
Abstract
Analysis of the connectivity between different neuronal cell types is dependent on an appreciation of their dendritic and axonal arborizations. A detailed study of the dendrites and axons of GABAergic neurons has been thwarted by the lack of a suitable technique for enhancing GABA immunoreactivity. This article describes a procedure using tetanus toxin which, when applied to organotypic hippocampal cultures, considerably enhances the immunoreactivity in the dendrites and axons of the GABA- and somatostatin-containing neurons and clearly demonstrates the co-localization of GABA and somatostatin immunoreactivities in the same neuron. Tetanus toxin was applied to the culture medium on Day 14 for a 24-hr period and the cultures were fixed at the end of Day 18. Tetanus toxin-treated cultures (n = 30) or untreated cultures (n = 40) were incubated for either GABA or somatostatin immunoreactivity. Tetanus toxin-treated cultures used for co-localization studies (n = 20) were incubated for both GABA and somatostatin immunoreactivity.
T
In the hippocampus, knowledge about the connectivity of the nonpyramidal, presumed GABAergic inhibitory neurons awaited the development of immunocytochemical methods that utilized antisera directed against glutamic acid decarboxylase (GAD) (Ribak et al. 1981; Somogyi et al. 1983). Somewhat later, antisera against 7-aminobutyric acid (GABA) became available and confirmed the observations made by previous workers using GAD (Storm–Mathisen et al. 1983; Somogyi et al. 1985). Although the GABAergic somata and, in some cases, their terminal fields were demonstrated, the detailed dendritic and axonal morphology of these neurons was still lacking (Freund and Buzsaki 1996). The best evidence for the detailed connectivity of individual GABAergic neurons was achieved by the elegant studies in which Golgi-type impregnations were combined with GABA or GAD immunocytochemistry (Somogyi and Hodgson 1985; Soriano and Frotscher 1989).
A method is needed that enhances or amplifies the antigenic signal and that might allow a more thorough examination of the connectivity between GABAergic neurons and other neurons.
Previous work from this laboratory showed that, when tetanus toxin was administered in vivo, a considerably enhanced immunoreactivity was observed in neurons containing somatostatin (Mitchell et al. 1995). Because tetanus toxin blocks neurotransmitter release, it might also lead to an increase in the GABAergic content of the cell and thus reveal a more detailed morphology of the GABAergic neurons. In the present study tetanus toxin was applied to organotypic cultures of the rat hippocampus which were subsequently processed for detection of GABA and for somatostatin, a peptide known to be present in GABAergic neurons (Somogyi et al. 1985).
Materials and Methods
Hippocampi were dissected from 8-day-old neonatal rats, placed in Geys medium supplemented with 28 mM glucose, and sliced on a McIlwain tissue chopper set to cut at 400 μm (Stoppini et al.1991). Slices placed on Millicell-CM membranes (Millipore; Bedford, MA) were incubated with 1.2 ml of medium comprising Eagle MEM (50%), Hanks BSS (25%), horse serum (25%), D-glucose (5 mg/ml), L-glutamine (1 mM). The medium was replaced every 3 days. On Day 14 of culture, tetanus toxin, [Alamone Labs; Jerusalem, Israel (N.B. Not the more readily available tetanus toxoid)] was added to the culture medium to give a final concentration of 20 ng/ml for a 24-hr period (this optimal concentration of tetanus toxin was determined from an unpublished pilot study). Tetanus toxin was added to 50 cultures (20 of which were used for co-localization studies). All cultures were fixed in 4% paraformaldehyde in 0.1 M phosphate buffer, pH 7.2, on Day 18, for 24 hr. Cultures were processed for immunocytochemistry.
The immunocytochemical procedure used to reveal somatostatin and GABA immunoreactivity was a modification of that used previously (Mitchell et al. 1995) but with increased incubation times to allow antibody penetration into the tissue. Briefly, endogenous tissue peroxidase was blocked by a 30-min incubation in hydrogen peroxide (0.05%) in methanol (10% in Tris-buffered saline, TBS). Incubation times were as follows: mouse anti-somatostatin (Chemicon, Temecula, CA; 1:5000 dilution, 72 hr); rabbit anti-GABA (Sigma, St Louis, MO; 1:3000 dilution, 72 hr); biotinylated horse anti-mouse or biotinylated goat anti-rabbit IgG (18 hr); avidin–biotin–HRP complex (18 hr). Somatostatin immunoreactivity was revealed with diaminobenzidine tetrahy-drochloride (DAB) 30 min, DAB and hydrogen peroxide (5 min) to give a brown reaction product and GABA immunoreactivity with Vector SG peroxidase substrate (Vector; Burlingame, CA) to give a blue final reaction product. All incubations were carried out in TBS to which Triton X-100 (0.3%) had been added. The sections received three 1-hr washes between each incubation. For co-localization studies, cultures (n = 20) were incubated for somatostatin immunoreactivity followed by incubation for GABA immunoreactivity.
Controls
Specific immunoreactivity for GABA was blocked either by treating cultures (n = 10) with GABA antibody that had been preadsorbed with GABA antigen coupled to polacrylamide beads (Ternyck and Avrameas 1972) or by replacing the primary antibody with TBS (n = 10). Somatostatin immunoreactivity was blocked by preadsorbing the antiserum with 10 nM somatostatin (n = 10). Cultures without tetanus toxin were also incubated for GABA (n = 20) or somatostatin (n = 20) immunoreactivity.
Results
GABA
Control cultures incubated in the presence of the preadsorbed GABA antibody or in the absence of the GABA antibody revealed no staining of GABAergic neurons. In cultures not exposed to tetanus toxin (Figure 1A), only the weakly stained somata of the GABAergic neurons were present.
Intensely immunoreactive GABAergic neurons were located throughout the septotemporal extent of the hippocampus, in all layers of the hippocampus proper, and in the dentate gyrus. Many neurons in strata oriens, pyramidale, and in the subiculum revealed extensive dendritic and axonal arborizations (Figures 1B and 1C) and an intense terminal staining around the principal cells in stratum pyramidale. A similar GABAergic immunoreactivity was observed in the basket cells of the dentate gyrus, whose dendrites extended deep into the molecular layers and whose axons revealed an intense terminal plexus around the granule cells.
Somatostatin
Control cultures incubated for somatostatin immunoreactivity in the absence of tetanus toxin revealed the characteristic signet-ring staining of their cytoplasm (Figure 2B), and those cultures incubated with preadsorbed antibody revealed no staining.
After tetanus toxin exposure, the somatostatin cells became intensely labeled and revealed not only increased staining of dendrites but also of axons and terminals. Particularly striking were the somatostatin neurons in stratum oriens (Figure 2C), with axons that extend vertically down through the pyramidal cells into stratum radiatum to terminate in stratum lacunosum moleculare. Somatostatin neurons with long dendrites and axons were also observed in the hilus of the dentate gyrus (Figure 1D). Most of these axons were confined to the hilar area and terminated on soma-tostatin-negative cells.
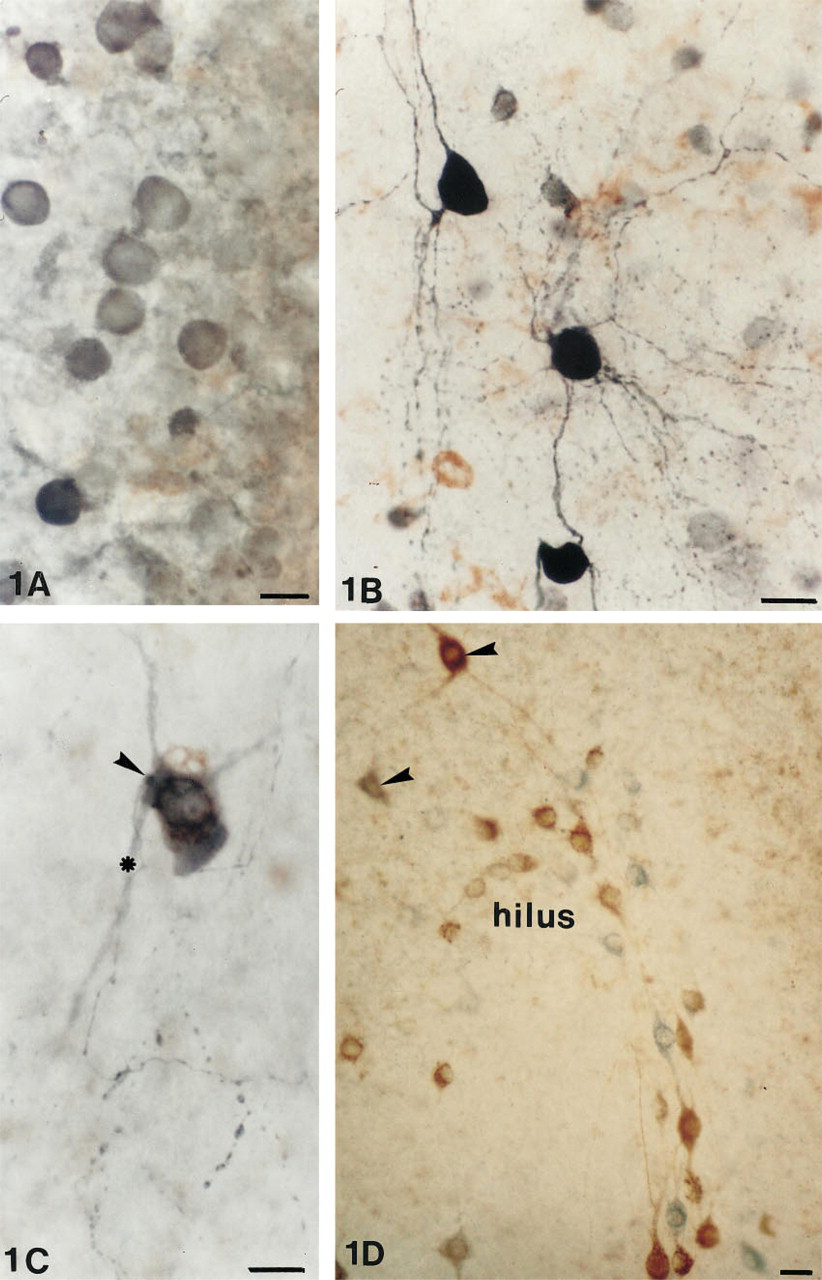
GABAergic neurons in untreated (
Many neurons in stratum oriens were intensely immunoreactive for GABA and somatostatin and were stained a deep blue-black color. In stratum radiatum throughout the hippocampus were isolated neurons, whose somata measured 30–35 μm in diameter, which co-localized somatostatin and GABA. The brown DAB reaction product indicative of somatostatin was quite distinct from the blue SG reaction product demonstrating GABA (Figure 1C). The beaded axons of these neurons could be followed into the dentate gyrus. In the dentate gyrus (Figure 1D), as in stratum oriens, many neurons were immunoreactive for either somatostatin or GABA alone. Many hilar cells were somatostatin-immunoreactive but GABA-negative. However, there were neurons in the hilus that co-localized somatostatin and GABA immunoreactivities (Figure 1D).
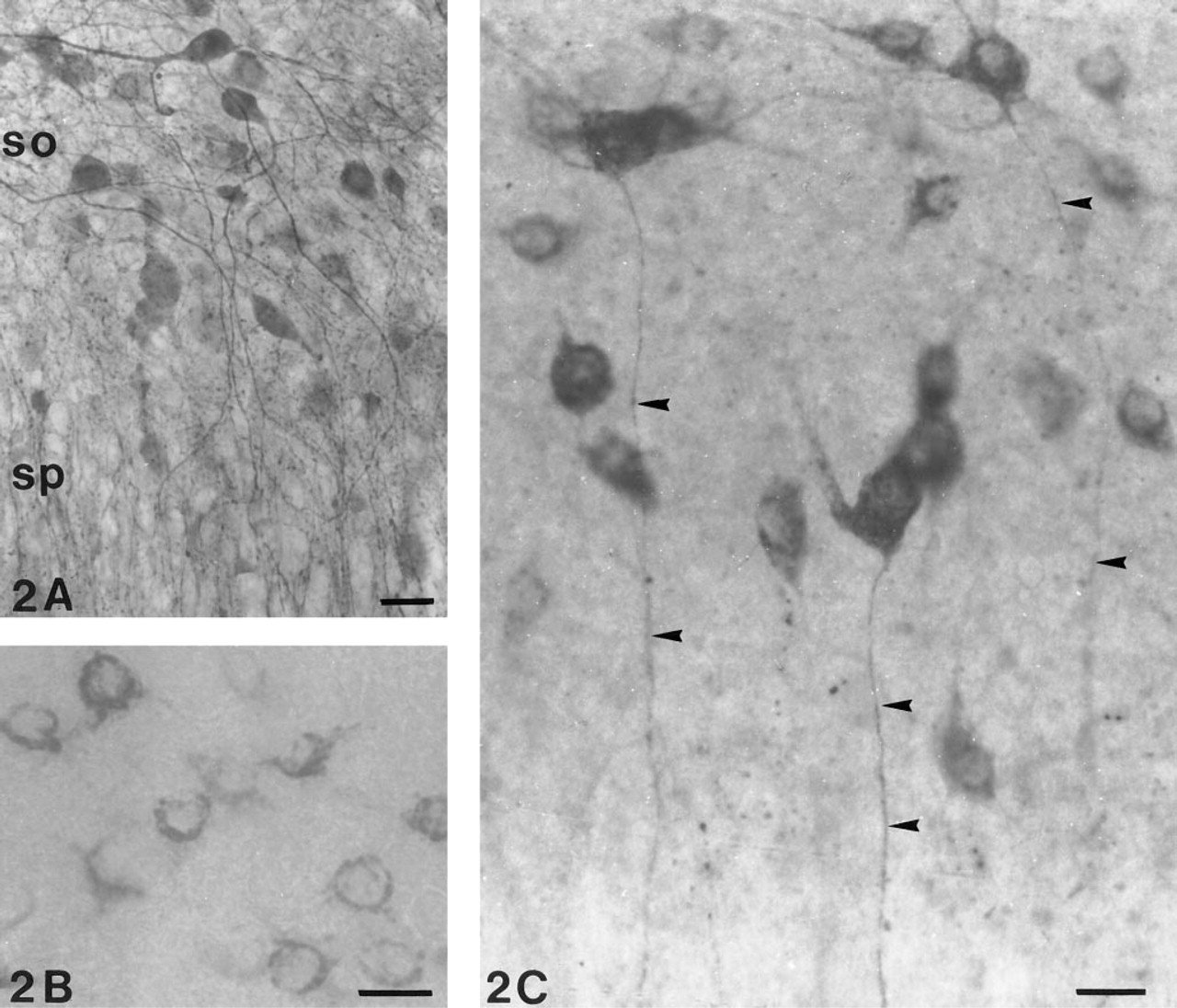
GABAergic neurons in stratum oriens (so) from the temporal pole of the hippocampus. sp, stratum pyramidale. (
Discussion
The present study has clearly demonstrated that a single dose of tetanus toxin enhances the immunoreactivity of the dendrites and axons of GABAergic and peptidergic hippocampal neurons in culture.
Although the preparation of the organotypic cultures causes the loss of all extrinsic afferents and results in some reorganization (Zimmer and Gähwiler 1984), the advantages of their use far outweigh any disadvantages (Gähwiler 1981,1984; Stoppini et al. 1991; Torp et al. 1992). Immunocytochemical and in situ hybridization studies clearly show that interneurons maintain a similar cytoarchitecture to their counterparts in vivo (Finsen et al. 1992; Torp et al. 1992; Best et al. 1994; Kunkel et al. 1994; Mitchell et al. 1996), and functionally have an active role in the inhibitory control of the principal neuronal populations (Gähwiler and Dreifuss 1982; Gähwiler 1984).
It is partly because these cultures maintain a structure and function similar to that seen in vivo and partly because of the large number of slices that can be obtained from each hippocampus that they are increasingly being used as in vitro models to investigate the pathophysiology of amino acid-based neurotrans-mission in conditions such as hypoxia and epilepsy (Benediktz et al. 1993; Casaccia–Bonnefil et al. 1993; Best et al. 1994,1996). In the present study they were used to circumvent the use of tetanus toxin in live animals which, in some cases, would have experienced intense seizure activity. Detailed information about the dendrites and axons of GABAergic neurons is limited (Freund and Buzsaki 1996). However, in the present study, the dendrites, axons, and terminals of the GABAergic neurons around the pyramidal and granule cells in the hippocampus proper and in the dentate gyrus were intensely immunoreactive and clearly visible. The dendrites and axons of the GABAergic cells in the subiculum were similar to those observed in Golgiimpregnated neurons.
The somatostatin immunoreactive neurons in the hippocampus and dentate gyrus of the present study are similar to those described previously (Kohler and Chan–Palay 1982; Kunkel et al. 1994). Those neurons with dendrites confined to stratum oriens and with axons that traverse the pyramidal cell to terminate in stratum lacunosum moleculare are similar to those described by McBain et al. (1994) and Sik et al. (1995), using intracellular studies.
The GABAergic neurons that also contained somatostatin and were found in stratum radiatum close to the border with lacunosum moleculare and whose axons could be followed into the dentate gyrus resemble the NPY neurons described previously in organotypic cultures and in vivo (Mitchell et al. 1996). Somatostatin and NPY are often co-localized in the same neuron, both in the hippocampus proper and in the dentate gyrus (Kohler et al. 1986).
Indirect evidence from intracellular labeling studies and from immunostaining against receptors associated with somatostatin neurons would confirm that the dendrites of hilar somatostatin neurons remain within the hilus and run parallel with the dentate granule cells (see review by Freund and Buzsaki 1996). The majority of axons from the hilar somatostatin neurons appear to be confined to the hilus, with only an isolated axon observed to pass through the granule cell layer into the outer molecular layer. This appears to contradict indirect evidence that axons from the hilar somatostatin neurons are responsible for the dense terminal plexus observed in the outer molecular layer (Bakst et al. 1986). However, the elegant studies of Han et al. (1993) demonstrated that the terminal plexus arising from a single hilar neuron (considered to be similar to a somatostatin neuron) spreads over a distance of 2.0 mm, i.e., corresponding to approximately two thirds the longitudinal length of the outer molecular layer. The majority of hilar somatostatin cells referred to above also co-localized GABA.
In summary, tetanus toxin enhances the immunore-activity of GABAergic dendrites and axons, which should facilitate an analysis of their connectivity and provide a better understanding of their function. Tetanus toxin may also facilitate the localization of other neurotransmitters in alternative culture systems.
Footnotes
Acknowledgements
Supported by Action Research Grant S/P 2796. I thank Prof Udo Schumacher and Dr Nicholas Best for their valued comments.