
Abstract
Thyroid hormone (T3) affects muscle development and muscle regeneration. It also interacts with the muscle regulatory gene MyoD in culture and affects myoblast proliferation. We studied the localization of MyoD protein using a well-characterized polyclonal antibody for immunohistochemistry. Relative numbers of myogenic precursor cells per field were identified by their MyoD expression during muscle regeneration in normal and mdx dystrophic mice, with particular reference to the expression in mononuclear cells and myotubes at various T3 levels. In regeneration by normal muscles, relatively few MyoD+ nuclei per field were present in mononuclear cells of euthyroid and hypothyroid mice. MyoD staining of mononuclear cell nuclei was approximately doubled in fields of regenerating muscles of normal hyperthyroid compared to euthyroid mice, and was observed in precursors that appeared to be aligned before fusion into myotubes. In mdx regenerating muscle, twofold more mononuclear cells positive for MyoD were present in all three treatment groups compared to normal muscles regenerating under the same conditions. Localization was similar to the pattern in normal euthyroid mice. However, in muscles regenerating in hyperthyroid mdx mice, both mononuclear cell nuclei and centrally located nuclei in a subpopulation (about 15%) of new myotubes formed after the crush injury were intensely stained for MyoD protein. The changes observed are consistent with reports on T3-induced alteration of muscle repair, and propose a link between MyoD regulation and the accelerated differentiation during regeneration under high T3 conditions.
T
To further characterize the cellular events associated with the effects of T3, we analyzed the expression of MyoD protein during repair in vivo in normal and mdx mice using immunocytochemistry. MyoD is one of four muscle regulatory factors (MRFs) that direct the specification of mesodermal cell transition into the muscle lineage (Weintraub 1993; Megeney and Rudnicki 1995; Cossu et al. 1996; Yun and Wold 1996). In muscle development, MyoD is expressed early in the sequence of the myogenic lineage, whereas myogenin is expressed later as myoblasts progress through the differentiation program. The MRFs are also detected in myogenic cultures (Wright et al. 1989; Hinterberger et al. 1991; Smith et al. 1993; Maley et al. 1994). This expression by cells already committed to the muscle lineage likely reflects the role of MRFs in the transition from proliferation to differentiation (Olson 1992, 1993; Weintraub 1993). As in development, during myogensis in culture, MyoD expression precedes myogenin expression (Hinterberger et al. 1991; Yablonka–Reuveni and Rivera 1994,1997). Cell culture studies also noted that MyoD is first seen in proliferating myoblasts (Tapscott et al. 1988; Yablonka—Reuveni and Rivera 1994,1997). MyoD expression in regenerating muscle occurs shortly after injury in mononuclear cells, as studied at the transcriptional level using in situ hybridization (Grounds et al. 1992) and at the protein level using immunocytochemistry (Füchtbauer and Westphal 1992). Analysis at the mRNA level also revealed a similar distribution of MyoD+ and myogenin+ myoblasts in the regenerating areas observed in normal (Grounds et al. 1992) and dystrophic muscles (Garrett and Anderson 1995). Therefore, MyoD is one of the early markers for myogenic cells in vivo (Grounds et al. 1992) and in culture (Yablonka–Reuveni and Rivera 1994). Moreover, T3 was shown to increase the expression of MyoD in culture (Carnac et al. 1992). We were therefore interested to use immunostaining for MyoD to identify relative numbers of myoblasts during muscle repair in vivo, especially in response to various levels of T3. On the basis of previous studies on normal C57 and mdx dystrophic mice, our hypothesis was that myoblasts expressing MyoD would be present in greater numbers per field in mdx dystrophic than normal regenerating muscles. We also hypothesized that MyoD+ mononuclear cell nuclei would be observed in greater relative numbers in regenerating muscles after treatment with T3 compared to regenerating muscles in untreated mice or those with low thyroid hormone levels, but that the pattern of expression during muscle differentiation would be similar between the two strains.
Materials and Methods
Animals and Thyroid Hormone Experiments
Normal control (C57Bl/10 ScSn) mice and the mdx mutant strain (C57Bl/10ScSn mdx) were bred and maintained in the Central Animal Care Facility at the University of Manitoba, on a 12:12-hr light: dark schedule. All treatments and surgical procedures were approved by the Animal Protocol Committee of the University of Manitoba.
Hypothyroidism was produced in normal control and mdx dystrophic mice as reported (McIntosh et al. 1994) by consumption of propylthiouracil (0.05%) in drinking water for 8 weeks, starting at 3 weeks of age. Hyperthyroidism was induced by injecting triiodothyronine (T3) sc at a dose of 2 μg/g body weight/day (Anderson et al. 1994; Pernitsky et al. 1996) for 4 weeks, starting at 5.5 weeks of age. Non-dystrophic normal control and mdx littermate mice were left untreated (euthyroid) for the same periods for each condition, respectively. For each of the treatment groups (hypothyroid, euthyroid, hyperthyroid) of normal and dystrophic animals, there were four to nine animals per group. The variation in group size was due to the examination of sections derived from more than one previous experiment.
Four days before sacrifice, anesthetized mice received a crush injury to the right tibialis anterior (TA) muscle as previously described (McIntosh et al. 1994, after Grounds and McGeachie 1989). The right TA was subject to crush injury and the left TA was only exposed by a skin incision but the muscle remained intact and served as a control muscle. After 4 days of recovery, the crushed and intact TA muscles were dissected from the hindlimbs, embedded together in OCT (Miles; Elkhart, IN) for longitudinal or cross-sections, and frozen for cryosectioning (7-μm-thick sections). The TA muscle was chosen for the present investigations because sections were available from animals in all three treatment groups and from both strains. The TA is often used for studies of muscle regeneration because it lies subcutaneously and is easily accessible to surgical or other interventions applied to stimulate muscle repair. The TA is also a large muscle relative to other muscles, such as the extensor digitorum longus muscle, and is quite homogeneous and therefore representative of fast-twitch muscles. The sequence and timing of the regeneration process in the TA muscle are very well characterized (e.g., Grounds and McGeachie 1989). In particular, the interval of 4 days post crush injury was chosen in accordance with previous detailed reports of morphometric and autoradiographic studies that documented myoblast proliferation and fusion, and myotube growth in normal and mdx regenerating muscles in our laboratory (McIntosh et al. 1994; Anderson et al. 1995; McIntosh and Anderson 1995; Pernitsky et al. 1996) and reports from other investigators (e.g., Grounds et al. 1992; Mitchell et al. 1992).
Immunohistochemistry
Sections consisting of pairs of crushed and uncrushed muscle were immunostained for MyoD protein using standard protocols for immunofluorescence. The antibody against MyoD was a polyclonal antibody raised in rabbit, and was a generous gift from Dr. S. Alemà (Institute of Cell Biology, CNR, Rome, Italy). Its characteristics were reported previously in studies of myogenesis of rat satellite cells (Yablonka–Reuveni and Rivera 1994) and mouse-derived myogenic cultures (Yablonka–Reuveni and Rivera 1997). As described in both reports, the antibody stains nuclei of mouse and rat myogenic cultures. In cultures, the staining pattern with the polyclonal antibody against MyoD is identical to that of a mouse monoclonal antibody against MyoD (clone 5.8A, originally reported by Dias et al. 1992), as concluded from double immunostaining with the polyclonal and monoclonal antibodies of myogenic cultures of mouse and rat origin. However, we found the polyclonal antibody was a better tool for recognizing MyoD+ nuclei in vivo because it gave a more intense stain and had lower background. High background staining is a general problem with mouse monoclonals used to stain mouse tissues. The antibody to MyoD immunoprecipitates a 45-kD protein, shifts the MyoD band in gel retardation assays of C2 nuclear extracts (Yablonka–Reuveni and Rivera 1994,1997), and does not crossreact with myogenin or myf5 in mass cultures from MyoD knockout mice (Yablonka–Reuveni and Rudnicki, work in progress) or in double staining experiments with myogenin monoclonal antibodies on intact fiber cultures (Yablonka–Reuveni and Rivera 1997).
The sections were warmed to room temperature, air-dried, and blocked overnight with sterile Tris-buffered saline (TBS) containing normal goat serum [0.05 M Tris, 0.15 M NaCl, 1% normal goat serum (NGS), pH 7.4]. After a thorough rinse (three times) with TBS containing Tween-20 (0.05%), sections were incubated with the primary antibody against MyoD for 1 hr at room temperature, followed by overnight incubation in a humid chamber at 4C. After rinsing in TBS-Tween-20 (three times), sections were incubated for 1–2 hr with fluorescein-conjugated donkey anti-rabbit IgG (Jackson ImmunoResearch Laboratories; West Grove, PA) diluted 1:75 in TBS–NGS, rinsed in TBS–Tween 20, and mounted in Vectashield mounting medium (Vector Laboratories; Burlingame, CA). Sections processed without primary antibody and those processed with nonrelated rabbit IgGs served as procedural controls and were run for each staining procedure on each type of tissue (hyperthyroid, euthyroid, hypothyroid, and normal and mdx muscle). These controls resulted in the absence of nuclear stain in intact and regenerating muscles. At least three to six sections from different animals of each group were each stained for MyoD in repeat experiments to confirm findings. Sections of crush-injured muscle were viewed in the regenerating region of the muscle (identified using phase-contrast optics), which was located midway between the crush site and surviving muscle as systematically defined in a previous report (McIntosh et al. 1994), to ensure that observations were representative of the previous findings for each treatment group. All sections were viewed on a Zeiss photomicroscope equipped with epifluorescence and phase contrast optics without knowledge of their source, and were photographed through the X20 objective for an overview of the regenerating tissue, using Kodak EL 135 (400 ASA) color film.
Results
The detailed characterization of muscle regenerating in normal C57 and mdx mice under low (hypothyroid), standard (euthyroid), and high T3 (hyperthyroid) conditions appears elsewhere (McIntosh et al. 1994; McIntosh and Anderson 1995; Pernitsky et al. 1996). Those previous reports include examination of the density of myotubes formed after 4 days of muscle regeneration, the proportions of new myotube nuclei that as myoblasts had proliferated and fused with myotubes between 3 and 4 days of recovery, and the growth and differentiation (expression of developmental myosin heavy chain) of new myotubes. Likewise, detailed descriptions of T3 effects on mdx mouse dystrophy are reported (as above; and Anderson et al. 1994). These earlier studies showed that the differences between the treatment groups can be seen at 4 days post injury. On the basis of the previous work, the present investigation was made under the same conditions, and focused on the 4 day post-injury time point using sections from the same muscles analyzed in the earlier studies as a representative profile of early regeneration and myotube formation.
MyoD Immunolocalization in Regenerating Normal (C57) Muscle
Intact fibers adjacent to the crush site (and fibers in normal contralateral muscles) showed no staining of nuclei. In euthyroid muscles after 4 days of repair, infrequent mononucleated cells contained MyoD+ nuclei, but many other mononuclear cells did not (Figure 1A). A low level of nonspecific background staining was present in the cytoplasm of cells in the crushed region of muscles. This type of nonspecific staining was seen by us using other antibodies as well (Anderson et al. 1991,1995). Sections in which the primary antibody was omitted from the staining protocol showed low background fluorescence after processing that included the careful blocking and washing steps reported above (not shown).
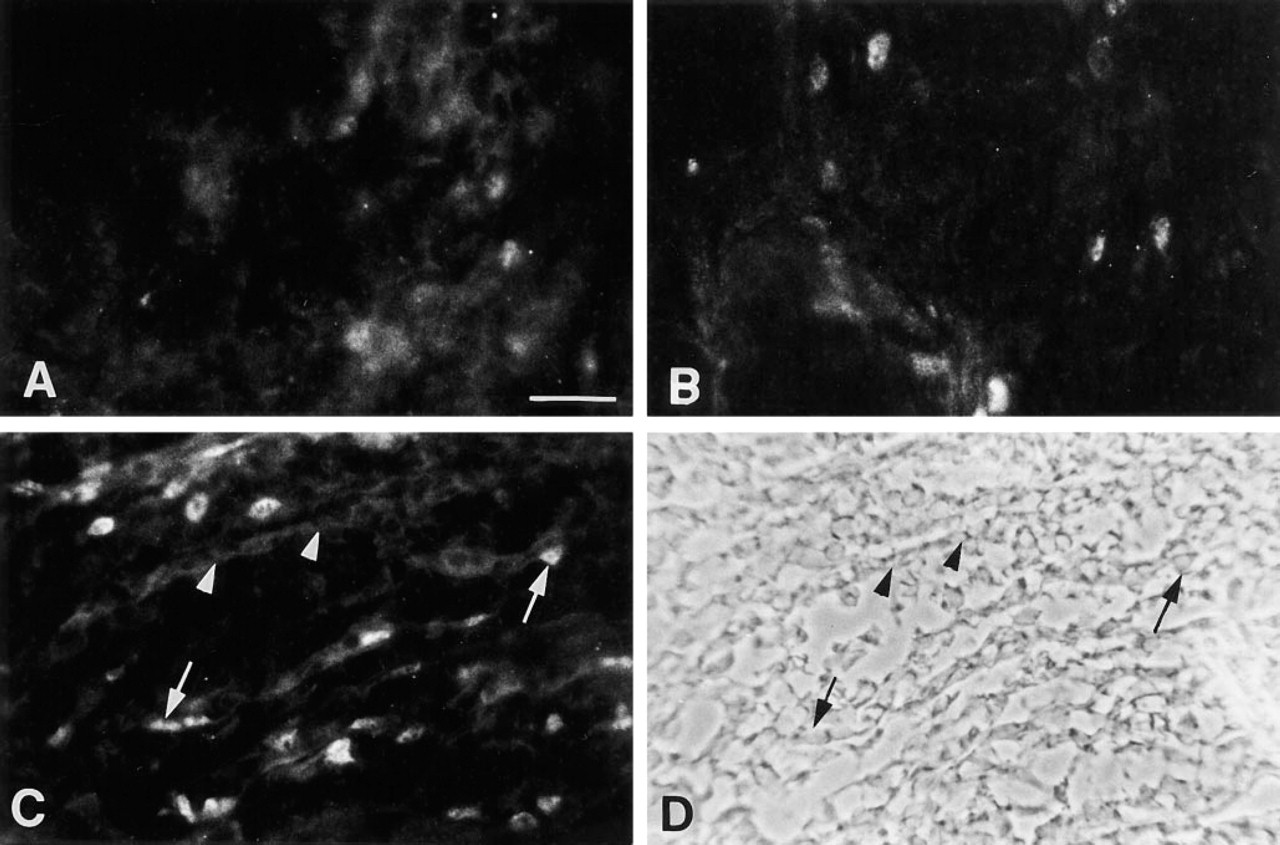
Immunofluorescence microscopy localization of MyoD protein in normal regenerating muscle. Sections were stained with a rabbit polyclonal antibody against MyoD, followed by fluorescein-conjugated donkey anti-rabbit IgG. The sections are from (
In hypothyroid mice, staining for MyoD protein and the relative number of MyoD+ nuclei in mononuclear cells per field of mononuclear cells were similar in muscle regenerating between the crush site and the surviving muscle compared to the areas of regenerating muscle in euthyroid mice (Figure 1B). Under hyperthyroid conditions in similar fields of regenerating muscle, we identified approximately twice as many mononuclear cells per field that were positive for MyoD in their nuclei compared to the number per field observed in euthyroid and hypothyroid regenerating muscles. As shown in Figure 1C, many of those MyoD+ nuclei were within the mononuclear cells that were aligned longitudinally before fusion into myotubes, as characterized using phase-contrast and comparison with other reports using sections stained with hematoxylin and eosin or Gomori's trichrome stain. Newly formed myotubes in the same region (identification confirmed using phase-contrast; Figure 1D) did not contain MyoD+ nuclei in normal muscles regenerating under hyperthyroid conditions.
MyoD Immunolocalization in Regenerating mdx Muscle
Overall, the number of MyoD+ nuclei in mononuclear cells per field was greater (roughly twofold) in all mdx treatment groups compared to the relative density of MyoD+ mononuclear cell nuclei that were observed in normal regenerating muscles. In euthyroid mdx mice, regenerating muscles had MyoD+ nuclei in mononuclear cells and very new myotubes (Figure 2A). As in normal muscle, there was background staining of the extracellular matrix surrounding intact fibers. The mdx hypothyroid group of regenerating muscles also demonstrated MyoD+ nuclei in mononuclear cells. In two sections of one muscle, MyoD+ nuclei were observed in myotubes and cells aligned longitudinally, although not fused into myotubes (Figure 2B). Finally, in the hyperthyroid mdx group, the relative density of mononuclear cells with MyoD+ nuclei was similar to that observed in regenerating muscles in euthyroid and hypothyroid mdx mice, some of which were noted to be very intensely stained (Figures 2C and 2D). In addition, intense nuclear staining for MyoD protein was observed in approximately 15% of the newly formed myotubes identified by phase-contrast (Figures 2E and 2F).
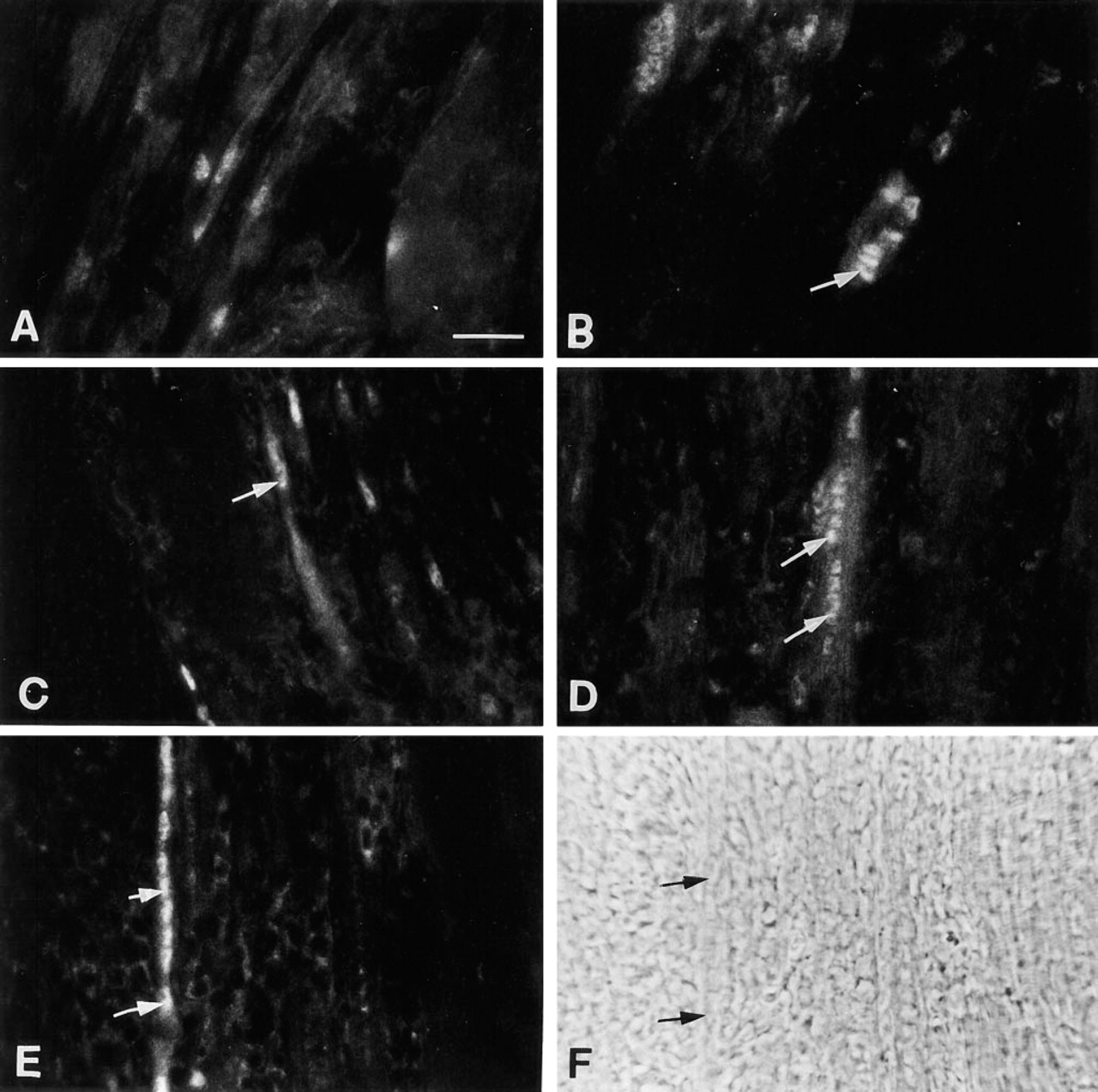
Immunofluorescence microscopy localization of MyoD in mdx regenerating muscle. The sections are from (
In intact fibers away from the crush injury (and in contralateral mdx muscles; not shown), there was no nuclear stain on myofibers that contained only peripheral nuclei (Figure 3A, hyperthyroid mdx group). However, MyoD+ nuclei were consistently observed in the very large myotubes (Figure 3B, hyperthyroid mdx group). These large myotubes (which contained central nuclei) were formed after earlier dystrophic damage, as characterized by other studies, and not as a result of the imposed crush injury. Nuclei in those large myotubes were less intensely stained relative to nuclei in the new myotubes formed since the crush injury. Undamaged fibers in regions closer to the regenerating area had occasional positive nuclei in their periphery. Examples are shown for surviving fibers in untreated mdx muscles (Figure 3C) and in older hyperthyroid fibers after regeneration (Figure 3B). By their location, these are nuclei in the satellite cells (i.e., cells that contribute myogenic precursors during growth and repair and are known to be located adjacent to myofibers).
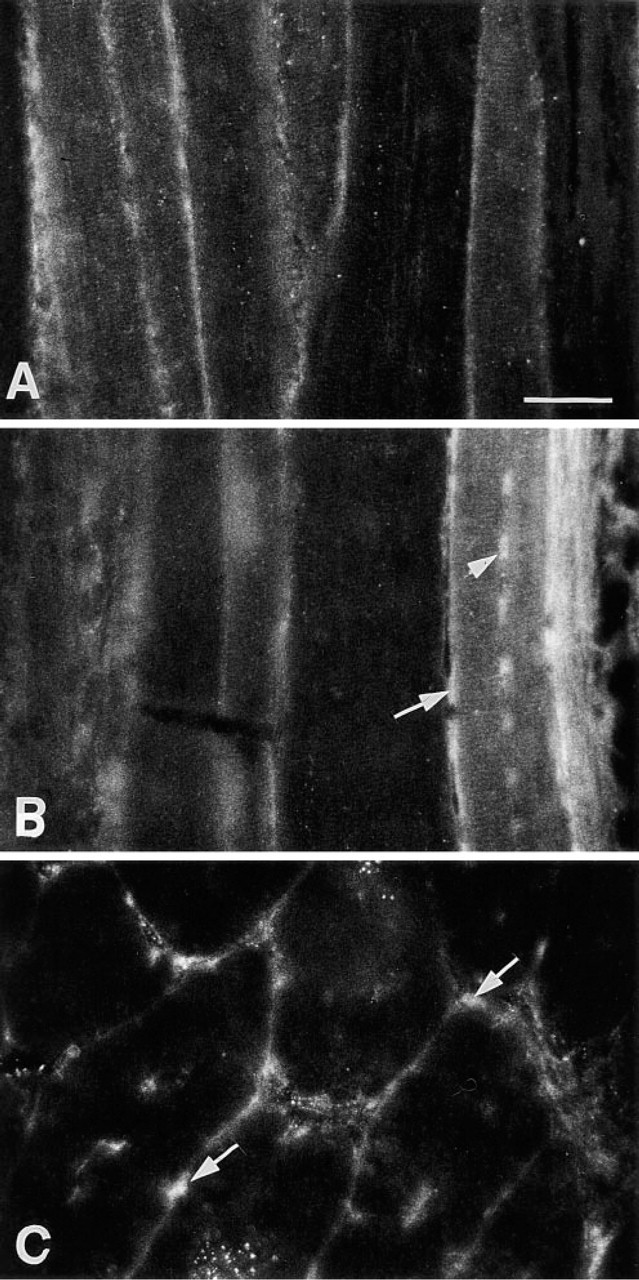
Immunofluroescence microscopy localization of MyoD in intact mdx muscle. Sections are from (
Discussion
In this study we have utilized immunostaining for MyoD to identify relative numbers of myogenic precursor cells at one time point during regeneration of normal and mdx muscle. With this approach, the influence of various levels of T3 was analyzed on representative regions of regenerating muscle containing mononuclear cells and new myotubes. Approximately two times more MyoD+ mononuclear cells per field were observed in mdx than in control regenerating muscles, and the relative density of MyoD+ nuclei in mononuclear cells in normal regenerating muscles increased under hyperthyroid conditions, in agreement with our hypothesis. However, we also noted MyoD+ myonuclei in well-differentiated large myotubes in mdx muscle regenerating from previous dystrophic damage, and in satellite cell nuclei next to fiber segments that survived the crush injury. Perhaps the most striking observation was that only repair in mdx muscles under high T3 conditions resulted in newly formed myotubes with MyoD+ nuclei. The relative number of MyoD+ mononuclear cells did not increase in mdx muscles regenerating in hyperthyroid compared to euthyroid conditions. These latter findings indicate that the pattern of differentiation is distinct in muscles regenerating in the two different strains, because MyoD+ nuclei persist in mdx but not in control myotubes regenerating from both dystrophy and crush injury during T3 treatment. The changes in normal muscle regeneration with various T3 conditions and the absence of relative changes in myoblast populations in mdx regenerating muscles are consistent with previous reports from the same experiments and also with detailed in vitro studies of myoblast proliferation and differentiation (Pernitsky and Anderson 1996; Pernitsky et al., submitted for publication). However, because these experiments examine only a single time point in regeneration, follow up studies later in regeneration are required to determine the length of MyoD expression in myotube nuclei under different T3 conditions, since that expression may relate to the functional endpoint of the regeneration process.
The changes in MyoD staining are consistent with observed changes in the repair process, determined through morphometric studies of myotube density and using autoradiographic tracing of myoblast fusion into myotubes (McIntosh et al. 1994; McIntosh and Anderson 1995; Pernitsky et al. 1996). The present work examined the relative density of MyoD+ cells in representative fields of muscle regenerating between the site of crush injury and the surviving muscle, rather than quantifying the exact numbers of myogenic precursors for a statistical analysis of MyoD expression. Previous reports have already measured the details of myoblast proliferation, myoblast fusion into myotubes during regeneration, and the density of myotubes formed under various T3 conditions (McIntosh et al. 1994; McIntosh and Anderson 1995; Pernitsky et al. 1996). At the time of the present studies, information on c-met receptor localization in both quiescent satellite cells and activated, proliferating myoblasts in vivo was not known (Tatsumi et al., submitted for publication). Myoblasts in the same muscles regenerating under euthyroid and high T3 conditions of normal and dystrophic mice were identified by staining with antibodies to developmental myosin heavy chain (Pernitsky et al. 1996), and can also be stained with antibodies to α-smooth muscle actin during or after MyoD expression in myogenesis on intact fibers (Yablonka–Reuveni and Rivera 1994), and by in situ hybridization with or without tandem immunostaining (e.g., Grounds et al. 1992; Garrett and Anderson 1995). Given those limitations, the semiquantitative changes in the relative level of MyoD expression in myoblasts per field were very consistent in muscles of each treatment group, because observations were made in the region between surviving fibers and the necrotic crush site in each muscle as described earlier (McIntosh et al. 1994). Together with previous findings on regeneration in mdx muscles and the influences of high and low levels of T3, the present MyoD immunolocalization study provides further understanding of muscle repair at the molecular level to supplement information from available quantitative studies from the same experiments.
MyoD+ mononuclear cells located in the satellite position near regions of muscle injury are probably activated satellite cells, as seen in cultures of intact fibers (Yablonka–Reuveni and Rivera 1994,1997) or elongating muscle precursors about to fuse with mature surviving fibers in regenerating muscles (Pernitsky et al. 1996). The location of MyoD in mononuclear cells and myotubes observed in this study, agrees with a previous report on regeneration of mouse TA muscle in the bupivacaine-treated autograft model of repair. That study showed that MyoD+ nuclei were present in myoblasts before fusion and in newly formed myotubes for 2 weeks post grafting. That 2-week interval in autograft regeneration was longer than observed from crush injury studies, because the former regenerative process is more prolonged and involves the entire muscle including the neurovascular supply (Fücht-bauer and Westphal 1992). In another study in rat tibialis anterior muscles regenerating after a freeze-injury, MyoD protein was observed in mononuclear cells. MyoD protein was only detected in nuclei of newly formed myotubes when repair occurred under denervation (or tenotomy) conditions (Koishi et al. 1995). Because downregulation of MyoD expression by electrical activity has been reported (Eftimie et al. 1991), the persistence of MyoD expression in a subset of myotubes formed in regenerating mdx but not in control muscles suggests that the innervation of those myotubes may be delayed by T3 treatment. However, the persistent MyoD protein in myotube nuclei in mdx regenerating muscles in this study was not accompanied by decreased myotube diameter (Pernitsky et al. 1996), so delays in reinnervation are unlikely to account for the prolonged MyoD expression in mdx myotube nuclei under hyperthyroid conditions.
Although exact comparisons are difficult because of the different models of injury-induced repair, it is possible that species and strain differences can explain the different results of these MyoD immunostaining reports, because only mdx but not control myotubes showed MyoD+ nuclei during regeneration under hyperthyroid conditions. Other strain differences have been noted in the response to T3 by myoblasts in vitro (Pertnitsky and Anderson 1996). Recent studies showed that the difference between normal and mdx myoblast reponses to T3 in vitro is partly explained by the ability of mdx myoblasts to continue cycling during MyoD expression. Normal control myoblasts, by comparison, respond to T3 by either differentiating and expressing MyoD protein or proliferating as MyoD-negative cells (Pernitsky and Anderson, submitted for publication). In addition, our preliminary observations (via immunolocalization) with primary myogenic cultures from rat and mouse muscles suggest that MyoD protein expression is a more consistent finding in myotube nuclei in mouse cultures compared to rat cultures. Many rat myotubes do not show detectable levels of MyoD, whereas mononuclear cells in rat and mouse cultures show similar stainability for MyoD. (Expression of myogenin protein is consistent in myotube nuclei in cultures from both species.) Although tissue culture studies show myotube nuclei that are positive for MyoD protein, the situation in vivo is much more complex and may provide a more stringent restriction on MyoD expression in myotubes. Perhaps T3 induces MyoD expression by myogenic precursors in mdx muscles to persist for longer during the repair process, and pushes that expression to continue past the stage when precursors fuse into myotubes. Alternatively, high levels of T3 may lead to a precocious fusion of myoblasts, not allowing sufficient time for the normal course of MyoD downregulation during myogenesis. Nevertheless, high T3 levels promote early differentiation of normal muscle during repair in vivo, as was previously reported (Pernitsky et al. 1996). However, because MyoD expression appears in both new and more mature (larger) myotubes formed by fusion under hyperthyroid conditions in mdx regenerating muscle, the effects of T3 are long-lasting. Perhaps distinctions between MRF expression in normal and mdx muscle fibers could help to account for the greater extent of repair in a given time period after injury of mdx than of normal muscles.
Our in vivo studies cannot hint as to whether T3 works directly on myoblasts or on an intermediate agent which in its turn affects myogenesis. However, different cell culture studies have shown that T3 directly affects myogenesis. T3 acts specifically to regulate MyoD (and myogenin) through thyroid response elements on those genes (Carnac et al. 1992; Downes et al. 1993; Muscat et al. 1993,1994,1995a,1995b). Exposure to added T3 results in a decrease in the proportion of myoblasts in S phase (Pernitsky and Anderson 1996). Additional tissue culture studies suggest that T3 may accelerate recruitment and the expression of MyoD, because cultures under low T3 conditions show a delay in myogenic differentiation (Carnac et al. 1992). The present results do not indicate whether T3 acts on cells that are already expressing MyoD or on myoblasts that have not yet entered the phase of MyoD expression (i.e. “pre-MyoD” cells, as discussed by Yablonka–Reuveni and Rivera 1997). Studies involving intact fiber cultures, and a direct analysis of population dynamics and the transition from the pre-MyoD to the MyoD+ state to the post-MyoD state would help to address that question. We are presently devising models to address this issue.
Different cell culture and in vivo models have led to a variety of conclusions regarding the expression of MyoD in proliferating and differentiating myoblasts. During myogenesis of satellite cells on isolated fibers, MyoD expression is completely correlated with myoblast proliferation, and MyoD expression is downregulated on transition into the myogenin-expressing state. Those studies used the same MyoD antibody as in the present experiments, which did not crossreact to detect myogenin. In cell cultures of dispersed tissue, MyoD expression was less transient and cells expressing myogenin were often positive for MyoD as well (Yablonka–Reuveni and Rivera 1997). Other cell culture analyses showed that although MyoD may first be expressed in proliferating myoblasts, the presence of MyoD in its active state signals cell cycle withdrawal and terminal differentiation (Halevy et al. 1995). Our studies at the immunohistochemical level did not attempt to specifically analyze the proliferation vs differentiation state of the myogenic precursor cells, but rather used MyoD protein expression to identify relative numbers of myogenic precursor cells in regenerating muscle in vivo.
Clarification of MyoD expression under different conditions in vivo is a valuable addition to understanding key events of precursor proliferation, myotube formation, and differentiation in muscle repair. In the present immunohistochemical study, we report semiquantitative data on relative expression of MyoD by mononuclear cells that are consistent with previous morphometric and autoradiographic studies on the same tissues, in particular the relatively greater expression by mononuclear cells in mdx than in normal regenerating muscles. However, the appearance of MyoD+ nuclei in newly formed mdx myotubes after injury under high T3 conditions suggests that the regulation of myoblast fusion and myotube differentiation may be modified in mdx mice and may contribute towards the differential regeneration capacity of mdx muscle compared to normal muscle. The study represents an initial approach to the molecular analysis of the influence of T3 on myoblast commitment and myotube differentiation during in vivo regeneration.
Footnotes
Acknowledgements
Supported by grants to JEA from the Manitoba Health Research Council and the Paul H.T. Thorlakson Foundation, and grants to ZY–R from the Cooperative State Research Service–US Department of Agriculture (Agreement No. 93-37206-9301) and the National Institutes of Health (AR 39677 and AG 13798). Studentships were awarded from the Medical Research Council of Canada (LMM) and the Manitoba Health Research Council (ANM).
We are grateful for the technical assistance of Anthony Rivera and Roy Simpson.