
Abstract
Tight-skin (Tsk) is a dominant gene mutation that causes a fibrotic skin disease in mice, similar to human scleroderma. Both conditions are characterized by increased numbers of dermal fibroblasts containing high levels of procollagen mRNA. Whether this fibroblast population arises from fibroblast growth or fibroblast transcriptional activation is debated. Proliferation and apoptosis of fibroblasts of normal and Tsk mice were studied in skin sections before, at onset, and in established fibrosis. Tissue sections were immunostained with proliferating cell nuclear antigen (PCNA) as proliferation marker. Apoptosis was investigated by in situ end-labeling of fragmented DNA and nuclear staining with propidium iodide. The expression of the apoptosis inhibitor Bcl-2 was investigated by immuno-histochemistry. We demonstrate differences in fibroblast proliferation and apoptosis related to postnatal skin growth and development. Neonatal skin exhibits the highest levels of proliferation and apoptosis in fibroblasts. In contrast, low proliferation and absence of apoptosis characterizes adult fibroblasts. Skin fibroblasts express Bcl-2 only in newborns, and at other ages Bcl-2 was restricted to epithelial cells. Our results also suggest that neither increased fibroblast proliferation nor defective apoptosis accounts for the fibrotic phenotype of Tsk. Therefore, transcriptional activation of extracellular matrix genes appears more relevant in the pathogenesis of Tsk fibrosis.
H
The kinetics of fibroblast growth have been extensively studied in vitro (Bruce et al. 1986; Schneider and Mitsui 1976; Martin et al. 1970). Studies suggest that proliferation and apoptosis are linked and are developmentally regulated. In general, cultured young or c-myc-expressing fibroblasts, with greater proliferation rates, exhibit lesser resistance to apoptosis than senescent fibroblasts (Wang 1995; Evan et al. 1992). This may be related to the need of cell cycle progression for both proliferation and apoptosis (Hoang et al. 1994; Wu and Levine 1994). In addition, the Bcl-2 protein, an inhibitor of apoptosis, is differentially expressed in young and senescent fibroblasts and is involved in their different susceptibility to apoptosis (Wang 1995). Both proliferation and apoptosis might regulate the rate of fibroblast growth in fibrotic diseases. In vivo studies comparing these processes in scleroderma vs normal skin have not been published.
The tight skin mouse (Tsk) is the result of a mutation on the fibrillin-1 gene in chromosome 2 (Siracusa et al. 1996; Green et al. 1976). Penetrance is complete and Tsk mice develop skin fibrosis with a consistent temporal sequence. Our studies and those of others in Tsk mouse show that its molecular pathology is similar to human scleroderma (Pablos et al. 1995; Jimenez et al. 1986). Although Tsk is normal at birth, in the second week of life it develops skin tightness and increased numbers of high procollagen mRNA-containing fibroblasts in the skin, with a pattern indistinguishable from that of human scleroderma. A severe constraint to study of the fibrotic process in human scleroderma in vivo is the difficulty in obtaining early and sequential tissue samples with appropriate controls. Therefore, Tsk provides an interesting model to sequentially study fibroblasts behavior during the development of fibrosis.
In this article we describe the proliferative and apoptotic rates of skin fibroblasts through normal mouse postnatal skin development and Tsk skin fibrosis. Proliferation was evaluated by proliferating cell nuclear antigen (PCNA) staining. PCNA is expressed in proliferating fibroblasts at the G1/S-phase transition, and its expression in fibroblasts in vitro parallels their proliferative ability (Stewart and Dell'Orcco 1992; Chang et al. 1991). Therefore, it serves as a good indirect marker for fibroblast proliferation. We have also studied apoptosis and the expression of Bcl-2, an apoptosis inhibitor. Our observations suggest that proliferation and apoptosis of fibroblasts normally occur at variable rates during postnatal life. Differences in fibroblast growth between Tsk and normal mouse do not appear to be involved in the skin fibrosis of this animal model of scleroderma.
Materials and Methods
Animals
TSK (C57BL/6-Tsk+/+pa) and normal (C57BL/6-+pa/+pa) mice were originally obtained from the Jackson Laboratory (Bar Harbor, ME). A Tsk colony has been established and maintained at our animal facility by brother-sister mating of the C57BL/6-Tsk+/+pa by normal (C57BL/6-+pa/+pa) mice. Tsk and normal mice at <1 day, 16 days, and 1 year of age were sacrificed under a CO2 atmosphere. Interscapular skin was removed and portions snap-frozen (liquid N2) for DNA extraction, or fixed for 1 hr at 4C in 4% paraformaldehyde in PBS and paraffin-embedded. Several sections of three paired normal and Tsk littermates at each age were used for quantitative analysis.
Histological Methods
Skin samples from Tsk and normal littermates were cut at 6 μm and mounted on silanized slides. Slides were dewaxed in xylene, rehydrated through an ethanol series and finally, after staining, dehydrated and mounted in Permount medium. Nuclear morphology was evaluated by propidium iodide (PI) and conventional hematoxylin staining. PI staining was performed at 1 μg/ml for 15 min at room temperature (RT) after digestion with DNAse-free RNAse A, 200 μg/ml in PBS for 1 hr at 37C.
PCNA and Bcl-2 Immunohistochemistry
Skin sections were treated with 1.2% H2O2 in absolute methanol for 30 min and stained by the indirect avidinbiotin-horseradish peroxidase method (ABC standard; Vector Laboratories, Burlingame, CA). Color development with diaminobenzidine (Vector Laboratories) was monitored by appearance of the normal Bcl-2 and PCNA brown staining in normal epidermis. The primary antibody to PCNA (PC10; Boehringer Mannheim, Mannheim, Germany) was applied at 1:1000 concentration overnight at 4C. This was the optimum of the tested concentrations (1:100-1:2000) in preliminary experiments. Anti-Bcl-2 (N-19; Santa Cruz Biotechnology, Santa Cruz, CA) was a specific rabbit polyclonal and was applied at 1:500 dilution. Negative controls without primary antibody were included.
Sections were counterstained with hematoxylin for quantification of stained and non-stained fibroblasts. PCNA and Bcl-2 indexes were calculated as the number of positive fibroblasts divided by the total number of fibroblasts × 100, and were evaluated by counting at least 1000 fibroblasts by two independent observers (one of them blinded).
In Situ Detection of Apoptotic Cells
We performed the terminal transferase-mediated dUTP end-labeling technique (TUNEL) as described by Gavrieli, with minor modifications (Gavrieli et al. 1992). Sections were permeabilized with 0.5% Triton X-100 in PBS for 10 min at RT because in preliminary experiments, proteinase K treatment had produced a high background. The labeling mixture consisted of 30 mM Trizma base, pH 7.2, 140 mM sodium cacodyalate, 1 mM cobalt chloride, and 1 mM digoxigenin-12-dUTP, containing 0.3 U/μl of terminal deoxynucleotidyl transferase (TdT) (Boehringer Mannheim), and was applied to the sections for 30 min at 37C in a humid chamber. The reaction was terminated in 300 mM sodium chloride, 30 mM sodium citrate buffer for 30 min at RT. Sections were washed three times in PBS and developed with an alkaline phosphatase-labeled anti-digoxigenin antibody (Boehringer Mannheim) at 1:250 concentration for 1 hr at RT. The color reaction was developed with NBT/BCIP (BioRad Laboratories; Richmond, CA) and counterstained with methyl green (Vector Laboratories). Scoring was expressed as the absolute number of apoptotic fibroblasts per area at a magnification of X400.
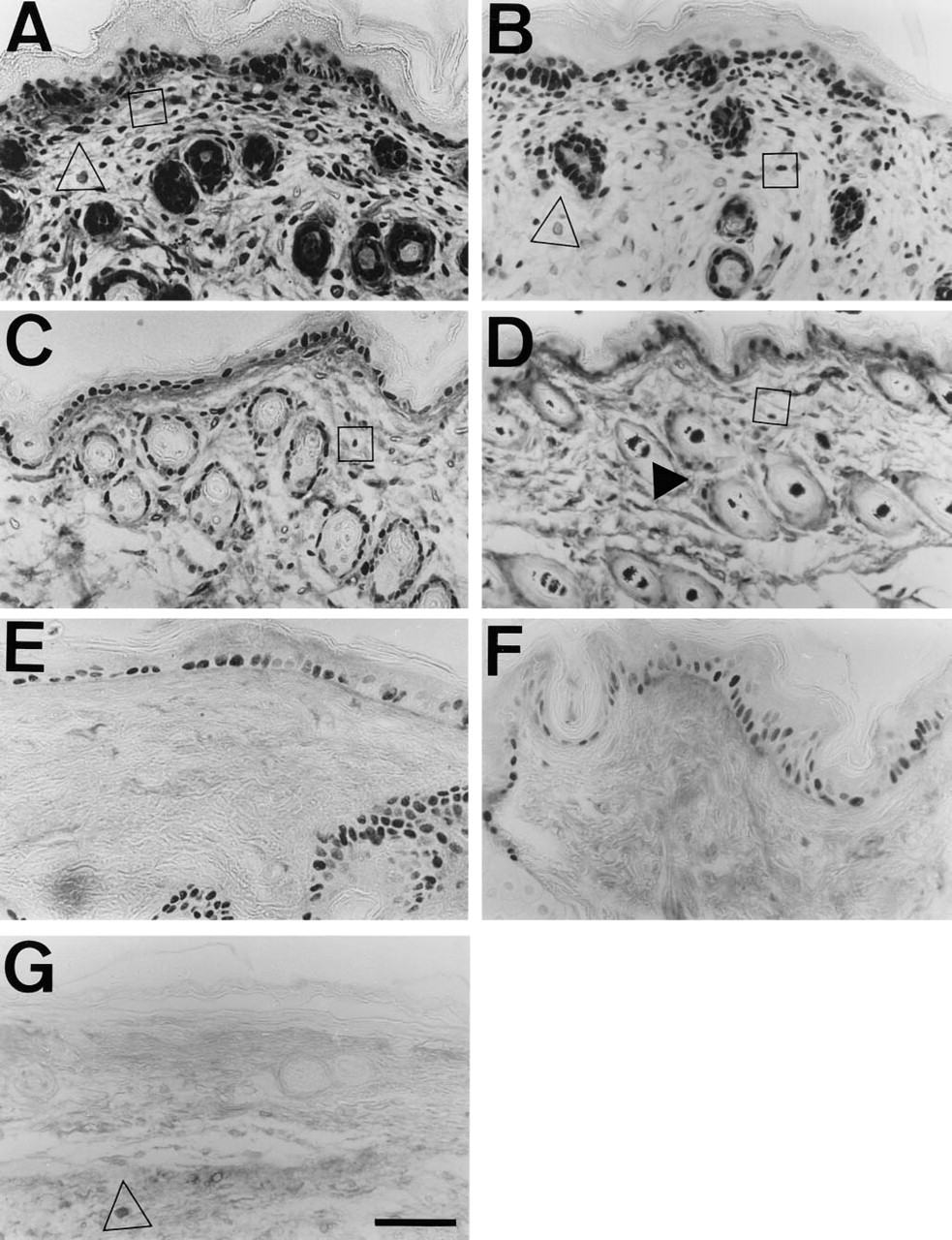
PCNA immunostaining of Tsk and normal mouse skin at different ages.
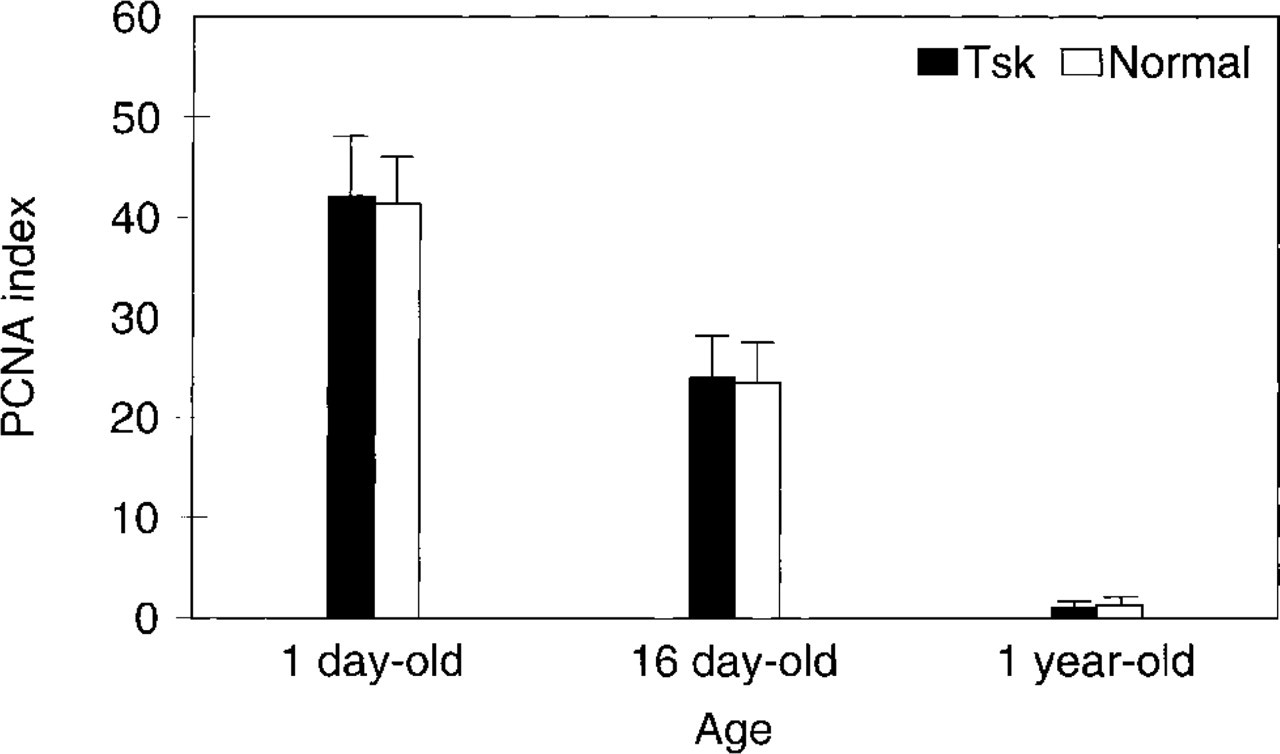
Proliferative index of skin fibroblasts in Tsk and normal mouse. PCNA index was maximal in the newborn and decreased with age. Mean was calculated in 1000 cells from different sections of three different animals at each age. Tsk and normal mean comparison failed to detect significant differences (newborn p=0.63; 16-day-old p=0.67; 1-year-old p=0.61).
DNA Extraction and Electrophoresis
Skin was ground under liquid N2 and digested with proteinase K (200 μg/ ml) (Boehringer Mannheim) in lysis buffer (10 mM Tris-HCl, pH 8, 10 mM EDTA, 0.1 M NaCl, 2% sodium dodecyl sulfate, and 39 mM dithiothreitol) at 56C overnight. The lysate was extracted with phenol/chloroform, ethanol-precipitated, and resuspended in 10 mM Tris-HCl, 1 mM EDTA, pH 8. DNA was treated with 50 μg/ml DNAse-free RNAse-A, re-extracted with phenol/chloroform, and ethanol-precipitated. DNA concentration was determined by spectrophotometry at 260 nm. Twenty μg of DNA was electrophoresed on 1.2% agarose gels in 1 X standard TAE buffer with DNA molecular weight standards (Gibco BRL; Gaithersburg, MD). Gels were stained with ethidium bromide and visualized under UV light.
Statistical Methods
Mean and SD from pooled data of different sections from three Tsk or normal animals at each age were calculated. Student's t-test was used to evaluate the statistical significance of differences between Tsk and normal mice. p values above 0.05 were considered not significant. Interobserver variability was found to be not significantly different.
Results
PCNA Staining
Both normal and Tsk skin at all ages studied revealed PCNA-stained nuclei in many epidermal keratinocytes, only at the basal and first suprabasal layers (Figure 1). This specific pattern was used as internal control for staining specificity in the evaluated sections, because the proliferative pattern of normal mouse skin fibroblasts was unknown. A high proportion of epithelial cell nuclei in hair follicles and glandular appendages was also stained. No differences in this pattern were observed between Tsk and normal mice. The proportion of labeled nuclei of epidermal and hair follicle cells (proliferative index) decreased slightly in older epidermis. Negative controls without primary anti-PCNA antibody did not show any nuclear staining, and only mast cells showed cytoplasmic staining due to nonspecific binding of avidin (Figure 1G).
Stained fibroblast nuclei were observed at all ages in both Tsk and normal mice. The spatial distribution was even in all skin layers. No accumulation of PCNA-stained fibroblasts in any particular area of dermis or hypodermis was detected. A high fibroblast proliferative index was found in newborn skin (<1 day old) (Figures 1A and 1B), decreased in the 16-day-old mouse (Figures 1C and 1D), and was very low in the 1-year-old mouse (Figures 1E and 1F). Tsk and normal mice displayed a similar fibroblast proliferative index at all stages (Figure 2). Statistically significant differences were not detected.
Apoptosis
Condensed chromatin, forming pyknotic or crescentshaped nuclei, and nuclear apoptotic bodies were scarce in dermal fibroblasts of newborn skin and absent in older samples by hematoxylin staining (Figure 3A) and PI staining (Figure 3B). These nuclei were mainly located in the dermis as isolated structures or small clusters. Newborn dermal fibroblasts were also labeled by TUNEL (Figures 3D, 4A, and 4B). TUNEL-stained slides were used for quantification. Quantification of TUNEL-stained fibroblasts nuclei as a proportion was difficult because the figures were under 1/1000 cells. Therefore, we obtained the data as the absolute number of stained nuclei per field at a magnification of X400. Comparison of means failed to show statistically significant differences between Tsk and normal skin (Tsk 0.8 ± 0.33, normal 0.7 ± 0.45 labeled nuclei per field; p=0.57). The TUNEL-negative controls, without TdT, did not show nuclear labeling (Figure 3C).
TUNEL-labeled nuclei appeared at all ages in epithelial cells of epidermis and hair follicles in much larger numbers than in fibroblasts (Figures 4A-4D). Interfollicular epidermal keratinocytes were preferentially labeled in the upper layers, and basal cells were not labeled. Labeling of hair follicle epithelial cells was minimal in hairless newborn skin and maximal at 16 days of age after emergence of hair. Nuclei were selectively labeled in the inner root layer of the medium third of hair follicles (Figures 4C and 4D). By DNA extraction and electrophoresis, we detected the DNA ladder diagnostic of apoptosis only in the 16-day-old skin samples (Figure 5). This coincides with a high number of hair follicle TUNEL-stained cells and a high density of hair follicles at this age.
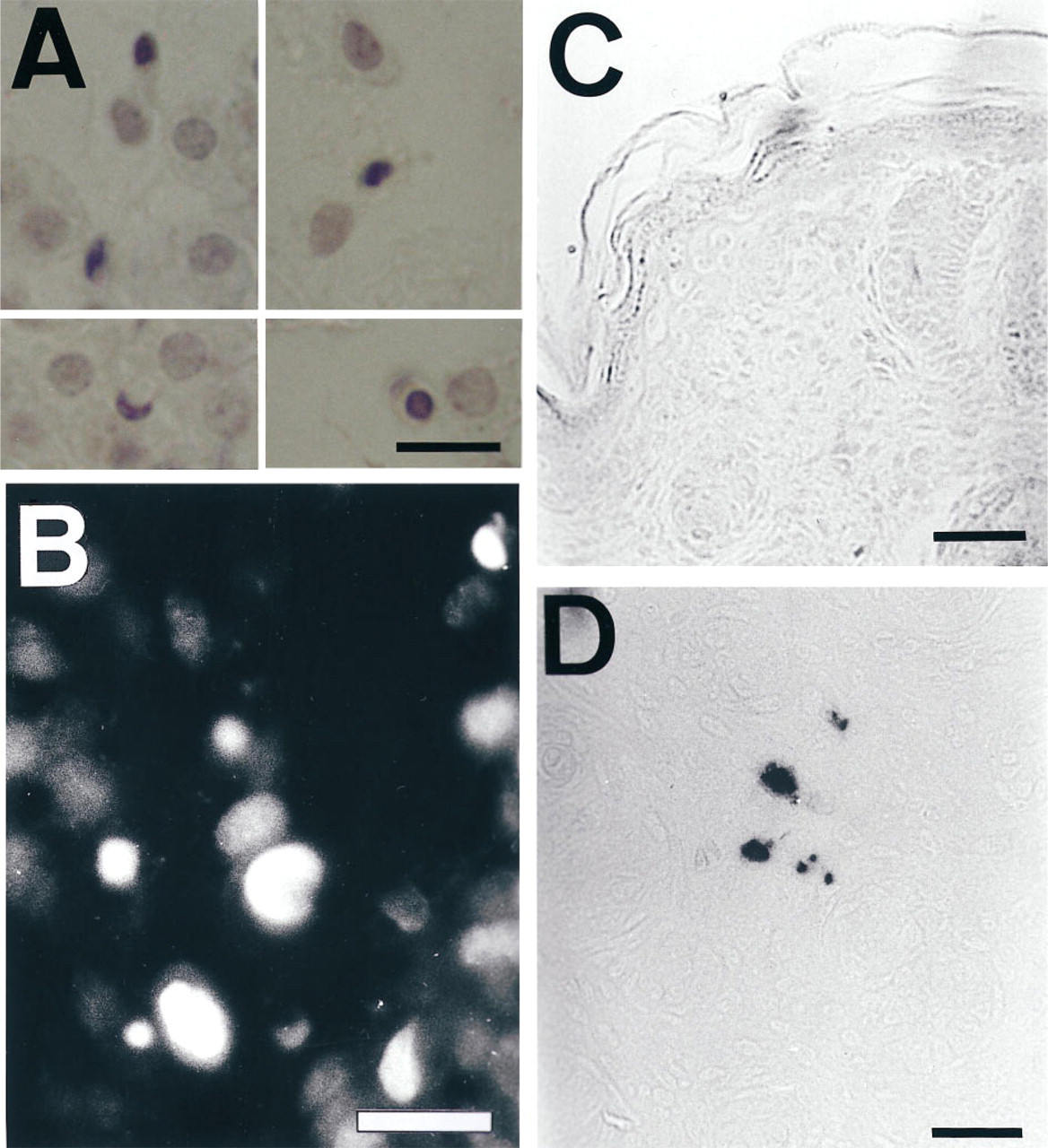
Apoptosis in dermal newborn mouse fibroblasts. Only in newborn skin are nuclei with condensed chromatin observed in dermal fibroblasts by hematoxylin
Bcl-2 Staining
Granular cytoplasmic Bcl-2 staining with mild perinuclear condensation in basal epidermal and appendageal epithelial cells was observed in Tsk and normal mice skin at all ages (Figure 4E). In newborn skin, cytoplasmic staining of many fibroblasts in the intermediate layers of the dermis was also seen (Figure 4F). The pattern was similar in Tsk and normal mouse skin. No statistically significant differences were detected between Tsk and normal mouse skin in the index of Bcl-2 labeled fibroblasts (normal 31 ± 6, Tsk 28 ± 7; p=0.31). In the 16-day-old (Figure 4E) and older normal or Tsk mice, there was no Bcl-2 staining in dermal fibroblasts or other connective tissue cells.
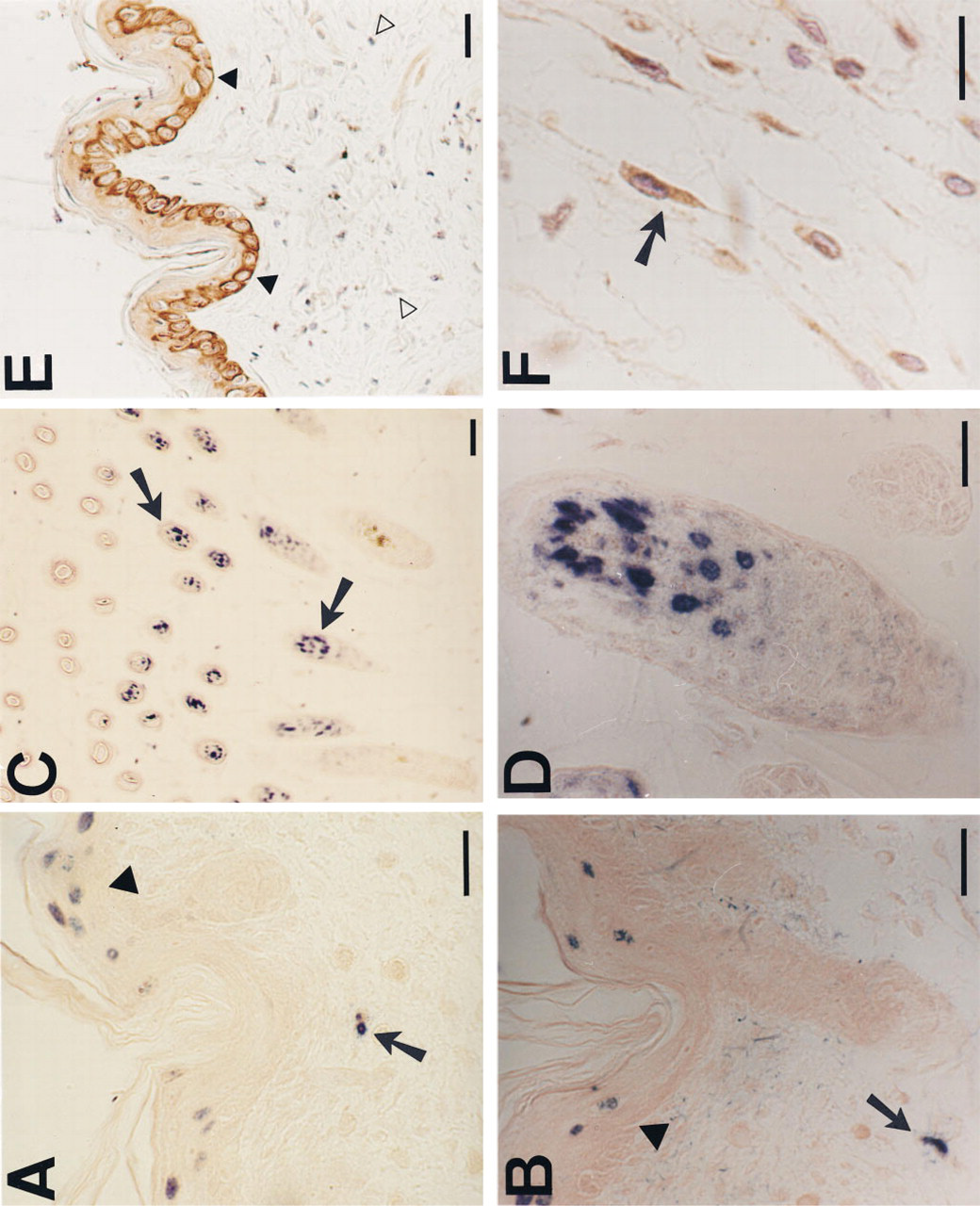
TUNEL and Bcl-2 staining of Tsk and normal mouse skin at different ages. Epidermal cells (arrowheads) and fibroblasts nuclei (arrows) are labeled by TUNEL in newborn normal
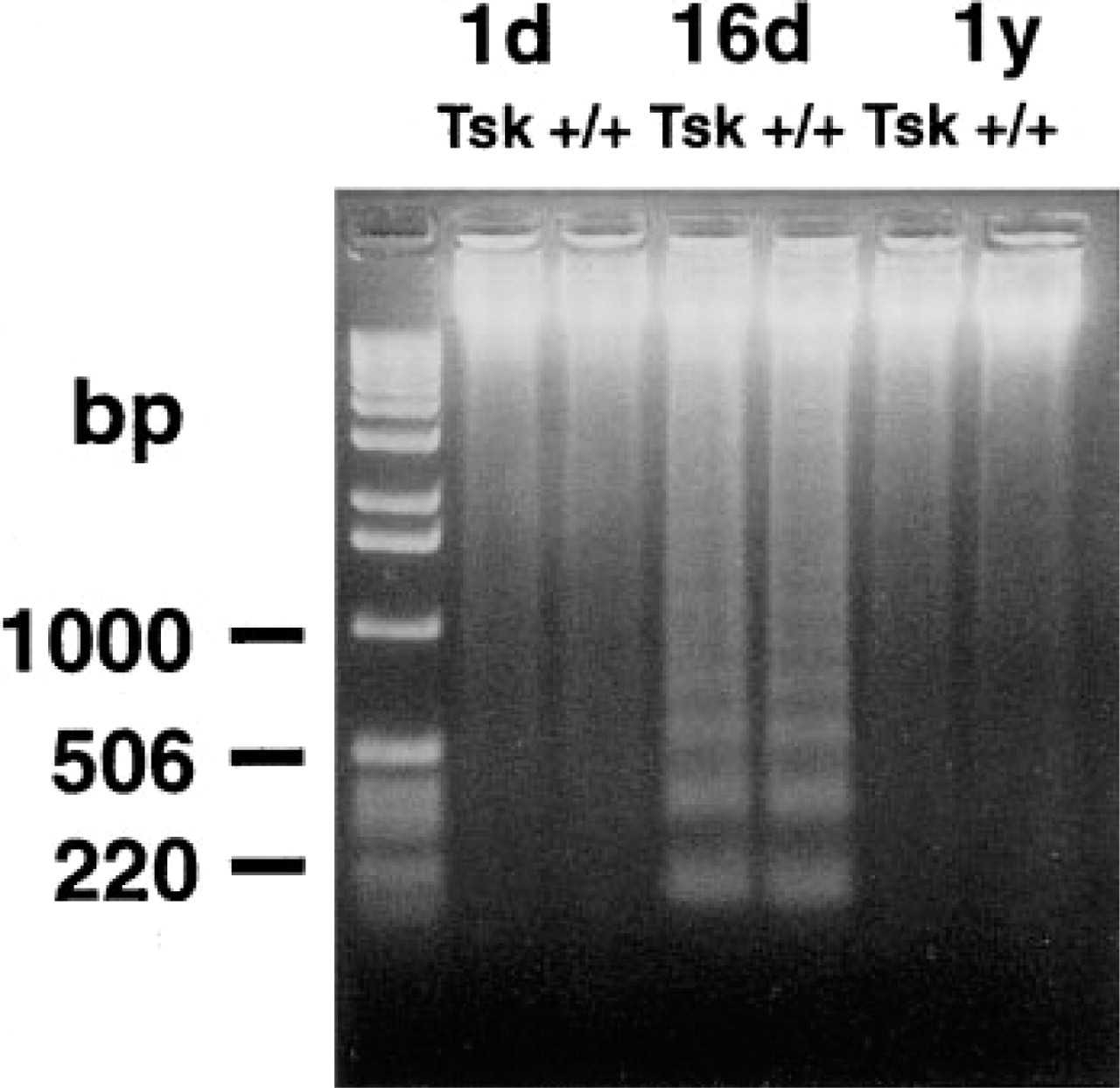
Electrophoresis of DNA from Tsk and normal skin. Ladder diagnostic of apoptosis is only observed in the 16-day-old mice, coinciding with a high number of TUNEL-positive cells in hair follicles at this age. DNA standard sizes are indicated at left.
Discussion
In vitro, fetal and neonatal skin fibroblasts have greater proliferative capacity than adult or aged samples (Bruce et al. 1986; Schneider and Mitsui 1976; Martin et al. 1970). In addition, fibroblasts in culture lose their proliferative capacity after a number of passages, and when senescence is achieved they are unable to proliferate. This occurs in parallel with a decrease in the expression of PCNA mediated by post-transcriptional mechanisms (Stewart and Dell'Orcco 1992; Chang et al. 1991). In vivo, our present work demonstrates a progressive reduction in the number of PCNA-expressing skin fibroblasts with aging, suggesting a parallel decrease in their proliferation rate in vivo similar to that observed in the in vitro aging models. Hence, this in vivo and in vitro analogy substantiates PCNA staining as a good marker for fibroblast proliferation.
In mouse, maximal proliferation was seen in the newborn and it decreased as early as the third postnatal week, when the dermis has reached its maximal thickness. This sequence closely resembles the rapid decrease in numbers of procollagen gene-expressing cells that we have previously reported within this time frame (Pablos et al. 1995). Therefore, it appears that a high rate of fibroblast proliferation is linked to high collagen gene expression during neonatal growth, and both are kept to a minimum in the adult mouse dermis. High levels of TGFβ mRNA are present in mouse dermal fibroblasts during the 2 postnatal weeks, and they became undetectable by in situ hybridization afterwards (Pablos et al. 1995). Although TGFβ is a potent inducer of procollagen gene transcription in fibroblasts (Ignotz et al. 1987; Varga et al. 1987), its in vivo effect on proliferation is less clear (Yamakage et al. 1992). In vitro, TGFβ is not mitogenic but enhances fibroblast susceptibility to other mitogens and induces expression of the fibroblast mitogen PDGF and its α-receptor (Ishikawa et al. 1990; Leof et al. 1986). Our present and previous observations in normal mouse dermis show that proliferation, matrix expression, and the presence of TGFβ message in the dermis follow a strikingly concordant pattern that suggests a causal link.
In human scleroderma, it is hypothesized that vascular or inflammatory cell mediators may selectively induce proliferation of the high collagen-producing fibroblasts present in normal dermis. We failed to detect differences in fibroblast proliferation between normal and Tsk fibrotic dermis. Nevertheless, Tsk skin fibrosis shows some differences from its human counterpart. First, it lacks the vascular and inflammatory lesions of human scleroderma (Jimenez 1988). Second, in human scleroderma there is an increase in the number of high collagen-producing fibroblasts compared with age-matched skin (Kahari et al. 1988; Scharffetter et al. 1988). In contrast, the onset of Tsk fibrosis coincides with a large number of high collagen-expressing fibroblasts in normal dermis, which is not properly reduced in Tsk thereafter (Pablos et al. 1995). Therefore, we investigated whether decreased apoptosis of the neonatal high collagen-producing fibroblasts in Tsk could be responsible for their persistence and the development of fibrosis. This does not appear to be the case, because apoptotic fibroblasts were found only in newborn dermis and at a similar density in Tsk and normal mice.
Apoptosis is related to high cell proliferation under many circumstances and has been regarded as a protective mechanism against the proliferation of partially damaged or wrongly programmed cells. Immortalized and primary embryo fibroblast cell lines with constitutive c-myc expression are susceptible to apoptosis in vitro (Evan et al. 1992). By contrast, fibroblasts transfected with c-myc mutants or in vitro aged are resistant to apoptosis (Wang 1995). In vivo, apoptosis of normal dermal fibroblasts has not been previously reported. In concordance with the in vitro studies, we detected apoptosis only in newborn skin fibroblasts, coinciding with their maximal proliferation rate.
One factor involved in the resistance to apoptosis of senescent fibroblasts is Bcl-2 (Wang 1995). In vitro, Bcl-2 is downregulated in young fibroblasts on serum deprivation, but it remains unchanged in older fibroblasts. We detected Bcl-2 immunostaining in the fibroblasts of normal and Tsk newborn skin but not in young or adult mouse fibroblasts, despite the staining of epithelial cells. In contrast to in vitro data, the expression of this factor does not appear to be involved in the lack of apoptotic cell death of adult fibroblasts in vivo. Bcl-2 has been previously detected in vivo in embryonic but not adult fibroblasts (Polakowska et al. 1994; Hockenbery et al. 1991). Therefore, newborn dermal fibroblasts appear similar to prenatal fibroblasts, yet their Bcl-2 expression is rapidly downregulated to the point that in the 2-week-old mouse it is no longer detectable. It might be speculated that this rapid downregulation contributes to apoptosis in newborn fibroblasts.
Although abnormal fibroblasts growth responses to different stimuli have been reported in human scleroderma (Yamakage et al 1992; Trojanowska et al. 1988; LeRoy et al. 1982), no consistent differences in their basal proliferation rates have been found. Our and other's unpublished work has also failed to show increased proliferation of Tsk fibroblasts in vitro, in keeping with the data reported here (Jimenez 1988). Recently, a novel transcriptional defect of Tsk fibroblasts in downregulating the activity of the collagen α1(I) gene promoter, caused by a decrease in the binding of a transcriptional inhibitor to AP-1 sites, has been described (Philips et al. 1995). This and the present observations point to the relevance of defects in transcriptional regulation of collagen expression, rather than to quantitative changes in the cellular kinetics of skin fibroblasts, as central to the skin fibrosis of Tsk. Further studies are needed to determine whether or not similar mechanisms operate in human scleroderma.
Acknowledgments
Supported in part by grants (94/0235 and 96/5254) from the Fondo de Investigaciones Sanitarias, Ministerio de Sanidad y Consumo, Spain.