
Abstract
Serum amyloid A (SAA) is an acute-phase reactant whose level in the blood is elevated to 1000-fold as part of the body's responses to various injuries, including trauma, infection, inflammation, and neoplasia. As an acute-phase reactant, the liver has been considered to be the primary site of expression. However, limited extrahepatic SAA expression was described in mouse tissues and in cells of human atherosclerotic lesions. Here we describe nonradioactive in situ hybridization experiments revealing that the SAA mRNA is widely expressed in many histologically normal human tissues. Expression was localized predominantly to the epithelial components of a variety of tissues, including breast, stomach, small and large intestine, prostate, lung, pancreas, kidney, tonsil, thyroid, pituitary, placenta, skin epidermis, and brain neurons. Expression was also observed in lymphocytes, plasma cells, and endothelial cells. RT-PCR analysis of selected tissues revealed expression of the SAA1, SAA2, and SAA4 genes but not of SAA3, consistent with expression of these genes in the liver. Immunohistochemical staining revealed SAA protein expression that colocalized with SAA mRNA expression. These data indicate local production of the SAA proteins in histologically normal human extrahepatic tissues.
Keywords
S
The properties and the physiological significance of the SAA family of proteins in normal and in disease states are poorly understood. Several functions have been proposed, including depression of the immune responses (Aldo-Benson and Benson 1982), inhibition of platelet aggregation (Zimlichman et al. 1990), involvement in cholesterol metabolism (Kisilevsky and Subrahmanyan 1992), participation in detoxification of endotoxin (Baumberger et al. 1991), induction of collagenase activity (Brinckerhoff et al. 1989), inhibition of the neutrophil oxidative burst (Linke et al. 1991), induction of migration of monocytes, polymorphonuclear leukocytes and T-lymphocytes (Badolato et al. 1994, Xu et al. 1995), and inhibition of cell adhesion to extracellular matrix components (Preciado-Patt et al. 1994). Within this array of possibilities there appears to be a common feature: the effects are compatible with a modulating potential on some of the constituents of the acute-phase reaction and inflammation.
In the present study we describe nonradioactive in situ mRNA hybridization and immunohistochemistry experiments revealing SAA mRNA and protein expression in many histologically normal human extrahepatic tissues, predominantly in their epithelial components. In addition, RT-PCR analysis of these tissues revealed the expression of the SAA1, SAA2, and SAA4 genes but not of the SAA3 gene.
Materials and Methods
Tissues
Multisection slides containing a composite of histologically normal human tissue sections arranged on a grid (Battifora and Mehta 1990) were purchased from Biomeda (Foster City, CA). Each tissue type is represented by two to five sections that were obtained from different individuals (men and women 19-80 years of age). Sections of selected archival tissues, formalin-fixed and paraffin-embedded (breast, prostate, colon, skin, and liver from men and women 39-83 years of age), and freshly frozen tissues (breast, large intestine, spleen, esophagus, and kidney from men and women 50-70 years of age) were obtained from the Department of Pathology, Hadassah University Hospital. The tissue samples (both the commercial samples and those from our archives) were taken from individuals suffering from benign or malignant tumors from areas that appeared histologically normal. In addition, freshly frozen specimens and formalinfixed, paraffin-embedded tissue sections from healthy individuals who underwent cosmetic surgery were also analyzed [breast reduction (four women 31-67 years of age) and skin (a women 70 years old, a man 54 years old)].
Probes
SAA probe was prepared from pGEM transcription vector that contained a 110-
In Situ Hybridization
In situ hybridization was performed as previously described (Urieli-Shoval et al. 1992), with slight modifications. Deparaffinized and rehydrated sections were digested with proteinase K (5 μg/ml, 40 min, 37C) and prehybridized in 100 μl hybridization buffer (50% deionized formamide/0.3 M NaCl/ 20 mM Tris-HCl, pH 8.0/5 mM EDTA/1 X Denhardt's/ 10% dextran sulfate) for 1-2 hr at 50C. Hybridization was initiated by addition of 50 μl hybridization buffer containing the labeled anti-sense probe and was allowed to proceed for 15-18 hr at 50C. Parallel sections were incubated with either the sense probe or the anti-sense probe mixed with a 10-fold excess of nonlabeled anti-sense probe, to be used as negative controls. After hybridization the sections were treated with RNase A (20 μg/ml, 30 min, RT), followed by 0.1 X SSC/0.5% Tween 20 (three times for 40 min, 37C) and then we proceeded with the immunodetection, which was accomplished with the Genius Nonradioactive Nucleic Acid Detection Kit (Boehringer-Mannheim) with modifications. Slides were incubated in 100 mM Tris-HCl, pH 8.0/150 mM MaCl (Buffer 1) containing 2% normal sheep serum and 0.3% Triton X-100 at RT for 30 min. A total of 100 μl of anti-digoxigenin antibody conjugated to alkaline phosphatase (1:1000 dilution in Buffer 1 containing 1% normal sheep serum and 0.3% Triton X-100) was applied to the slides, which were incubated in a humid chamber overnight at 4C. After washing in Buffer 1, color solution (alkaline phosphate substrate kit III; Vector Laboratories, Burlingame, CA) was applied to the slides and incubation was carried out at RT in a dark chamber until a satisfying intensity of color developed. Sections were counterstained with acridine orange/safranin O (Urieli-Shoval et al. 1992).
Immunohistochemistry
Immunohistochemistry was performed using the Histostain- Plus SP kit, which offers superior sensitivity (Zymed Laboratories; San Francisco, CA). Deparaffinized and rehydrated sections were digested with protease (Sigma #P8038; St Louis, MO) (0.1% in PBS, 10 min, RT) and then treated according to the manufacturer's instructions with the ready-touse kit components. The primary antibody used (at 1:20 dilution) was clone “mcl,” which is a monoclonal antibody against amyloid fibril protein AA (Linke 1984) and is of IgG2a isotype (Diagnostic BioSystems; Fremont, CA). Negative controls included replacement of primary antibodies by normal mouse IgG (Zymed Laboratories) or PBS.
RT-PCR
Total RNA from fresh frozen tissues was prepared using Tri Reagent (Molecular Research Center; Cincinnati, OH). Synthesis of cDNA was performed using oligo(dT)12-18 primer and the SuperScript II reverse transcriptase (Gibco BRL; Gaithersburg, MD). Amplification of cDNA was done using Red Hot DNA Polymerase (Advanced Biotechnologies; Surrey, UK) and primers specific for each of the four known human SAA genes; SAA1, 2, 3, and 4. The sequences of the various primers and their specificity were previously described (Urieli-Shoval et al. 1994). Mixtures were incubated in a thermocycler (MJ Research; Watertown, MA) under conditions previously described (Urieli-Shoval et al. 1994). PCR amplification products were sequenced using the above primers and the Taq DyeDeoxy Terminator Cycle sequencing kit (Applied Biosystems; Foster City, CA). Gel analysis of sequencing reactions was carried out on an automated DNA sequencer (model 373, Applied Biosystems) in the sequencing services of the Weizmann Institute (Rehovot, Israel).
Results
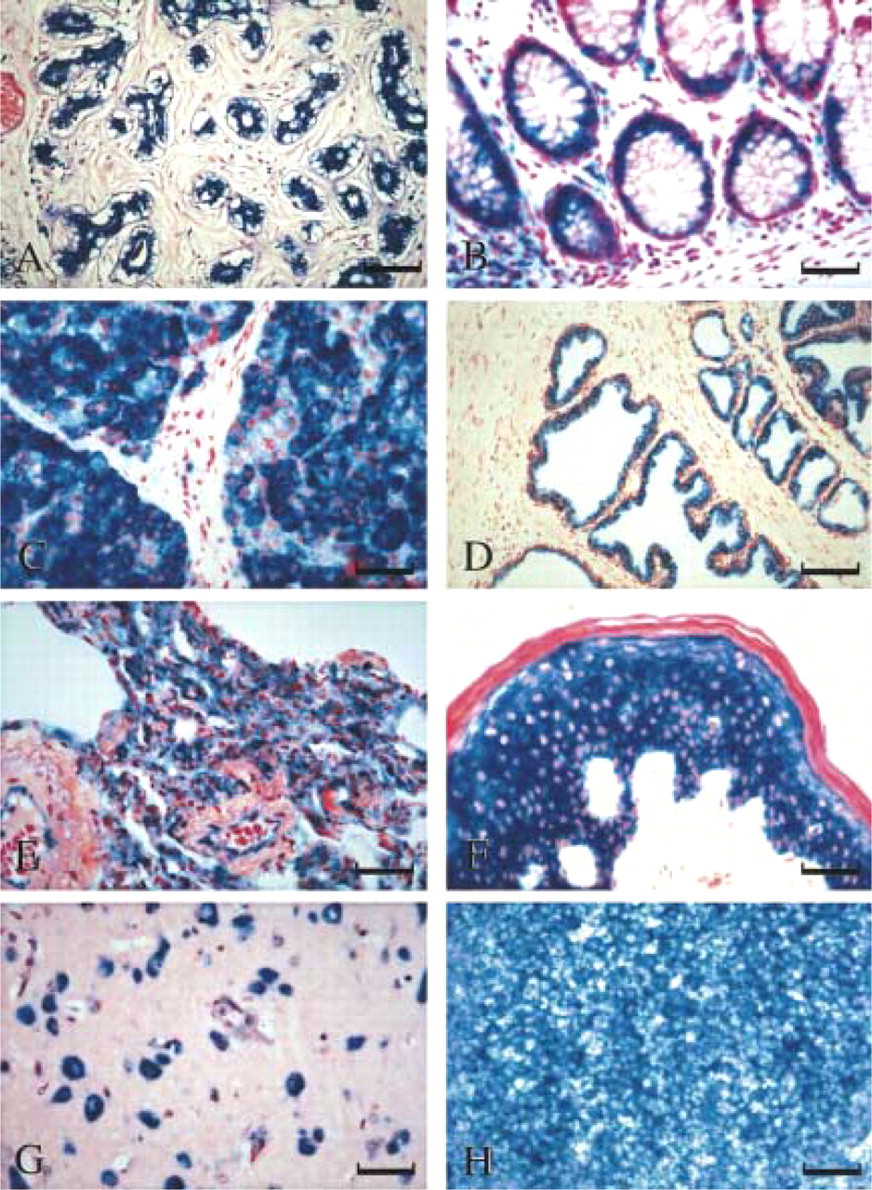
We applied the nonradioactive in situ hybridization technique, using digoxigenin-labeled SAA RNA probe (Urieli-Shoval et al. 1992) on commercial multitissue slides to determine which human tissues express SAA mRNA and what cell types are responsible for such expression. Experiments were repeated at least three times on three different batches of the commercial slides. We initially used these slides that allowed us a large survey of tissues. Experiments were then repeated on selected tissues from our archives (breast, prostate, colon, skin, and liver). Two controls were used for nonspecific hybridization: the sense probe and the anti-sense probe mixed with a 10-fold excess of nonlabeled anti-sense probe. Both resulted in diminished signals. These experiments revealed SAA mRNA expression in many human tissues, predominantly in their epithelial components, as detailed below. Staining of a few selected tissues is shown in Figure 1.
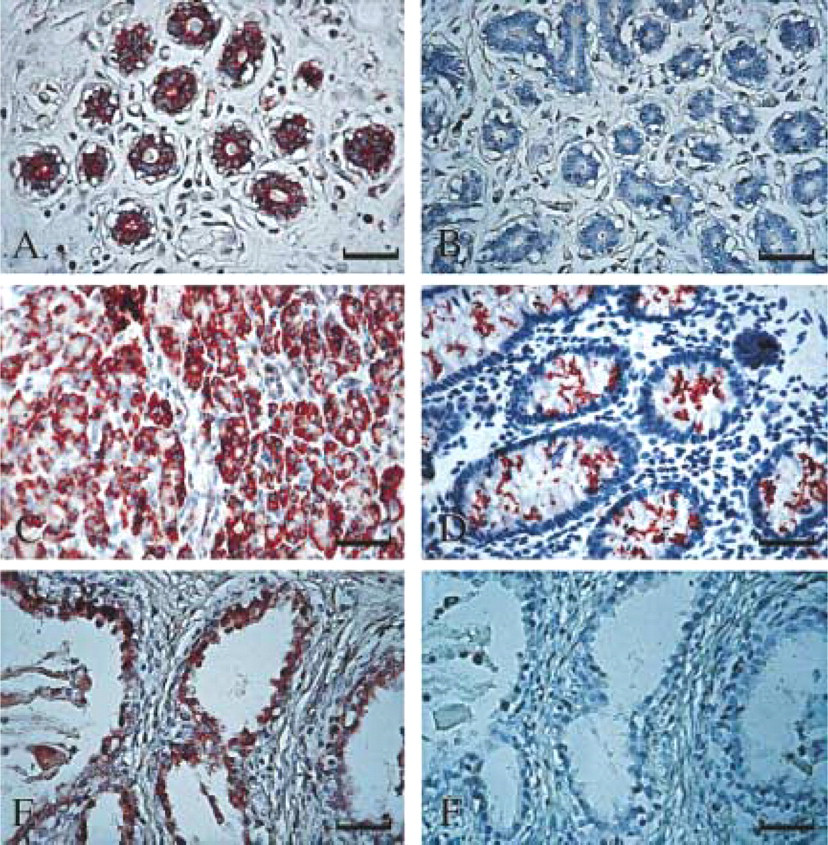
We applied immunohistochemistry to determine whether the presence of SAA mRNA is accompanied by SAA protein synthesis. The antibody used was a monoclonal antibody against human amyloid fibril protein AA (Linke 1984). Negative controls included replacement of the primary antibody by either normal mouse IgG or PBS, both resulting in diminished signal. SAA protein expression was found in most tissues examined, with good correlation to the SAA mRNA expression. Staining of a few selected tissues is shown in Figure 2. The following is a description of our findings.
Breast
Strong in situ hybridization signal was observed in lobular epithelium and terminal duct epithelium; major duct epithelium stained weakly positive. Scattered lymphocytes and plasma cells stained positive. Surrounding loose and dense fibrous connective tissue stroma, fat tissue, and myoepithelial cells were negative (Figure 1A). Immunohistochemistry revealed SAA protein expression co-localized with SAA mRNA expression (Figure 2A). Figure 2B shows a negative control.
Large Intestine, Small Intestine, and Stomach
Positive in situ hybridization signal for SAA was observed in mucosal glandular epithelial cells. Lymphocytes and plasma cells scattered in the lamina propria and lymphocytes in lymphoid follicles of large intestine also stained positively. The smooth muscle of the muscularis mucosa and the loose fibrous connective tissue of the lamina propria and submucosa were negative (Figure 1B). Immunohistochemical staining revealed SAA protein expression, localized to the epithelium (Figure 2D).
Pancreas
Strong in situ hybridization signal was observed in exocrine glandular epithelium; the interlobular duct stained weakly positive. Interlobular septa and interstitial connective tissue were negative. Islets of Langerhans stained weakly positive (Figure 1C). Immunohistochemical staining revealed SAA protein expression localized to the epithelium (Figure 2C).
Prostate
Strong in situ hybridization signal was observed in glandular epithelium. The surrounding fibrous tissue stroma was negative, and scattered lymphocytes and plasma cells in the stroma stained positively (Figure 1D). Cystically dilated glands stained weakly positive. Immunohistochemical staining revealed SAA protein expression localized to the epithelium (Figure 2E). Figure 2F shows a negative control.
Lung
Strong in situ hybridization signal was observed in alveolar lining epithelium and in macrophages. In a medium- sized muscular artery, endothelial cells stained positively and muscle cells were negative. Interstitial connective tissue was negative (Figure 1E).
Skin
Strong in situ hybridization signal was observed in the epidermis (stratified squamous epithelium), whereas stratum corneum and dermis were negative (Figure 1F). In dermal blood vessels, focal positivity of endothelial cells was observed. The skin epidermis also stained positive by immunohistochemistry (not shown).
Brain
In situ hybridization signal was observed in the pyramidal neurons of the cerebral cortex and in Purkinje cells of the cerebellum, whereas non-neural support cells stained negatively (Figure 1G). Neurons also stained positively by immunohistochemistry (not shown).
Nonradioactive in situ hybridization demonstrating SAA mRNA expression in histologically normal human tissues. The blue cytoplasmic staining represents positive SAA signal. (
Immunohistochemistry demonstrating SAA protein expression in histologically normal human tissues. The brown staining represents positive SAA signal. Tissue sections were immunostained with a monoclonal antibody against human amyloid protein AA (
Tonsil and Spleen
Positive in situ hybridization signal was observed in the crypt squamous epithelium of tonsil and in lymphocytes and plasma cells in the underlying stroma. Lymphocytes in lymphoid follicles of the tonsil stained strong positive (Figure 1H), as did the lymphocytes of the white pulp of the spleen (not shown).
In addition, positive staining was observed in the epithelial components of the following tissues: the distal and proximal convoluted tubules of the kidney, follicular epithelium of the thyroid, the secretory cells of the anterior pituitary, and the cytotrophoblast and syncytiotrophoblast lining the chorionic villi of placenta. Liver hepatocytes stained positively (not shown).
RT-PCR Analysis
We carried out RT-PCR analysis on RNA extracted from a few selected tissues to further support the in situ hybridization findings and to determine which of the four known human SAA genes are expressed. Oligonucleotide primers specific for each of the four known human SAA genes, SAA1, SAA2, SAA3, and SAA4, were used. This analysis revealed that fragments of the predicted sizes were amplified when the SAA1-, SAA2-, and SAA4-specific primers were used on RNA extracted from breast tissue. No PCR product was obtained with the SAA3-specific primers (Figure 3). PCR amplification products were sequenced and compared to the SAA sequences reported in the GenBank/EMBL Databases. The breast-derived SAA1 sequence had 99% nucleotide homology with liverderived clones with accession numbers M10906 (clone pA1) and M23698 (Sipe et al. 1985; Kluve-Beckerman et al. 1988). The breast-derived SAA2 sequence had 99% nucleotide homology with clones with accession numbers X51445, M26152 (clones pAS8, pSAA82) (Steinkasserer et al. unpublished observations; Kluve- Beckerman et al. 1986). The breast-derived SAA4 sequence had 99% nucleotide homology with a clone with accession number M81349 (Whitehead et al. 1992). RT-PCR analysis of RNA from other normal tissues, i.e., large intestine, spleen, kidney, and esophagus, also revealed expression of the SAA1, 2, and 4 genes. In some cases the PCR product representing the SAA2 gene was very weak to barely detectable (not shown). A summary of the tissues examined by the three methods described is presented in Table 1.
Discussion
Serum amyloid A is an acute-phase protein and has been considered to be expressed primarily in the liver. For many years, studies using animal or cultured cell models focused on investigating SAA expression and regulation in liver cells and on its possible function as a modulator of the acute-phase reaction and inflammation. Extrahepatic expression of the SAA genes was described, to a limited extent, in mouse tissues (Ramadori et al. 1985; Meek and Benditt 1986; Meek et al. 1989b) and, more recently, in cells of human atherosclerotic lesions (Meek et al. 1994). In the present study we demonstrate for the first time that the SAA mRNA (Figure 1) and protein (Figure 2) are widely expressed in many histologically normal human tissues, including stomach, small and large intestine, tonsil, breast, prostate, thyroid, lung, pancreas, kidney, skin epidermis, and brain neurons (Table 1). SAA expression was localized predominantly to the epithelial components of these tissues. Expression was also observed in lymphocytes and plasma cells scattered in the stroma of many tissues and in lymphoid follicles. The expression in endothelial cells lining the blood vessels, previously described (Meek et al. 1994), was also confirmed.
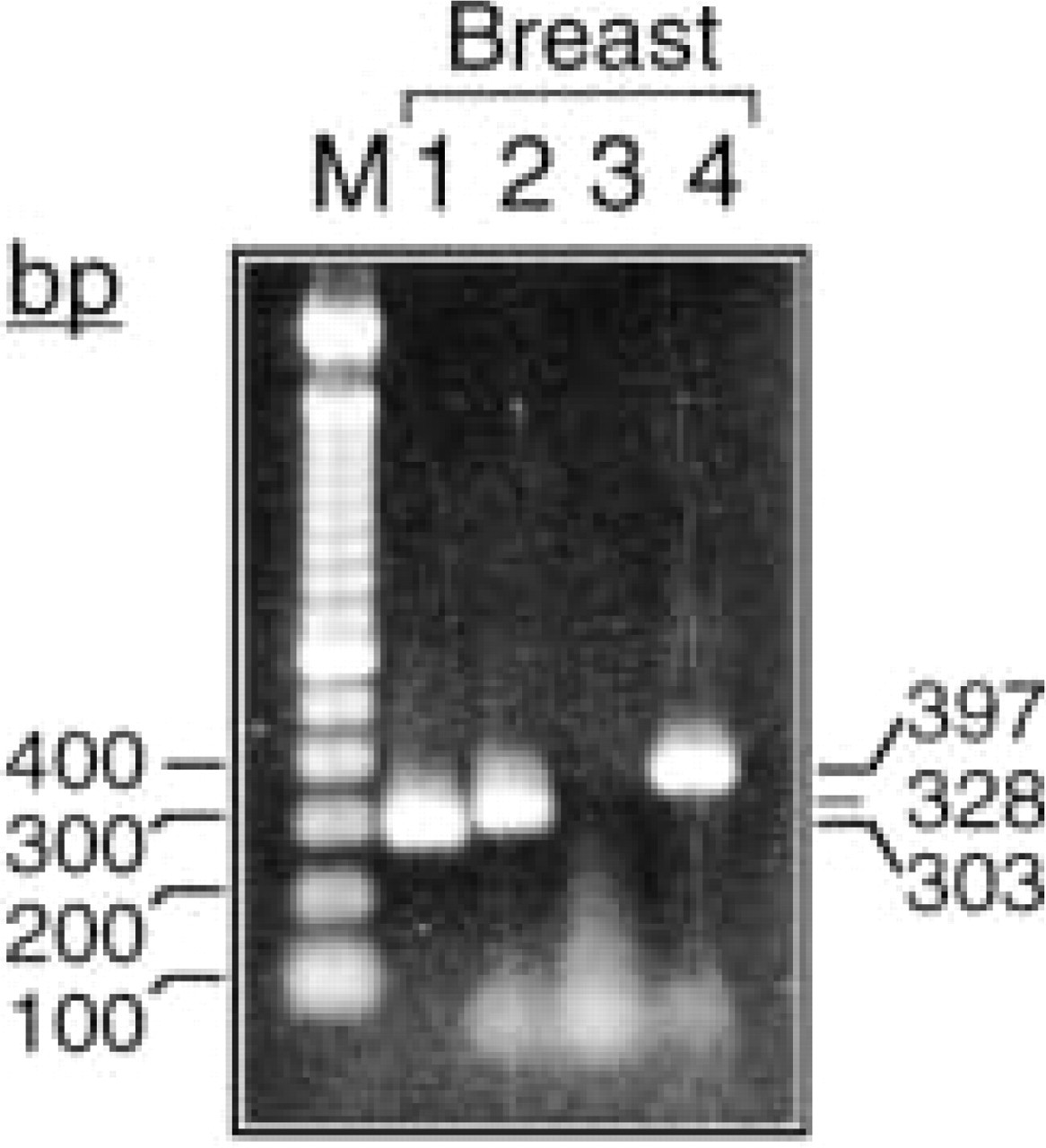
RT-PCR analysis demonstrating expression of the SAA1, SAA2, and SAA4 genes in human breast. The analysis was performed on total RNA using primers distinguishing among the four known human SAA genes. PCR fragments were analyzed on a 2% agarose gel, stained with ethidium bromide, and photographed. Sizes of amplified fragments are indicated along right margins. Markers of a DNA ladder (100-
SAA expression in histologically normal human tissues
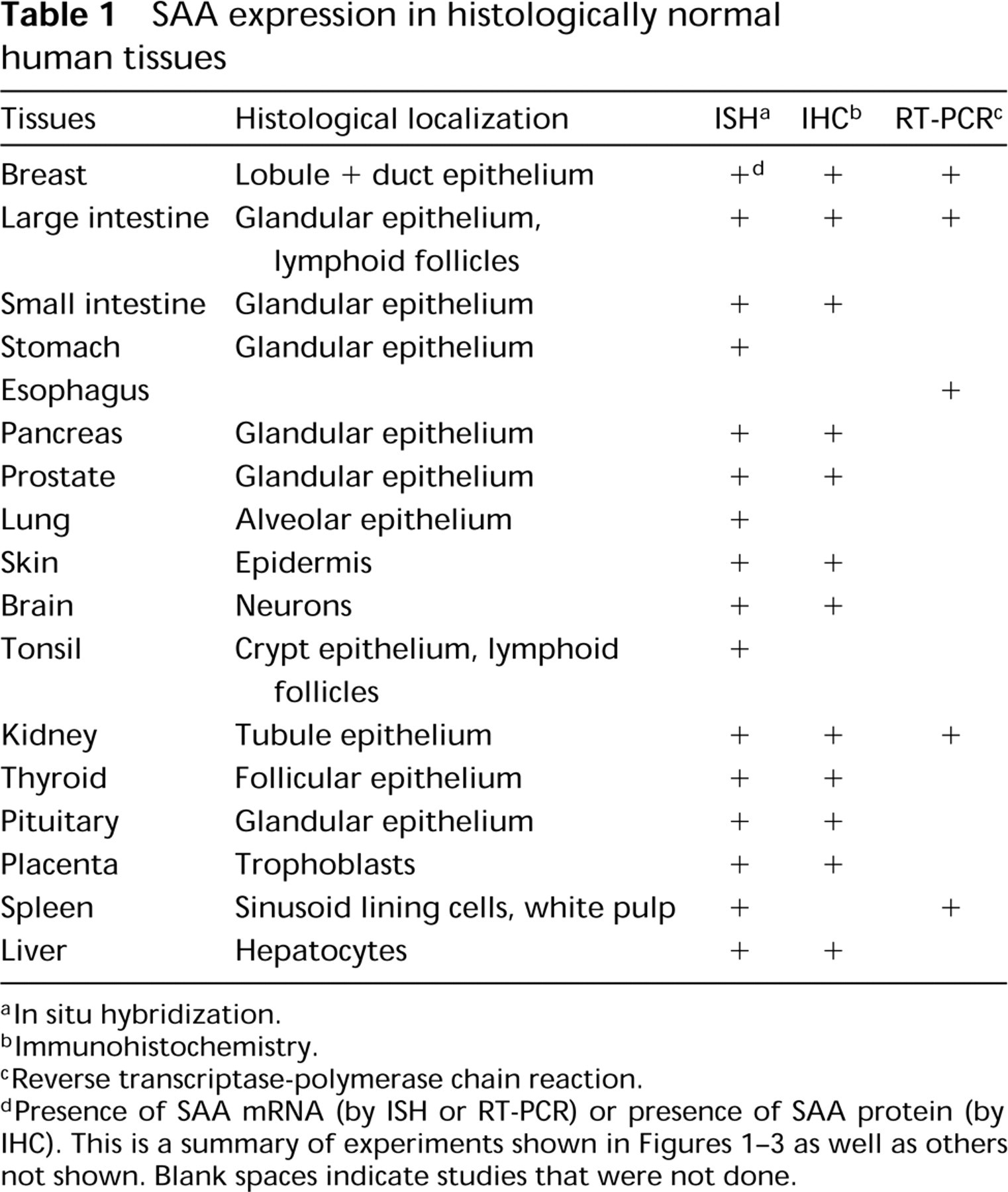
In situ hybridization.
Immunohistochemistry.
Reverse transcriptase-polymerase chain reaction.
Presence of SAA mRNA (by ISH or RT-PCR) or presence of SAA protein (by IHC). This is a summary of experiments shown in Figures 1-3 as well as others not shown. Blank spaces indicate studies that were not done.
The role of SAA expressed in the epithelial components of human tissues is not clear. However, epithelium- specific gene expression is fundamental to both embryogenesis and the maintenance and function of adult tissues, and its impairment might be associated with diseases such as cancer. Our survey revealed that SAA is expressed in tissues of different epithelial types and degree of differentiation. Therefore, SAA may have a role in the maintenance of the epithelial phenotype similar to other epithelium-specific genes such as keratins or junction proteins (Birchmeier and Behrens 1994; Kouklis et al. 1994). In addition, the SAA expressed on the surface of epithelia communicating with the external environment, such as the intestinal tract or lung airways, may be part of the host defense mechanism that protects these tissues from microorganisms and potential injuries. For example, the inhibitory effect of SAA on O2 - production by neutrophils (Linke et al. 1991; Gatt et al. in press) may help to prevent oxidative tissue damage. It should be noted that the differences in the amino acid sequence of the three expressed SAA proteins suggest that each SAA protein may be adapted to a particular function, as previously observed in products of other conserved polygene families (Poyart et al. 1992).
The human SAA gene family is composed of four discrete loci containing two highly homologous genes, SAA1 and SAA2, and two less related genes, SAA3 and SAA4. SAA1 and SAA2 corresponding proteins are the predominant circulating SAA proteins during the acute-phase response (Uhlar et al. 1994; Betts et al. 1991). The SAA3 gene was reported as a nonexpressed gene (Kluve-Beckerman et al. 1991). The gene for SAA4 is constitutively expressed and its protein product is a constituent of normal, non-acute-phase high-density lipoprotein (Whitehead et al. 1992; Steel et al. 1993). Expression of the genes SAA1, SAA2, and SAA4 can be induced in cultured human smooth muscle cells (Meek et al. 1994) and in human monocyte/ macrophage cell lines (Urieli-Shoval et al. 1994). In the mouse, whereas all the four SAA genes are expressed in liver, only one gene (SAA3) is almost exclusively expressed in extrahepatic tissues (Meek and Benditt 1986). Here we show that histologically normal human extrahepatic tissues, including breast (Figure 3), express all three expressed genes, both the “acute-phase” genes SAA1 and SAA2 and the “constitutive” gene SAA4. The significance of these findings requires further investigation.
The study was first done on commercial multitissue slides, enabling us to perform a large survey of tissues. Later, experiments were performed on histologically normal tissues from our archives (either fixed-embedded tissues for in situ hybridization and immunohistochemistry or freshly frozen tissues for RT-PCR), revealing similar results. Furthermore, SAA mRNA and protein were also identified in tissues obtained from healthy individuals who underwent cosmetic procedures (breast, skin), suggesting epithelial SAA expression in the absence of a systemic acute-phase reaction. These results are in line with those of Meek et al. (1989b) and Steel et al. (1996), who described SAA mRNA and protein expression in epithelial cells of certain mouse tissues (kidney, large intestine) and in various human epithelial cell lines.
In conclusion, the concept that SAA is expressed primarily in the liver can now be broadened because there is clearly SAA expression in human extrahepatic tissues. Therefore, in addition to SAA release to the circulation by the liver during an acute-phase response, one or more SAA proteins produced in human tissues may be constitutively expressed to serve as a first line of defense or in proper maintenance of human tissues (housekeeping role). The possibility that local SAA plays a role in the development of AA amyloidosis requires further investigation.
Footnotes
Acknowledgments
Supported in part by the Israel Ministry of Absorption and the Israel Cancer Association.
We thank Dr R.L. Meek (the Heart Institute of Spokane) for the p125 cDNA clone, and Dr Y. Azar (Hadassah University Hospital) and Dr R.P. Linke (Max-Planck Institute) for helpful discussions.
Dedicated to the memory of the late Earl P. Benditt, MD.