
Abstract
The first step in the catabolism of branched-chain amino acids (BCAA), reversible transamination, is catalyzed by one of the two isoforms of branched-chain amino acid aminotransferase (BCAT). The mitochondrial isoenzyme (BCATm) is widely distributed among tissues, whereas the cytosolic isoenzyme (BCATc) is restricted to only a few organs. Remarkably, BCATc is the prominent isoenzyme found in brain. The physiological significance of the subcellular compartmentation of BCAT is still not understood. To contribute to the elucidation of the cellular distribution of the two isoenzymes in brain, we used cultured rat glial cells in an immunocytochemical study to determine the pattern of BCAT isoenzyme expression by glial cells. Antiserum against BCATm generated a punctate staining pattern of astroglial cells, confirming the mitochondrial location of this isoenzyme. In contrast, the cytosol of galactocerebroside-expressing oligodendroglial cells and O2A progenitor cells displayed intense staining only for BCATc. In addition, subpopulations of astroglial cells exhibited BCATc immunoreactivity. The presence of BCATm in astrocytes is consistent with the known ability of these cells to oxidize BCAA. Furthermore, our results on BCATc provide support for the hypothesis that BCATs are also involved in nitrogen transfer from astrocytes to neurons.
Keywords
T
BCAA, along with other large neutral amino acids, readily pass the blood brain barrier in exchange with brain glutamine (Cangiano et al. 1983). Therefore, it has been hypothesized that BCAA are involved in brain nitrogen metabolism and glutamine synthesis (Cooper and Plum 1987). Using cerebellar explants and cultured astroglial cells, Yudkoff et al. (1983,1994a) showed that [15N]-BCAA readily furnish amino nitrogen for the glial glutamate pool. Bixel and Hamprecht (1995) have shown recently that even when astrogliarich primary cultures are supplied with sufficient glucose, BCAA are metabolized rapidly by the cultured cells. Therefore, it has been proposed that BCAA may also serve as an energy source in brain.
Understanding the involvement of different brain cell types in BCAA metabolism, and the metabolic function(s) of the two BCAT isoenzymes in neural tissue, requires knowledge of the location of BCATm and BCATc in different cells within the brain. As a first step, we have investigated the cellular distribution of BCAT isoenzymes in rat glial cells, using pure astroglial and astroglia-rich rat brain primary cultures. Quite surprisingly, BCATm was found to be the predominant isoenzyme in astroglial cells. BCATc was found in astroglia, but at lower levels and with a heterogeneous distribution. However, strong staining of BCATc was found in oligodendroglial cells and O2A progenitor cells. The possible physiological implications for brain metabolism of this unique distribution of BCAT isoenzymes are discussed.
Materials and Methods
Materials
Dulbecco's modified Eagle's medium (DMEM) and fetal calf serum (FCS) were obtained from Gibco (Eggenstein, Germany). All cell culture plastic ware was from Nunc (Wiesbaden, Germany). Acrylamide, N,N'-methylene bisacrylamide, and sodium dodecyl sulfate (SDS) were from Bio-Rad Laboratories (München, Germany). Anti-glial fibrillary acidic protein (GFAP) monoclonal antibody, 5-bromo-4-chloro-3-indolylphosphate, 4-nitroblue tetrazolium chloride, and the molecular mass marker (Combothek) were from Boehringer Mannheim (Mannheim, Germany). Triton X-100 and Tween 20 were from Serva (Heidelberg, Germany). Nitrocellulose filter sheets were purchased from Millipore (Eschborn, Germany). Alkaline phosphatase-conjugated goat anti-rabbit IgG, fluorescein isothiocyanate (FITC)-labeled sheep anti-mouse IgG, and tetramethylrhodamine isothiocyanate (TRITC)-labeled goat anti-rabbit IgG were obtained from Sigma. The monoclonal antibody A2B5 was a gift from Dr. F. Walsh (London, UK). Monoclonal antibodies (MAbs) against galacto-cerebroside (GalC; Ranscht et al. 1982) were obtained as a gift from Dr. B. Ranscht and commercially from Boehringer Mannheim. The preparation of the antibodies against BCATc from rat brain (Hall et al. 1983) and BCATm from rat heart has been described (Wallin et al. 1990).
Cell Culture
Astroglia-rich primary cultures derived from the brains of newborn Wistar rats (bred in the animal facilities of the institute) were prepared and cultured as described previously (Hamprecht and Löffler 1985). Cells were seeded into plastic culture dishes 50 mm in diameter at a plating density of 2.5-3.0 × 106 viable cells per dish. The cells were incubated at 37C in a humidified atmosphere of 10% CO2/90% air in a nutrient medium containing 90% DMEM/10% FCS, 20 U/ml penicillin, and 20 μg/ml streptomycin sulfate. Every seventh day the medium was renewed using 90% DMEM/10% FCS. The vast majority of the cells in the cultures were astroglial cells (Reinhart et al. 1990). Pure astroglial primary cultures were prepared by the method of Verleysdonk and Hamprecht (1995). Rat glioma cells of the C6-BU-1 line were cultured as described previously (Amano et al. 1974).
Immunoblotting
The cultures were washed with PBS and scraped off the culture dish, homogenized in 100 mM Tris-HCl, (pH 7.4; 0.25 ml/mg protein) by 20 up-and-down strokes at 1400 rpm in a Potter-Elvehjem-type glass/teflon homogenizer. Blots were performed on proteins separated by SDS-PAGE electrophoresis (Burnette 1981). Transfer onto nitrocellulose was performed at a current of 0.3 A for 12 hr in transfer buffer (25 mM Tris, 192 mM glycine, adjusted to pH 9.0 with KOH). Nitrocellulose strips were treated with 1% (w/v) milk powder (from a grocery store) in incubation buffer [IB; 20 mM Tris-HCl, 150 mM NaCl, 0.02% (w/v) Tween 20, pH 7.4] at room temperature (RT) for 2 hr to block unspecific protein binding sites. After washing the strips three times with IB, they were treated with the antisera (diluted 1:500 in IB) at RT for 2 hr. Unbound antibodies were removed by washing three times with IB and the strips were incubated with secondary alkaline phosphatase-conjugated anti-rabbit antibody diluted 1:3000 in IB. After washing the strips with IB, color development was carried out in staining buffer (100 mM Tris-HCl, 50 mM NaCl, 5 mM MgCl2, 365 μM 5-bromo-4-chloro-3-indolyl-phosphate, 45 μM 4-nitroblue tetrazolium chloride, pH 8.9).
Immunocytochemistry
Cells grown to confluency on glass coverslips (22 X 22 mm) for 6-8 days after seeding were prepared for immunocytochemistry essentially as reported previously (Hamprecht and Löffler 1985; Hallermayer and Hamprecht 1984). For staining of cytosolic proteins, cells were fixed at RT with 4% paraformaldehye in PBS at pH 7.4 for 10 min. Fixed cells were washed twice in PBS for 5 min and once in PBS/0.1 M glycine for 5 min. Cells were permeabilized in PBS/0.3% (w/v) Triton X-100 for 10 min. Immunofluorescent double labeling was carried out using the indirect two-antibody method with FITC-labeled anti-mouse IgG for GFAP and TRITC-labeled anti-rabbit IgG for BCAT as secondary antibodies. Antibodies were diluted in PBS containing 0.3% Triton X-100 and 10% normal goat serum. The coverslips carrying the cells were then placed in a humidified chamber and treated with a mixture of the two primary antibodies for 2 hr and with the secondary antibodies for 1 hr. After each incubation, the cells were washed twice in PBS/0.1% Triton X-100. Labeling of BCAT together with GalC or A2B5 was performed stepwise. After washing the cells once with DMEM, they were immediately incubated at RT for 1 hr with anti-GalC or A2B5 antibodies (diluted with DMEM), respectively. Thereafter, the cells were washed twice with PBS, fixed, and permeabilized as described above, before they were incubated with BCAT antiserum for 2 hr and with the two secondary antibodies for 1 hr. coverslips were mounted cells down using 50% (v/v) glycerol in PBS. The preparations were viewed by glycerol immersion optics using a Zeiss fluorescence microscope (IM35) with a Plan-Neofluar X 25 objective.
Results
The presence of BCAT isoenzymes in homogenates of cultured rat brain cells was demonstrated by Western blot analysis (Figure 1), using an antiserum generated against BCATm from rat heart mitochondria (Wallin et al. 1990) and a BCATc antiserum obtained by immunization with BCATc purified from the cytosol of rat brain (Hall et al. 1993). The protein bands recognized by the two antisera were visualized by the insoluble reaction product generated by alkaline phosphatase coupled to the secondary antibody. In the Western blots, homogenates derived from rat astroglia-rich and pure astroglial primary cultures (Figure 1, Lanes 2 and 3) staining with BCATm antiserum resulted in a single band at approximately 41 kD (Figure 1, Lanes 6 and 7). BCATc antiserum in homogenates of rat astroglia-rich primary cultures recognized a protein with the molecular mass of approximately 47 kD, identifying BCATc (Figure 1, Lane 9). Each of these two isoenzymes could be derived from astroglial cells and/or other glial cells. Therefore, the experiment does not definitely answer the question of whether or not astroglial cells contain BCATm and/or BCATc. Therefore, homogenates of a pure astroglial culture were subjected to Western blot analysis. Again, single bands with the molecular masses of 47 kD and 41 kD were obtained (Figure 1, Lanes 4 and 10), demonstrating that BCATc and BCATm are indeed present in astroglial cells. C6 glioma cells and the C6-BU-1 glioma cells (Amano et al. 1974) that are derived from them display properties of both oligodendroglial and astroglial cells (van Calker and Hamprecht 1980). Homogenates of C6-BU-1 cells subjected to Western blot analysis were strongly immunoreactive for both BCAT isoenzymes (Figure 1, Lanes 5 and 8).
Western blot analysis established the presence of the two BCAT isoenzymes in the cultures. The next step was the cellular allocation of either isoenzyme in astroglia-rich primary cultures and pure astroglial cultures by examining the co-localization of the BCAT isoenzymes and cell type-specific markers. This was performed by using, in an indirect immunofluorescence double-labeling technique, antibodies against GFAP (Bignami et al. 1972), GalC (Raff et al. 1979), or tetrasialoganglioside (Kasai and Yu 1983), markers for astrocytes, oligodendrocytes, and O2A progenitor cells, respectively, in combination with antiserum against BCATc or BCATm.
In the double-labeling experiments shown in Figure 2, antisera against the BCAT isoenzymes were applied in combination with an antibody against the astroglial marker GFAP, using pure astroglial primary cultures. Staining for BCATm revealed co-localization of BCATm and GFAP in flat cells of irregular morphology (Figures 2A and 2B). Fluorescence corresponding to BCATm appeared in a distinctive punctate pattern in the cytoplasm of the cell somata, whereas the nuclei were clearly unstained. This is compatible with a mitochondrial localization for the antigen. Comparison of the intracellular distribution of the BCATm staining (Figure 2A) with the corresponding cell shapes (Figures 2A and 2B) demonstrated that the stained mitochondria appeared to be concentrated unevenly in the region surrounding the nucleus. Because of the thickness of the cells, some of the fluorescent grains representing mitochondria were out of focus. Figures 2D and 2E show the results of the simultaneous application of antibodies against BCATc and GFAP on the same cultures. The fluorescence signal for BCATc was distributed throughout the entire cell. In the area of the nuclei, the staining showed a higher intensity. Furthermore, not all GFAP-positive cells displayed the same BCATc immunoreactivity.
In the experiments shown in Figures 3B and 3E, an anti-GalC MAb was used for characterization of oligodendroglial cells in astroglia-rich primary cultures in combination with antisera against each BCAT isoenzyme (Figure 3). The cells that stained intensively with anti-GalC MAb had small, round somata, from which extended several thin and long processes characteristic of oligodendroglia. This morphology can also be seen in the corresponding phase-contrast micrograph, which shows cells sitting on a confluent layer of flat astroglial cells (Figures 3C and 3F). With BCATm antiserum, virtually no staining was detected in the GalC-positive cells (Figure 3A). In contrast, the oligodendroglial cells were intensively and evenly stained for BCATc throughout the cell soma and the thin extended processes (Figure 3D).
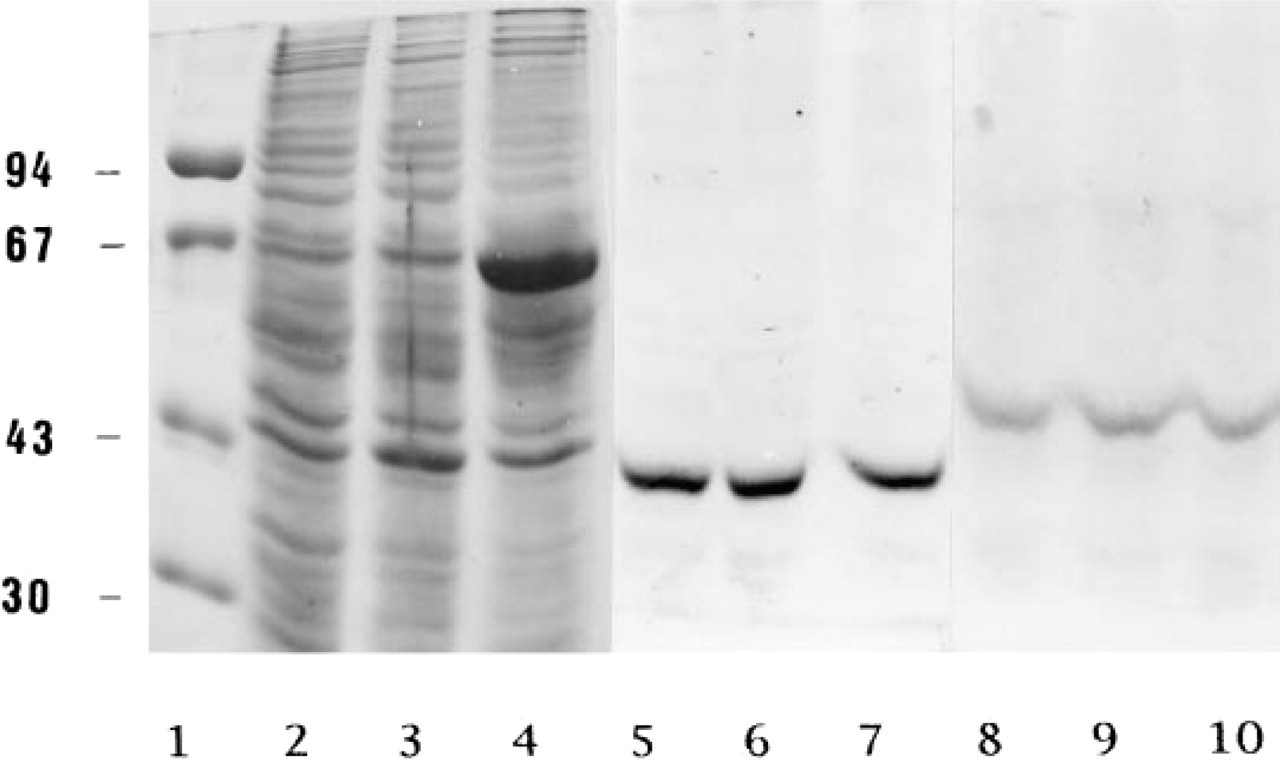
Western blot demonstrating the presence of BCATm and BCATc in glial cells in culture. The homogenates of several types of cultured brain cells were electrophoretically separated by 10% SDS-PAGE. After electroblotting onto nitrocellulose, Lanes 5-10 were immunostained by sequential incubation with respective antiserum, alkaline phosphatase-conjugated anti-rabbit IgG, and a solution containing nitroblue tetrazolium chloride and bromochloroindolyl-phosphate. Lanes 1-4, Coomassie Brilliant Blue staining (Laemmli 1970) of the molecular mass marker proteins (Lane 1, molecular mass in kD on the ordinate: phosphorylase 94 kD; bovine serum albumin 67 kD; ovalbumin 43 kD; carbonic anhydrase 30 kD) and of the protein (50 μg) in the homogenates derived from C6-BU-1 glioma cell cultures (Lane 2), astroglia-rich primary cultures (Lane 3), and pure astroglial cultures (Lane 4). Lanes 5-7 (8-10) illustrate immunostaining for BCATm (BCATc) using the homogenates of C6-BU-1 glioma cell cultures [Lane 5 (8)], astroglia-rich primary cultures, [Lane 6 (a)], and pure astrocyte cultures [Lane 7 (10)].
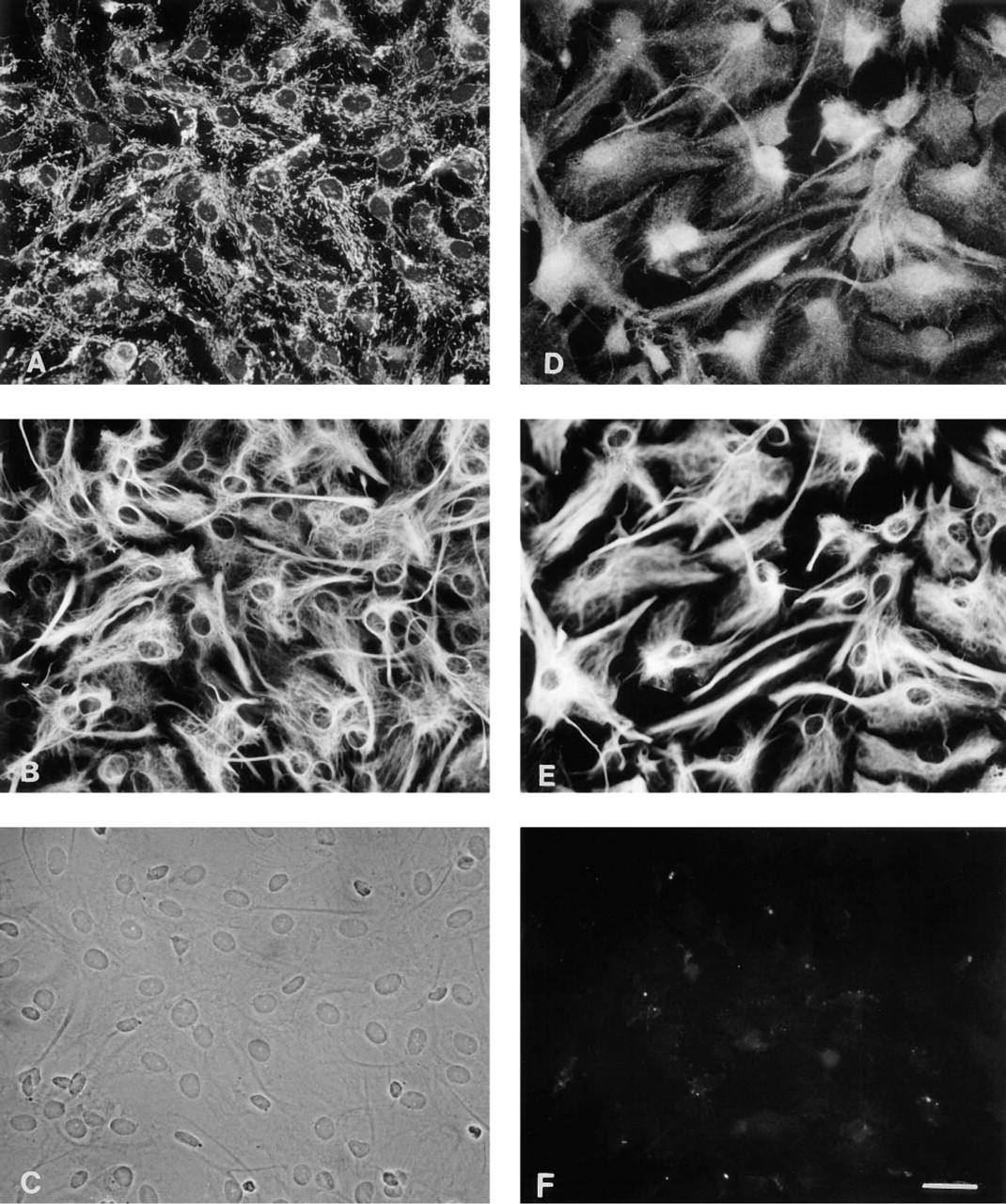
Immunocytochemical double-staining for the BCAT isoenzymes BCATm (
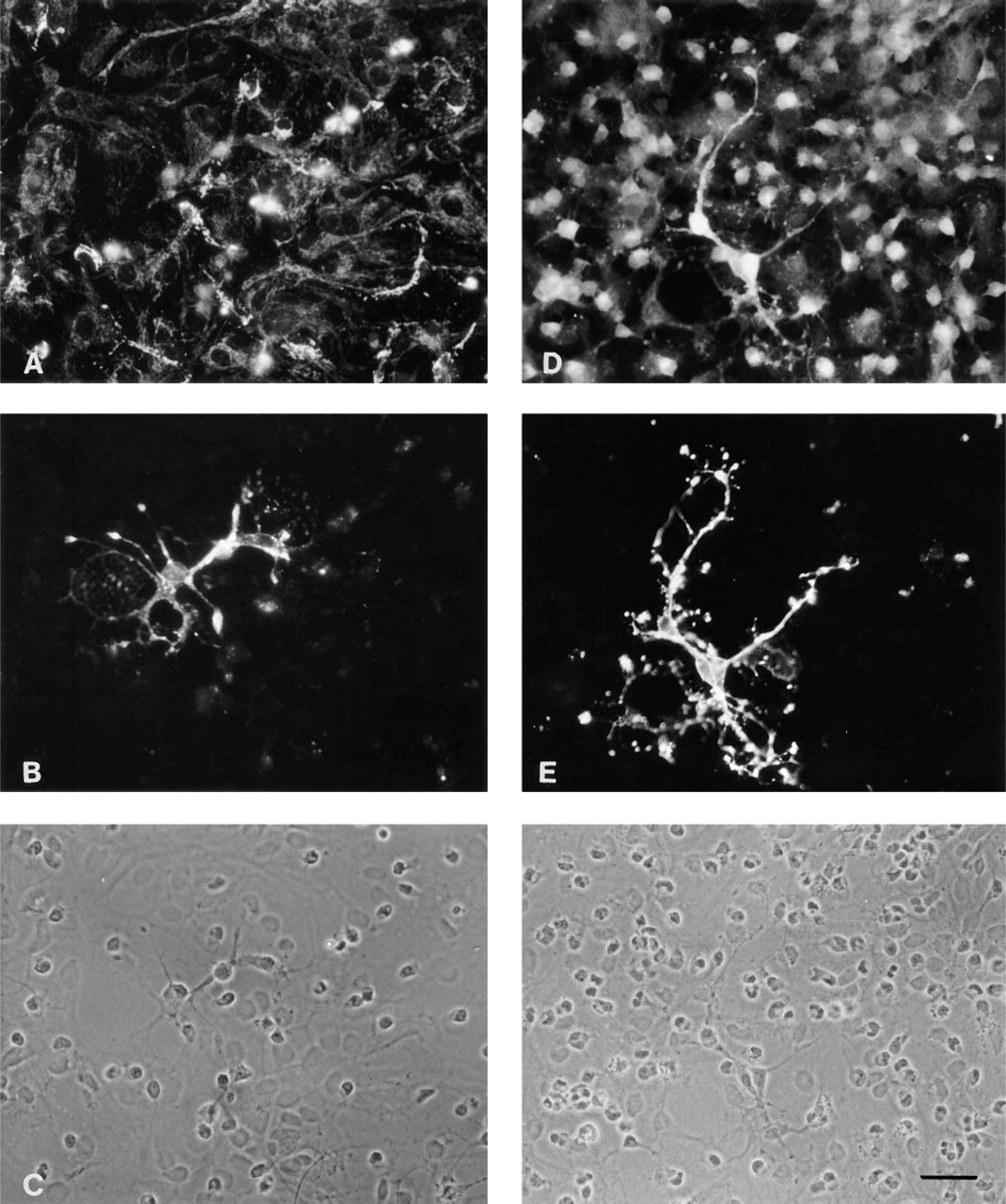
Immunocytochemical double-staining for the BCAT isoenzymes BCATm (
O2A lineage cells also were present in the mixed glial cultures. A tetrasialoganglioside specific for O2A progenitors, which could be stained with the antibody A2B5, allowed identification of the cells. Figures 4B and 4E show the stellate shape of typical O2A lineage cells, with spike-like projections extending from the small cell bodies. O2A lineage cells were only slightly immunoreactive for BCATm (Figure 4A), but their cytoplasm stained intensively for BCATc (Figures 4D-4F). On the other hand, not all BCATc-positive cells were apparently stained by the A2B5 antibodies (Figures 4D-4F, long arrows).
Discussion
The results of the present study suggest cellular localization of the BCAT isoenzymes in two types of glial cells in brain, astrocytes, and oligodendrocytes. The data show that in pure astroglial primary cultures the mitochondrial isoenzyme BCATm is located in astroglial cells. The mitochondrial location of the enzyme is confirmed by the punctate staining pattern in these cells. BCATm is the predominant BCAT isoenzyme outside of the CNS. It is a bifunctional protein that catalyzes the transamination of BCAA and the transport of the α-keto acid products (Hutson and Hall 1993). The presence of BCATm is consistent with the ability of astrocytes to degrade and oxidize BCAA (Bixel and Hamprecht 1995). Furthermore, the presence in the inner mitochondrial membrane of the transporter domain of BCATm suggests that the branched-chain α-keto acids (BCKA) generated by BCATm can be translocated into the cytosol and therefore are not necessarily available for oxidation by the mitochondrial branched-chain α-keto acid dehydrogenase. A second possible function of BCATm is transport of BCAK (Hutson and Hall 1993) generated by cytosolic transamination. This transamination could be catalized by BCATc of the same cell or of neighboring cells.
The fact that all GFAP-positive astroglial cells in culture appeared to stain strongly for BCATm may point to a constitutive expression of the gene of this isoenzyme, at least in cultured astroglial cells. In contrast, the staining for BCATc varied considerably, suggesting the existence of at least two populations of astroglial cells, one with low or moderate staining intensity and another population that did not stain for BCATc. This may indicate that expression of BCATc is a regulated astroglial function. The elevated staining by the BCATc antiserum of the nuclear area may be due to a thick layer of cytosol around the nucleus rather than a reflection of nuclear location of the isoenzyme. Oligodendroglial cells and O2A progenitor cells were also present in astroglia-rich cultures. In contrast to astroglial cells, oligodendroglial cells, identified by their specific surface marker GalC, contained BCATc as the only BCAT isoenzyme (Figure 3). The lack of tetrasialoganglioside (Figure 4) points to an advanced degree of maturity (Abney et al. 1983). The tetrasialoganglioside containing O2A progenitors, in addition to being considerably stained by BCATc antiserum, appeared to be slightly stained for BCATm. Furthermore, on the basis of the strong fluorescence in these cells, as analyzed by the indirect immunofluorescence technique, these cells contain BCATc in high concentrations.
The pattern of expression of BCAT isoenzymes raises some questions about the physiological significance of these enzymes in brain metabolism. The BCAA are believed to play an important role in brain nitrogen metabolism. It is documented that BCAA are readily taken up into brain via the blood-brain barrier (Cangiano et al. 1983), and Yudkoff et al. (1983, 1994b) have proposed that BCAA are the major external nitrogen source in brain for the formation of glutamate and glutamine. Astrocytes have a highly active glutamate uptake system (Westergaard et al. 1995; Hösli et al. 1986) and contain glutamine synthetase, which is preferentially located in this cell type (Martinez-Hernandez et al. 1977). In the glutamate/glutamine cycle (Berl et al. 1977; Hamberger et al. 1977; Benjamin and Quastel 1972), the neurotransmitter glutamate and, to a lesser extent, GABA (Van den Berg et al. 1978) released by neurons are removed from the extracellular space by astrocytes and converted to glutamine. Glutamine, which is then released by astrocytes, is taken up by neurons, where mitochondrial phosphate-dependent glutaminase (Kvamme et al. 1982; Weiler et al. 1979; Shank and Aprison 1977) catalyzes hydrolysis of glutamine to ammonia and neurotransmitter glutamate. In GABAergic neurons, glutamate regenerated from glutamine is further transformed to GABA by glutamate decarboxylase. According to a suggestion by Yudkoff et al. (1996), leucine could be the form in which this nitrogen is returned from neurons to the neighboring astrocytes. This hypothesis is based (a) on studies with [15N]-leucine (Yudkoff et al. 1983, 1994b), in which it was demonstrated that BCAA are the major external nitrogen source for the formation of glutamate and glutamine in astrocytes, (b) on the release of α-ketoisocaproate from leucine-fed astroglial cells into the culture medium (Yudkoff et al. 1995; Bixel and Hamprecht 1994), (c) the capability of synaptosomes to take up α-ketoisocaproate, and (d) on the observation that α-ketoisocaproate is transaminated to leucine in synaptosomes (Yudkoff et al. 1995). In addition, this hypothesis relies on the presence of glutamate dehydrogenase in neurons to fix the free ammonia and form glutamate that can be transaminated by BCAT (Rothe et al. 1994; Dennis and Clark 1977). The leucine generated from its corresponding α-ketoacid in the neuron could then cycle back to the astrocyte, where it would donate its amino group for the formation of glutamate, and subsequently of glutamine via glutamine synthetase. If glutamate dehydrogenase is active, the BCAA can also provide free ammonia for glutamine synthesis. This results in a counter-traffic of glutamine and α-ketoisocaproate from astrocytes to neurons and of glutamate and leucine in the opposite direction, assuming that BCAT activity is present not only in astroglial cells but also in neurons (Figure 5). Preliminary evidence for the presence of BCAT in neurons in culture (Bixel et al. 1995; and Bixel, Hutson, and Hamprecht, unpublished results) supports the concept of the neuronal fixation of ammonia in leucine.
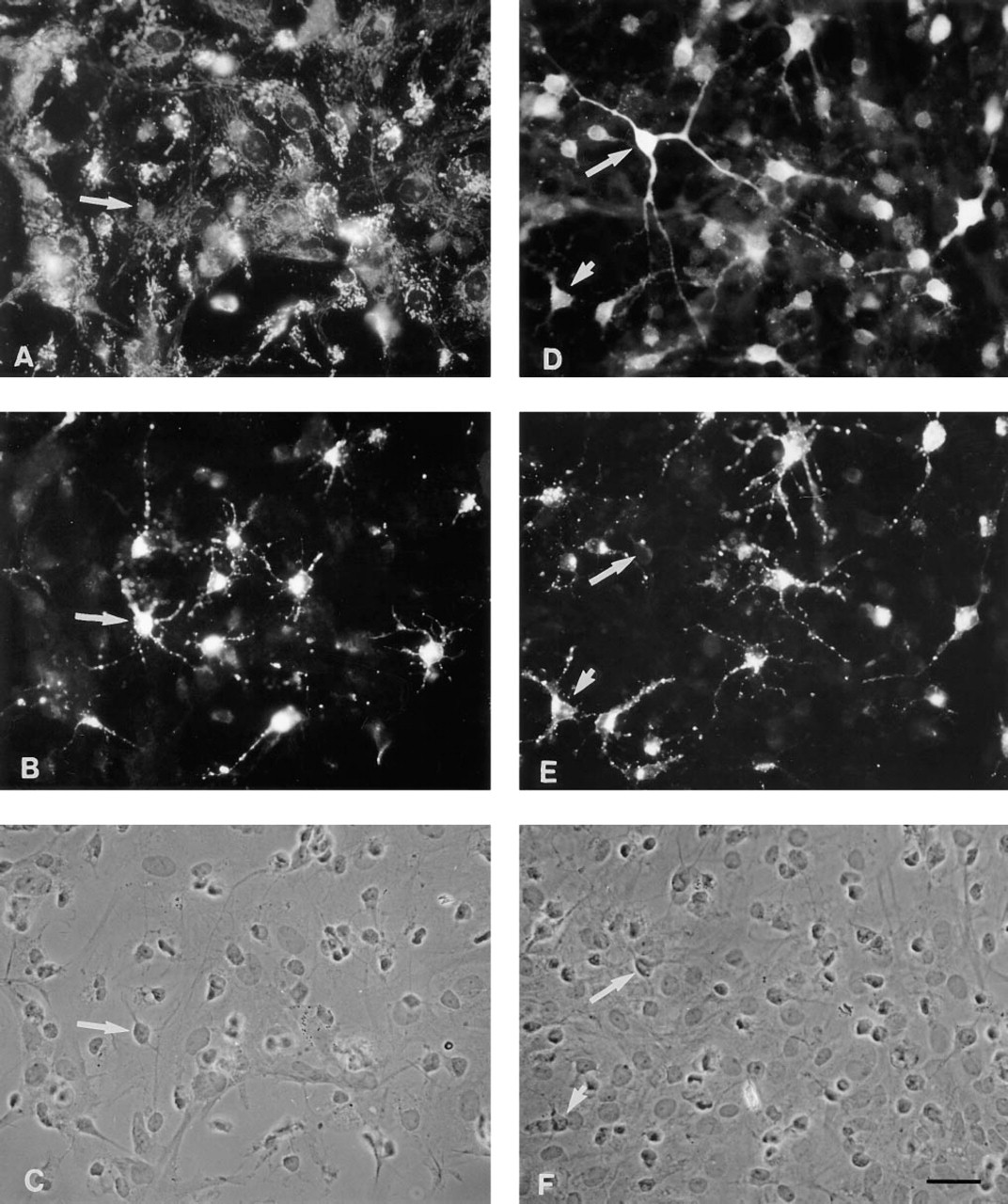
Immuncytochemical double-staining for the BCAT isoenzymes BCATm (
The pattern of expression of the BCAT isoenzymes in cultured astroglial and oligodendroglial cells, i.e., the variable co-existence of both isoenzymes in the former and the exclusive presence of BCATc in the latter cells, may not necessarily reflect the expression pattern in brain. In culture the cells are taken out of their normal cellular context, and cellular interactions occurring in brain by cell surface molecules or humoral factors that would be necessary for an expression to be induced or suppressed may be missing in culture. Because such factors may change during development, the expression of the BCAT isoenzymes in different brain cell types could vary with the stage of brain development and the region of the brain. Therefore, the cellular distribution of these isoenzymes must be studied in various brain areas as a function of the development from embryonic to adult animal life. Discrepancies between in vitro and in vivo functions may provide access to the factors regulating the expression of the BCAT isoenzymes.
The equilibrium constants for most aminotransferase reactions, including BCAT (Taylor et al. 1970), are close to unity and the BCAT isoenzymes can catalyze reversible transfer of amino groups, which is known to occur in situ (Hutson et al. 1978) and in vivo (Harper et al. 1984). The relative rates of deamination of BCAA vs reamination to BCAA depend on the concentrations of BCAA, BCKA, glutamate, and α-ketoglutarate in the cell, as well as on the rate of product removal (Hall et al. 1993). Astroglial cells, at least those in culture, resemble muscle (a) in the presence of BCATm as the predominant isoenzyme, (b) in being capable of releasing the BCKA generated from the corresponding amino acids (see above) owing to the fact that the rate of transamination exceeds that of oxidative decarboxylation of the α-keto acids by the branched chain α-keto acid dehydrogenase enzyme complex. In the rat, skeletal muscle is believed to be the primary source of BCKA for oxidation by liver (Hutson et al. 1978). Reamination of BCKA is also thought to occur in extrahepatic tissues (Harper 1989).
In addition to shuttling nitrogen among cells in the brain, oxidation of the BCKA may play a role in brain energy metabolism. Release of the BCKA by astrocytes (Yudkoff et al. 1996; Bixel and Hamprecht 1995) could supply the keto acids for oxidation by neurons, oligodendrocytes, and microglial cells. To determine which types of cells can oxidize BCKA requires information on the cellular location of the corresponding dehydrogenase complex.
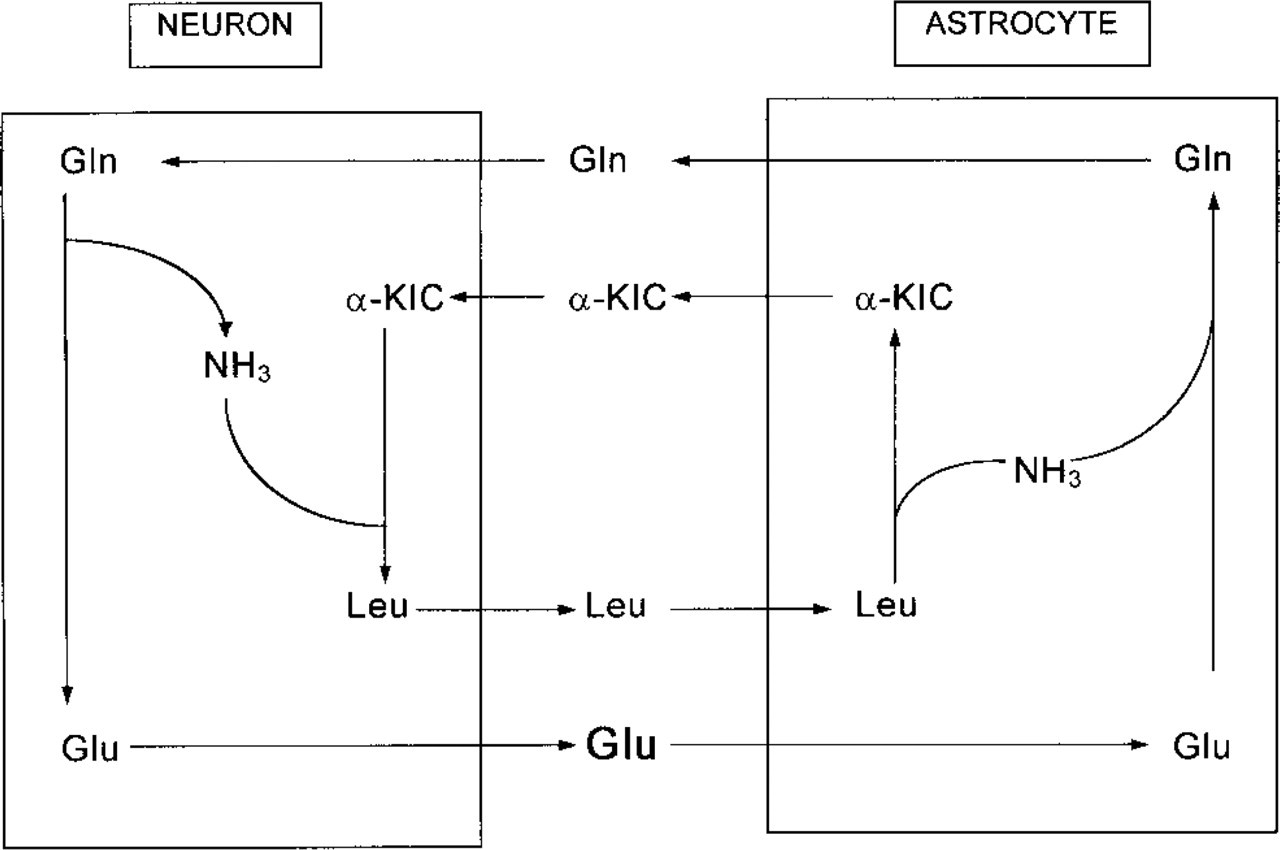
Schematic demonstrating putative exchange of metabolites between astrocytes and neurons. GIn, glutamine; Glu, glutamate; α-KIC, α-ketoisocaproate; Leu, leucine.
The role of BCATc in oligodendroglial cells is not known at this time. If the catabolic pathways are present in these cells, then BCAA or BCKA can be degraded, thus serving as an energy source. The main synthetic function of oligodendrocytes is to produce myelin. Because leucine and isoleucine are eventually degraded to acetyl-CoA, they might also serve as lipid precursors in these cells.
Acknowledgments
Supported by the University of Tübingen, by NATO Collaborative Research Grants Program CRG 950864, and by grant DK 34738 from the National Institutes of Health.
We thank Drs Barbara Ranscht and Frank Walsh for the anti-galactocerebroside and A2B5 antibodies, respectively.
We also thank Dr Brigitte Pfeiffer for helpful suggestions concerning the immunocytochemistry and Dr Heinrich Wiesinger for critically reading the manuscript.