 (GRA7) antigen reveals several interesting structural features consistent with this antigen being a member of the GRA family. First, the P29 antigen contains a canonical signal peptide at the N-terminus to permit the protein to enter the secretory pathway. Second, this antigen contains two hydrophobic domains of 15 amino acids and 29 amino acids. The larger hydrophobic domain has a high degree of α-helical character and is probably the transmembrane region that anchors the protein to the parasitophorous membrane. Third, this antigen contains a potential N-linked glycosylation site just downstream of this transmembrane region, which may also be important in targeting this protein to the membrane. Fourth, no amino acid homology exists between GRA7 and the other GRA proteins. The structural features of the P29 (GRA7) antigen are similar to those of the GRA4, GRA5, and GRA6 proteins in that all of these proteins contain amino acid sequences that fulfill the requirement of transmembrane regions (Mevelec et al. 1992; Lecordier et al. 1993,1995). In addition, both GRA4 and GRA6 contain a potential N-linked glycosylation site upstream of the transmembrane domain, whereas this site is located downstream from this region in the GRA7 protein. The GRA5 protein contains no potential N-linked glycosylation sites. Of the seven GRA proteins identified to date, only three are localized to the parasitophorous vacuolar membrane: GRA3, GRA5, and GRA7. It would be interesting to examine the interplay among GRA3, GRA5, and GRA7 in the biogenesis of the parasitophorous vacuolar membrane on penetration of the tachyzoite into the host cell.
Note Added in Proof
During review of this work, the same gene and protein, and localization of this protein to the dense granules, have been described by Fischer et al. (1998) and Jacobs et al. (1998).