
Abstract
Rat gastrointestinal (GI) tract is rich source of galectins, a family of mammalian galactoside-binding lectins. To determine which tissue component is the relevant glycoconjugate ligand for the galectins, we produced recombinant galectin-1 and surveyed its binding sites on tissue sections of rat GI tract. Mucin and epithelial surface glycocalyces of both gastric and intestinal mucosa were intensely stained. This finding raises the possibility that some GI tract galectins known to be secreted by the epithelia may recognize these glycoconjugates and crosslink them into a macromolecular mass. This galectin-ligand complex may play a role in protecting the epithelial surface against luminal contents such as gastric acid, digestive enzymes, and foreign organisms.
Keywords
G
To test the possibility that galectins recognize any cell and/or ECM component other than laminin, we produced recombinant rat galectin-1 and surveyed its binding site on tissue sections of rat GI tract that are known to be rich not only in glycoconjugates but also in galectins themselves (Leffler et al. 1989). We found that mucin and epithelial cell surface glycocalyces (ECSGs) are intensely stained with recombinant galectin-1.
Materials and Methods
Reagents
The reagents used for SDS-polyacrylamide and agarose gel electrophoresis were obtained from Bio-Rad Japan (Tokyo, Japan). Sepharose 4B was from Pharmacia Biotec (Tokyo, Japan). Restriction enzymes and DNA standards were from Fermentas (Vilnius, Lithuania). Unless otherwise noted, all other reagents of analytical grade were purchased from Sigma (St Louis, MO).
DNA Cloning and Expression of Recombinant Galectin-1
Poly(A) RNA was obtained from frozen rat lungs (Rockland; Gilbertsville, PA) using an mRNA isolation kit (Stratagene; La Jolla, CA). Reverse transcription was done with avian myeloblastosis virus reverse transcriptase (Cetus; Emeryville, CA) using 1–2.5 μM of anti-sense downstream primer 5'-CT
A DNA fragment was cut out from the clone-positive pCR-script vector using NcoI and BamHI restriction enzymes, gel-purified and subcloned in between the same sites in a pET 11d expression vector (Novagen; Madison, WI). A host bacterial strain BL21(DE3) (Novagen) was transformed by the vector and inoculated onto agar plates containing ampicillin. Several colonies were separately picked up into distilled water, boiled, and then colony PCR was done using the gene-specific primer set to ensure successful ligation of the target insert into the expression vector. The insert-positive bacterial clone was separately cultured in 50 ml SOC medium overnight, centrifuged, resuspended in 1 ml SOC medium containing 8% glycerol, numbered, and stored at −80C. Fifty μl from the bacterial stock solution was inoculated into 5 ml SOC medium and then into 200 ml SOC medium at 37C until the absorbance at 600 nm reached about 0.6. Protein production was induced by adding 5 mM IPTG to the culture medium for 2 hr. The bacteria were incubated at 30C during IPTG induction to keep the recombinant protein in a soluble form according to the manufacturer's bulletin. The bacteria were then cooled on ice, collected by centrifugation, and the pellet was resuspended in 20 ml protein extraction solution (PES) containing 58 mM Na2HPO4,18 mM KH2PO4,75 mM NaCl, 2 mM EDTA, and 2 mM PMSF. After one cycle of freeze-thawing, 4 mM 5-iodoacetamido-fluorescein (5-IAF) (Molecular Probes; Eugene, OR) for the labeling experiments or 4 mM iodoacetamide for the control experiment was added into the solution and then the bacteria were lysed with an ultrasound sonificator (Tomy-Seiko; Tokyo, Japan) at maximal power twice for 10 sec. Cell debris was removed by centrifugation at 18,000 X g for 20 min. The soluble extract was loaded into a column containing 50 ml lactose-coupled Sepharose 4B resin and the unbound fraction was extensively washed with PES until the resin became free of the yellowish color of 5-IAF, except for a yellowish band bound to a narrow area (2–3 mm in height) just beneath the surface of the resin. Bound protein was eluted with 20 ml PES containing 100 mM lactose. The eluted protein was dialyzed against 200 ml PES twice for 12 hr at 4C, concentrated using Amicon Y-10 ultrafiltration membranes, and then stored at −80C in the dark. Crude extract from IPTG-induced BL21 and affinity-purified recombinant galectin-1 (hereafter referred to as r-galectin) were analyzed with SDS-PAGE.
Histocytochemistry Using Fluorescein-labeled r-Galectin
Ten Wistar rats (6–8 weeks old) of both sexes were sacrificed with CO2 according to the guidelines of our institutional animal welfare committee. Fundic and pyloric areas were obtained from the stomach. The duodenum (just below the pyloric ring), jejunum (1 cm distal to the duodeno-jejunal junction), and ileum (1 cm proximal to the ileocecal junction) were dissected from each animal and opened. The luminal contents were washed away with jet stream of acetate buffered saline (ABS; 10 mM sodium acetate, 150 mM NaCl, pH 7.4) through a 20-gauge needle attached to a 20-ml plastic injector. The tissues were then cut into small cubes, fixed in 4% paraformaldehyde plus 2.5% glutaraldehyde in 0.1 M acetate buffer for 4 hr, and incubated in 10 mM glycine in the same buffer for 1 hr to quench residual aldehydes. Of the several fixation schedules tested, this schedule was the best one for preserving general tissue structure and galectin binding to cryosections.
For light microscopic histochemistry, the tissue specimens were soaked in 0.6 M sucrose in 0.1 M acetate buffer overnight and embedded in M-1 Embedding Matrix (Lipshaw; Pittsburgh, PA). Cryosections 5 μm were cut in a cryotome (Microm; Walldorf, Germany) and placed on poly-L-lysine-coated glass slides. The sections were covered with blocking solution (BS) containing 10 mM sodium phosphate (pH 7.4), 150 mM NaCl, and 3% bovine serum albumin for 30 min to mask nonspecific protein binding sites. After removal of BS with tissue papers by tilting the slides, the sections were incubated with fluorescein-labeled r-galectin diluted (1 μg/ml BS) with BS for 30 min, washed in PBS (10 mM sodium phosphate, 150 mM NaCl, pH 7.4) three times for 5 min each, and then examined under an epifluorescence microscope (Olympus; Tokyo, Japan).
For electron microscopic cytochemistry, the tissues were infiltrated stepwise in increasing concentrations of sucrose (from 0.6 to 2.3 M sucrose) in 0.1 M acetate buffer (pH 7.4). The tissue blocks were attached to aluminum stubs and snap-frozen in liquid nitrogen. Ultrathin frozen sections 90–100 nm thick were cut with a diamond knife in an Ultracut S microtome equipped with an FC4E cryoattachment (Reichert; Vienna, Austria), and picked up on collodion-coated nickel grids by means of a droplet of 2.3 M sucrose in a platinum loop. The grids were inverted onto a droplet of distilled water to remove sucrose and transferred to a BS droplet for 30 min. The grids were then incubated on a droplet containing fluorescein-labeled r-galectin diluted (0.1 μg/ml BS) with BS for 30 min, washed on a droplet of PBS three times for 15 min each, and then stained with rabbit anti-fluorescein antibody (Molecular Probes) diluted 200 times with BS for 30 min. After washing on a droplet of PBS three times for 15 min each, the grids were floated on a droplet of goat anti-rabbit IgG antibody labeled with 10 nm colloidal gold (Bio-Cell; Cardiff, UK), which was diluted 200 times with BS for 30 min, then washed on ABS three times for 5 min each to remove phosphate ions, and stained on a droplet of 1% aqueous uranyl acetate solution for 30 sec. After brief coating on a droplet of aqueous 1.5% methylcellulose solution, the grids were inverted onto a filter paper, air-dried at 60C for 1 hr, and examined under a 1200EX electron microscope (JEOL; Tokyo, Japan).
In control experiments, fluorescein labeled r-galectin was preincubated with 10 mM thiodigalactopyranoside (TDG) for 30 min, anti-fluorescein antibody was omitted from the staining steps, or non-labeled r-galectin (treated with non-labeled iodoacetamide) was used.
Statistical Analysis
Fundus, jejunum, and ileum were obtained from three rats. Each GI tract site was cut into three tissue blocks and processed for cryosectioning as above. Differences in the degree of r-galectin binding to mucin granules of each GI tract site were compared by counting the number of gold particles per unit area of granules. The electron-dense core region of fundic mucin granules, which was devoid of gold labeling, was excluded from counting. To assess intestinal mucin labeling, goblet cells located in the middle one third of villous processes were examined by careful trimming of the tissue blocks during cryosectioning. At least 0.25 μm2 X 4 areas within mucin granules of 10 fundic surface mucous cells or intestinal goblet cells were examined per tissue block per animal. Data were expressed as the mean ± SEM and were analyzed with Fisher's least significant difference (LSD) method. The numbers of gold particles attached to microvilli of enterocytes located in the middle third of villous processes of jejunum and ileum were counted in the same way as above and were analyzed by Student's t-test.
Results
Preparation of Recombinant Galectin-1
By reverse transcription of lung poly(A) RNA followed by PCR amplification using two primers flanking the entire coding sequence of rat galectin-1, we obtained about a 420-base pair (BP) DNA product (not shown) that matched the expected molecular mass of rat galectin-1 DNA. The DNA product was ligated into an expression vector and recombinant protein production was triggered with IPTG. We found a protein band migrating at molecular weight of 14.5 kD in the crude extract of bacteria (Figure 1). The extract was then passed through a lactose-coupled resin column. The lactose-eluted fraction contained a single protein band at a molecular weight of 14.5 kD (Figure 1), indicating that biologically active r-galectin was successfully produced. Because the r-galectin was proved to be a good histochemical probe (see below), no further detailed characterization of the protein was performed.
For use as a histochemical probe, r-galectin was irreversibly alkylated with 5-IAF (or non-labeled iodoacetamide), because this procedure stabilizes the protein against oxidative inactivation of the sugar-binding activity. Under these conditions, the protein can be stored for at least 3 months (Whitney et al. 1986) without any loss of activity. This procedure also attaches a histocytochemical marker to the protein, which enables us to localize the protein-binding sites on tissue sections directly under a fluorescence microscope and indirectly under an electron microscope by means of anti-fluorescein antibody.
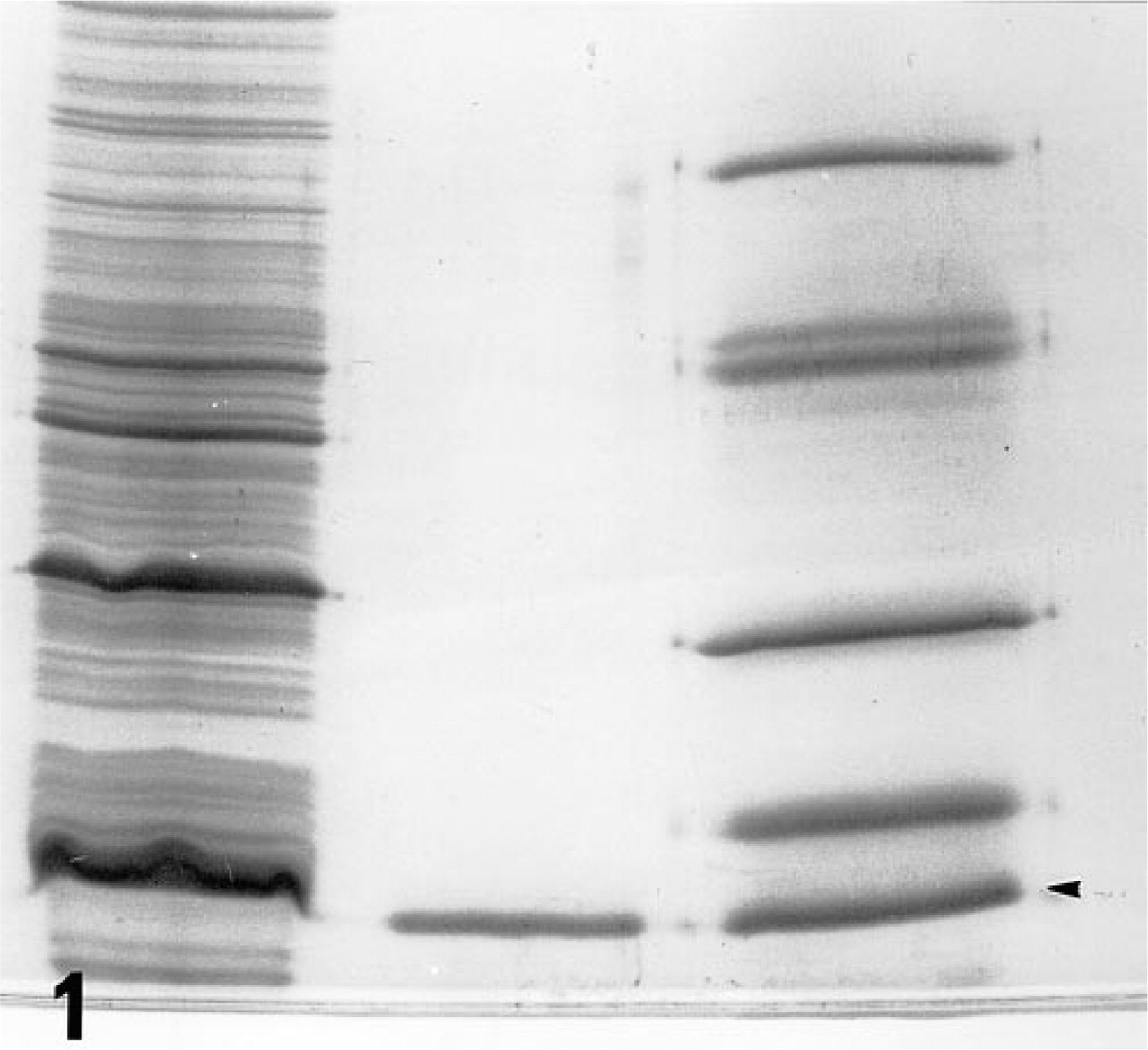
SDS-PAGE analysis of recombinant protein. A crude extract of IPTG-treated bacteria with recombinant vector coding the entire reading frame of rat galectin-1
Binding Sites of r-Galectin in Gastrointestinal Tract
Fluorescence microscopic examination revealed that r-galectin recognizes distinct cell populations and cellular sites. In the fundic glands of the stomach, the apical cytoplasm of surface mucous cells and the mucous neck cells were intensely stained (Figure 2). Large, scattered oval cells located in the middle portion of the glands were also stained. In the pyloric glands, the apical cytoplasm of surface mucous cells was intensely labeled (Figure 3). In the small intestine (Figures 7 and 8), the epithelial brush border was intensely stained. The staining appeared to be much denser on the surface of villi than on that of crypts. Scattered under this brush border were goblet cell granules which were also stained with r-galectin. Secretory granules of Paneth cells and Brunner's gland cells were barely stained. When r-galectin was preincubated with TDG, all staining was completely abolished (not shown).
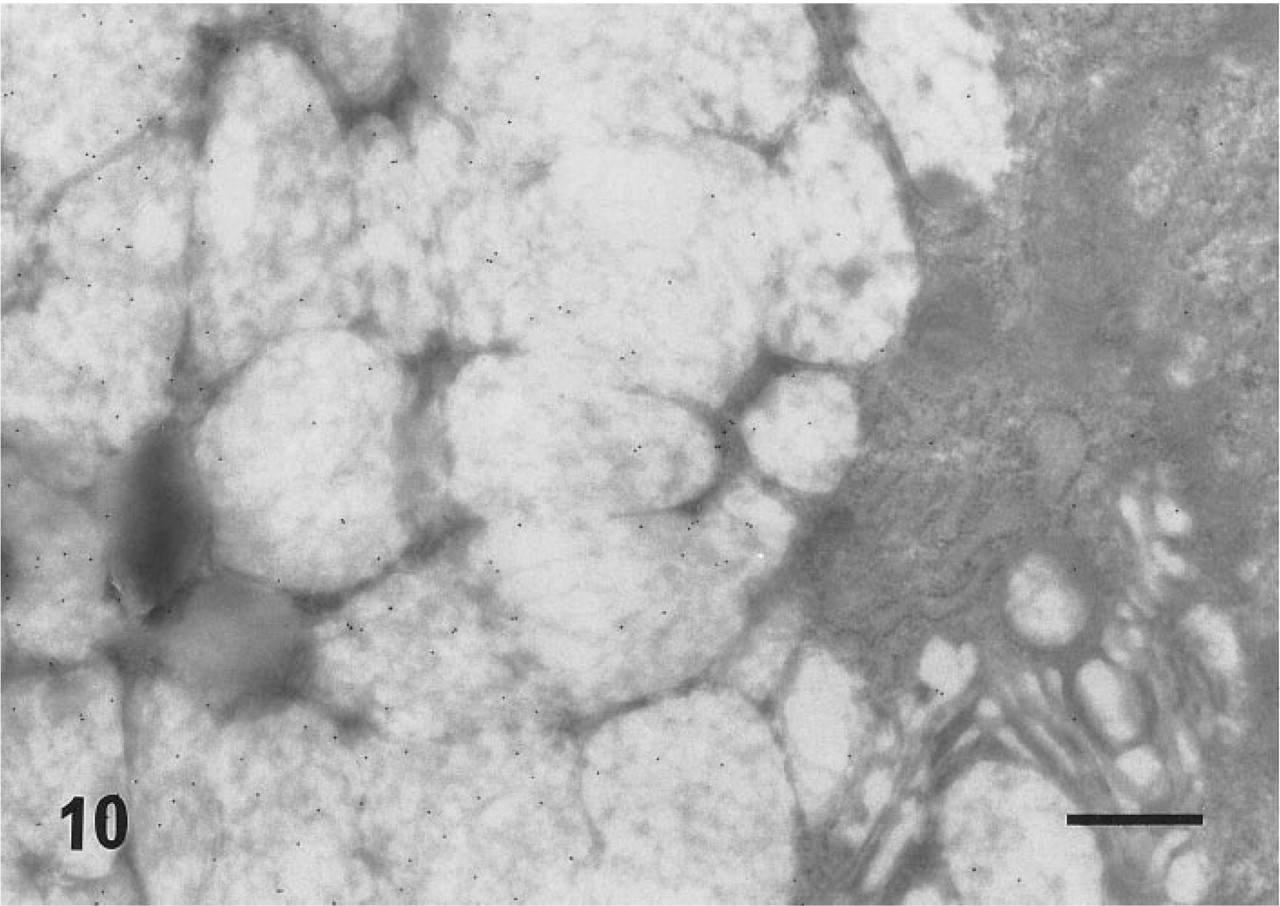
In ultracryosections, the secretory granules of both surface mucous cells (Figure 4) and mucous neck cells of the fundic glands were intensely labeled. Although the mucosa was thoroughly washed with a jet stream of physiological saline before fixation, apparent labeling was observed on the apical plasma membrane of the surface mucous cells (Figure 5). The cell surface and the intracytoplasmic canaliculi of parietal cells were also labeled (Figure 6), indicating that the large oval cells seen in the fundic glands are parietal cells. The secretory granules and surface membranes of the pyloric surface mucous cells were also labeled (not shown). The microvilli of enterocytes from duodenum to ileum were intensely labeled with r-galectin (Figure 9). The secretory granules of intestinal goblet cells were also labeled (Figure 10), but the staining density appeared to be much sparser compared with that of the mucin granules of gastric glands and the microvilli of adjacent enterocytes. Incubation of r-galectin with TDG before staining (Figure 11) completely abolished immunogold labeling in all r-galectin-reactive GI tract sites.
Statistical analysis (Figure 12) of the labeling density of fundic and intestinal mucin granules verified that mucin of the fundic surface mucous cells (150.3 ± 15.92/μm2) showed significantly (p<0.001) higher affinity to galectin-1 than did goblet cell mucin of the jejunum (28.9 ± 1.95/μm2) and ileum (30.3 ± 2.13/μm2). There was no significant difference in the labeling density of goblet cell mucin (above values) and apical microvilli (jejunum 95.8 ± 3.45/μm2 vs ileum 102.9 ± 4.75/μm2) between the two distinct intestinal regions examined.
Discussion
To examine galectin-binding sites in GI tract tissues, we chose galectin-1 as a cytochemical probe among several kinds of GI tract galectins for the following reasons. (a) This protein undergoes little post-translational modification, except for the N-terminal methionine cleavage and acetylation (Clerch et al. 1988), such as intramolecular phosphorylation, that is known to occur in galectin-3 and influence the function of this protein (Huflejt et al. 1993). This permits galectin-1 to be produced in a relatively simple bacterial expression system. (b) The protein is known (Cooper et al. 1991) to remain as a soluble protein compared with galectin-4 (Chiu et al. 1994), which is produced as insoluble form when expressed in bacteria. (c) The protein can be labeled with iodoacetamido-fluorescein via the intramolecular cysteine residues (Cooper et al. 1991; Whitney et al. 1986) without attaching a cytochemical marker such as an extra epitope sequence of viral origin. We therefore cloned the entire reading frame of rat galectin-1 DNA (Clerch et al. 1988) by RT-PCR technique and expressed the protein in E. coli. The recombinant protein could be purified through lactose-coupled resin and ran as only a single band in SDS-PAGE gel with a molecular weight of 14.5 kD, which is identical to the known molecular weight of endogenous galectin-1. This indicates that the r-galectin is biologically active and usable as a cytochemical probe for detecting endogenous ligand(s) for this protein or possibly its related protein family. The r-galectin recognized intensely the mucin and ECSG within the GI tract tissues. This finding was surprising to us, because previous studies (Gu et al. 1994; Cooper et al., 1991) have suggested that one of the ligands for galectin-1 is laminin, a large glycoprotein constituting basement membranes. The reason why the basement membrane (laminin) of the GI tract was not recognized by r-galectin is unclear. However, one possible interpretation is that laminin may be fully occupied by endogenous galectin-1 so that no more substrate is available for a newcomer, exogenous r-galectin. This idea is consistent with our previous immunohistochemical study (Wasano et al. 1990) showing that endogenous galectin-1 is externalized by intestinal smooth muscle cells into their pericellular ECM space to interact with the glycoconjugate ligand located there. To confirm this assumption, however, a procedure should be developed by which one can remove endogenous galectin from its binding sites before staining with an exogenous galectin probe.
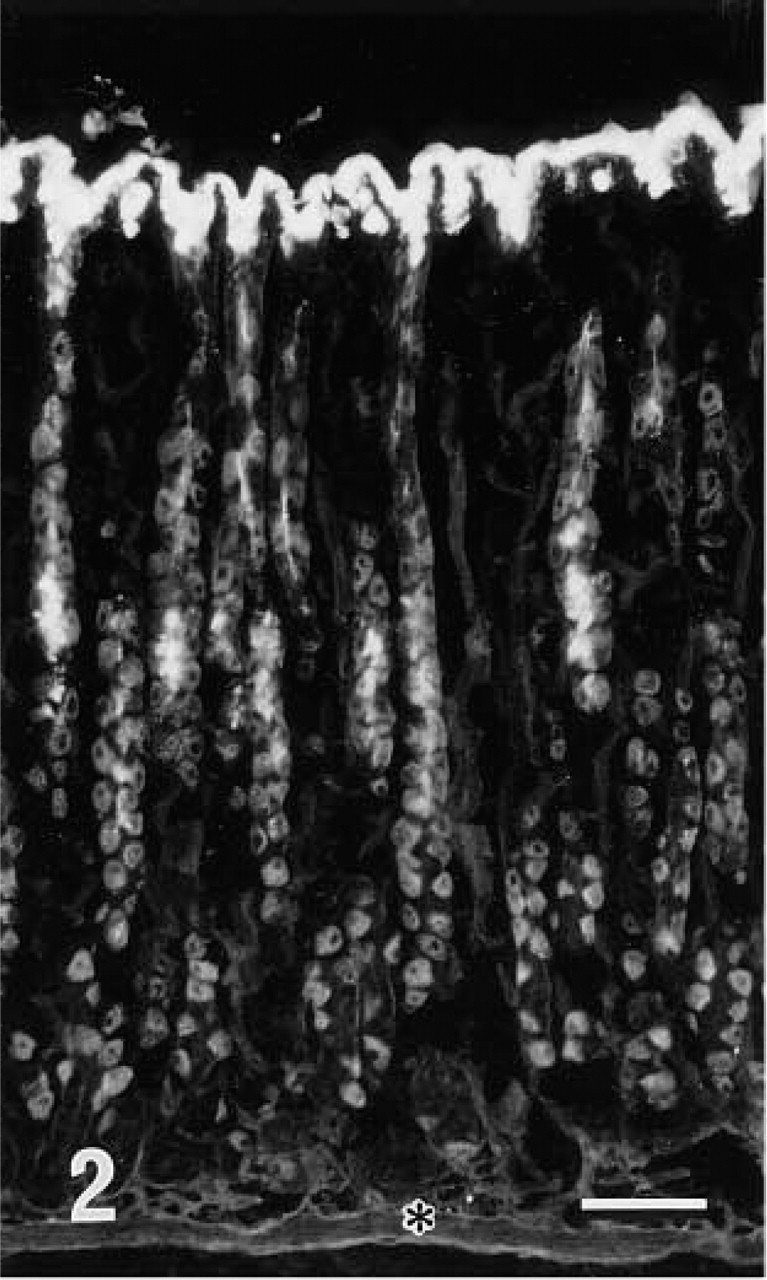
Fluorescence micrograph of fundic glands stained with r-galectin labeled with 5-IAF. Cells covering the mucosal surface and cells located at the neck portion of the glands are stained. In the middle two thirds of the glands, large oval-shaped cells are also stained. The position of the lamina muscularis mucosae (LMM) is indicated by an asterisk. Bar = 100 nm.
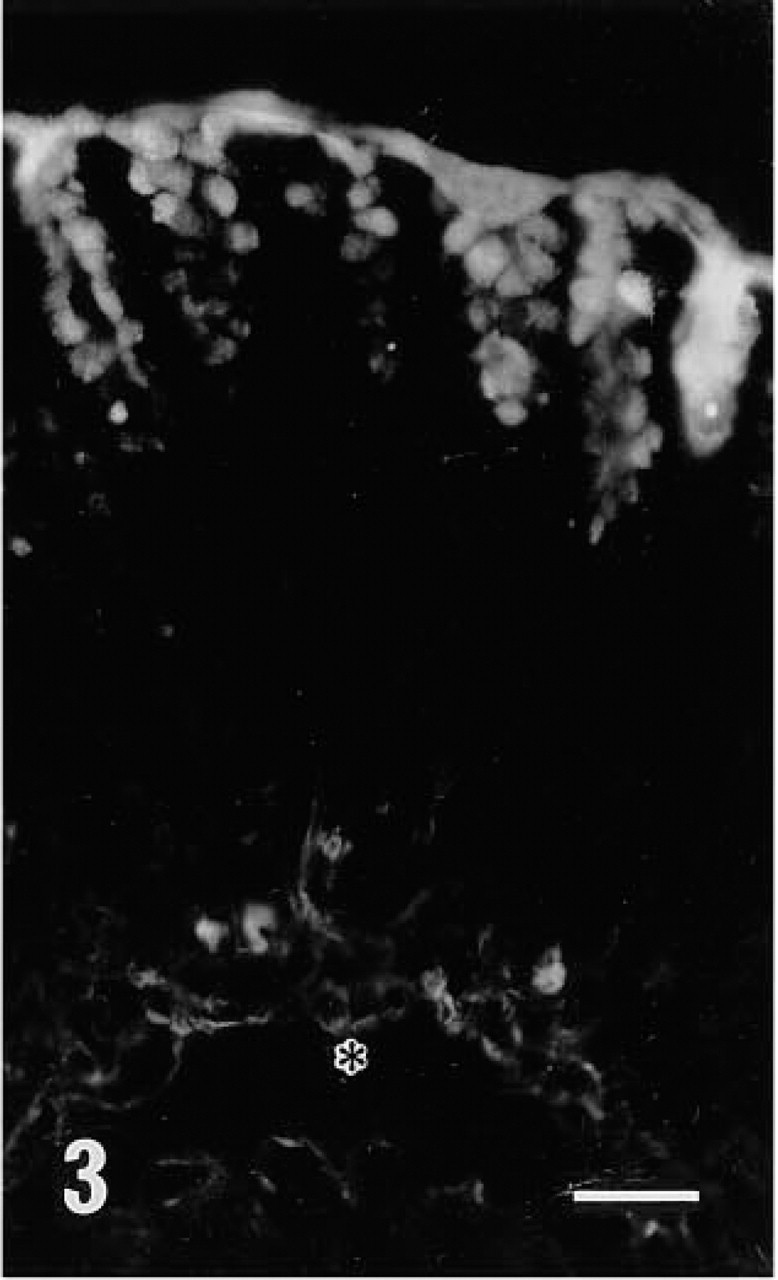
Fluorescence micrograph of pyloric glands stained with r-galectin labeled with 5-IAF. Surface mucous cells are positive. The position of LMM is indicated by an asterisk. The nature of several weak fluorescent structures seen at the bottom of the glands is unclear. Bar = 100 nm.
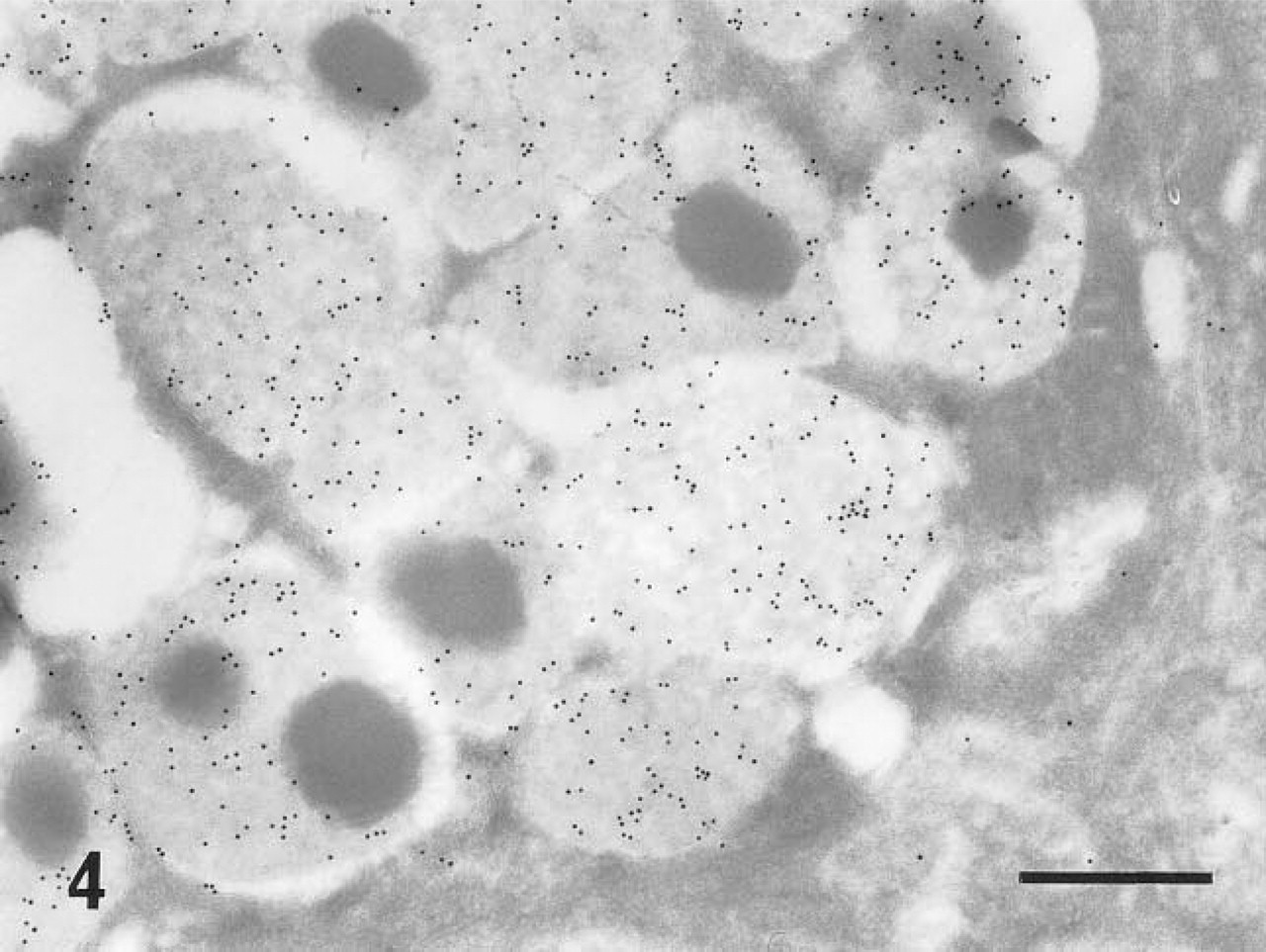
Electron micrograph of secretory granules of a surface mucous cell of a fundic gland. This specimen was stained with r-galectin labeled with 5-IAF, and galectin-binding sites were indirectly detected by anti-fluorescein antibody followed by a secondary antibody conjugated with 10-nm colloidal gold (r-galectin-immunogold sequence). The granule contents are intensely labeled. Several gold particles are also seen on a Golgi apparatus located in the lower right corner. Bar = 400 nm.

Apical cell surface of a surface mucous cell of a fundic gland labeled with the r-galectin-immunogold sequence. Gold particles (arrowheads) are seen closely attached to the apical plasma membrane. Bar = 200 nm.
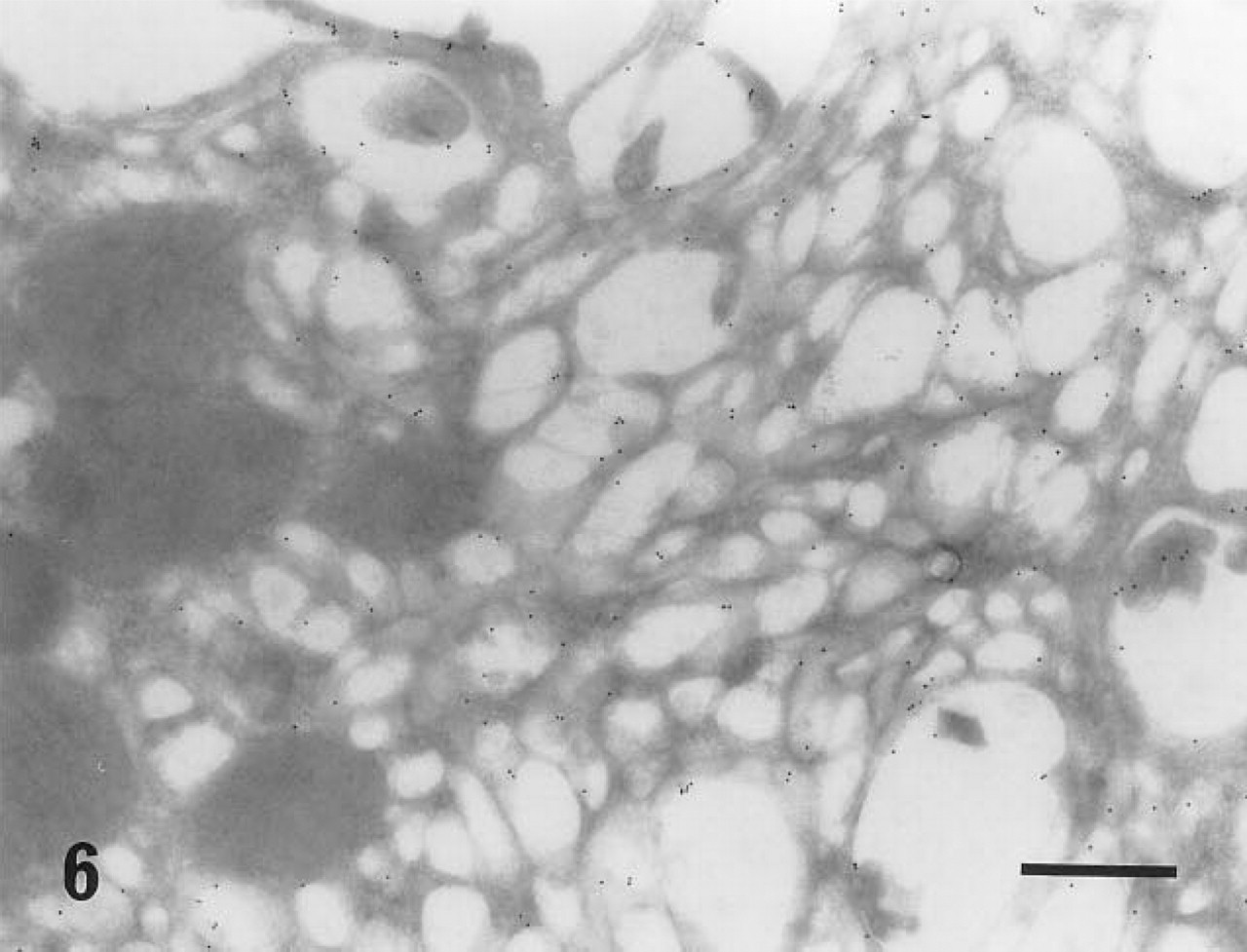
Intracytoplasmic canaliculi of parietal cells stained with the r-galectin-immunogold sequence. Gold particles are seen on the luminal surface of the canaliculi. No gold particles are found on several mitochondria on the left side of this micrograph. Bar = 500 nm.
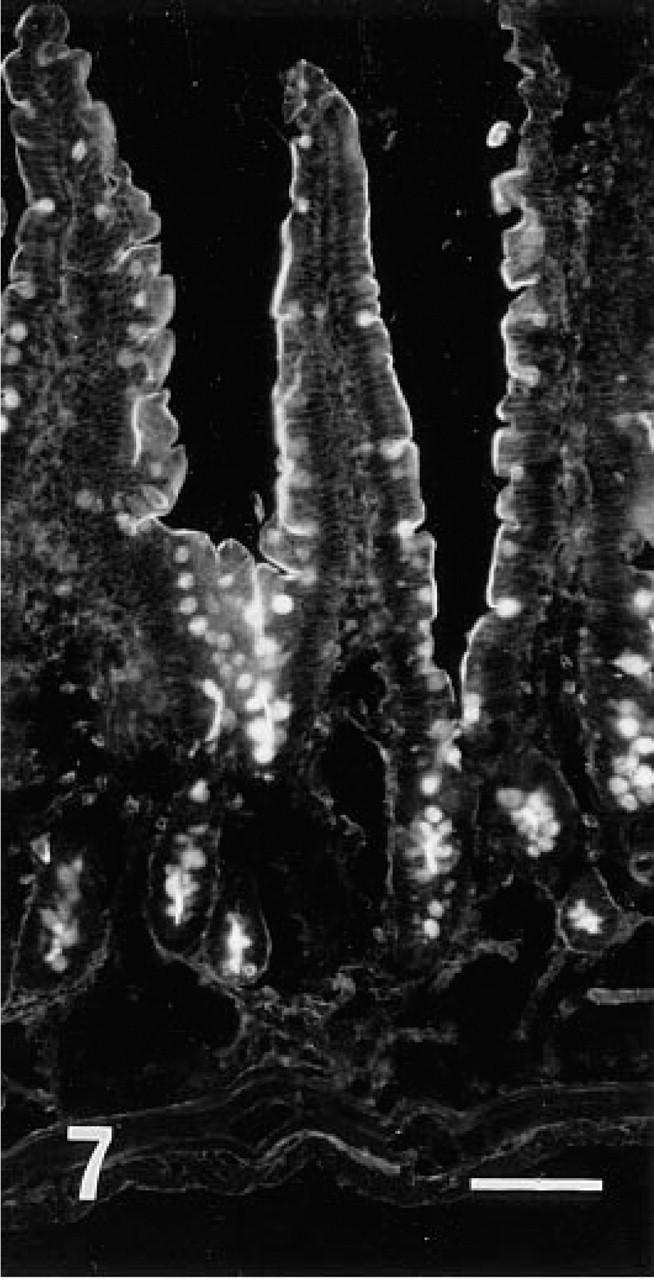
Fluorescence microscopic view of jejunum stained with r-galectin labeled with 5-IAF. Brush border and goblet cell granules are stained. Bar = 100 nm.
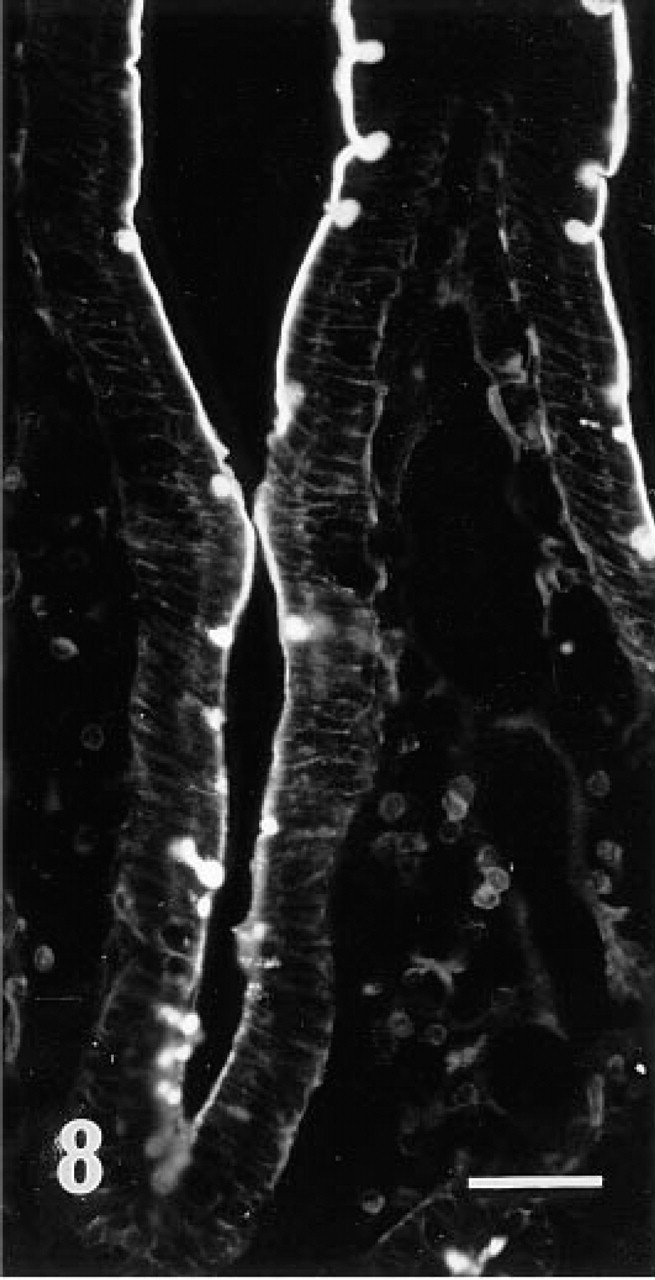
A close-up view of the villi stained with r-galectin labeled with 5-IAF. The brush border of the upper villous portion appears to be more intensely stained than that of the lower portion. Bar = 200 nm.
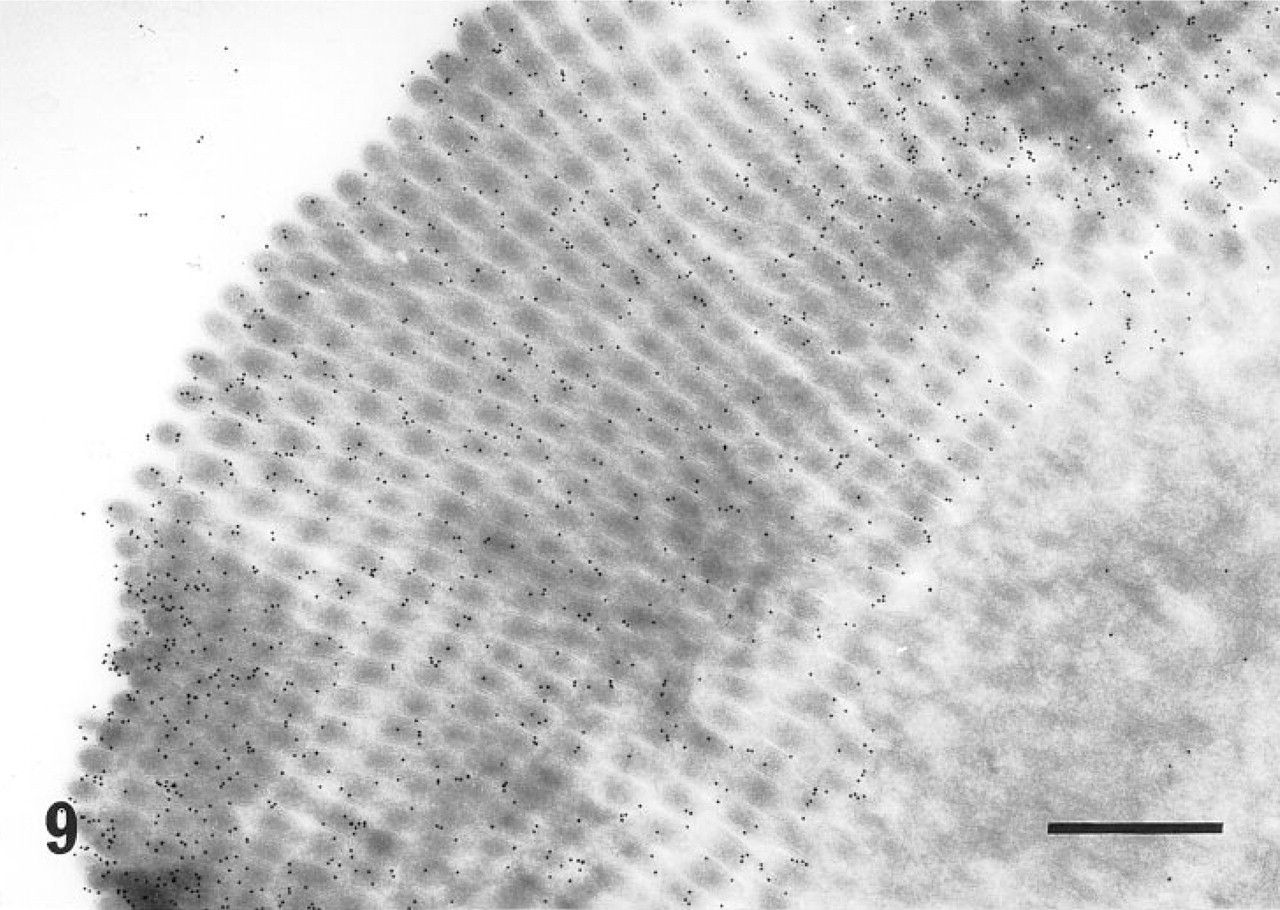
Electron micrograph of the apical surface of an enterocyte from the jejunum, stained with the r-galectin-immunogold sequence. The microvilli are densely labeled with immunogold particles. Bar = 500 nm.
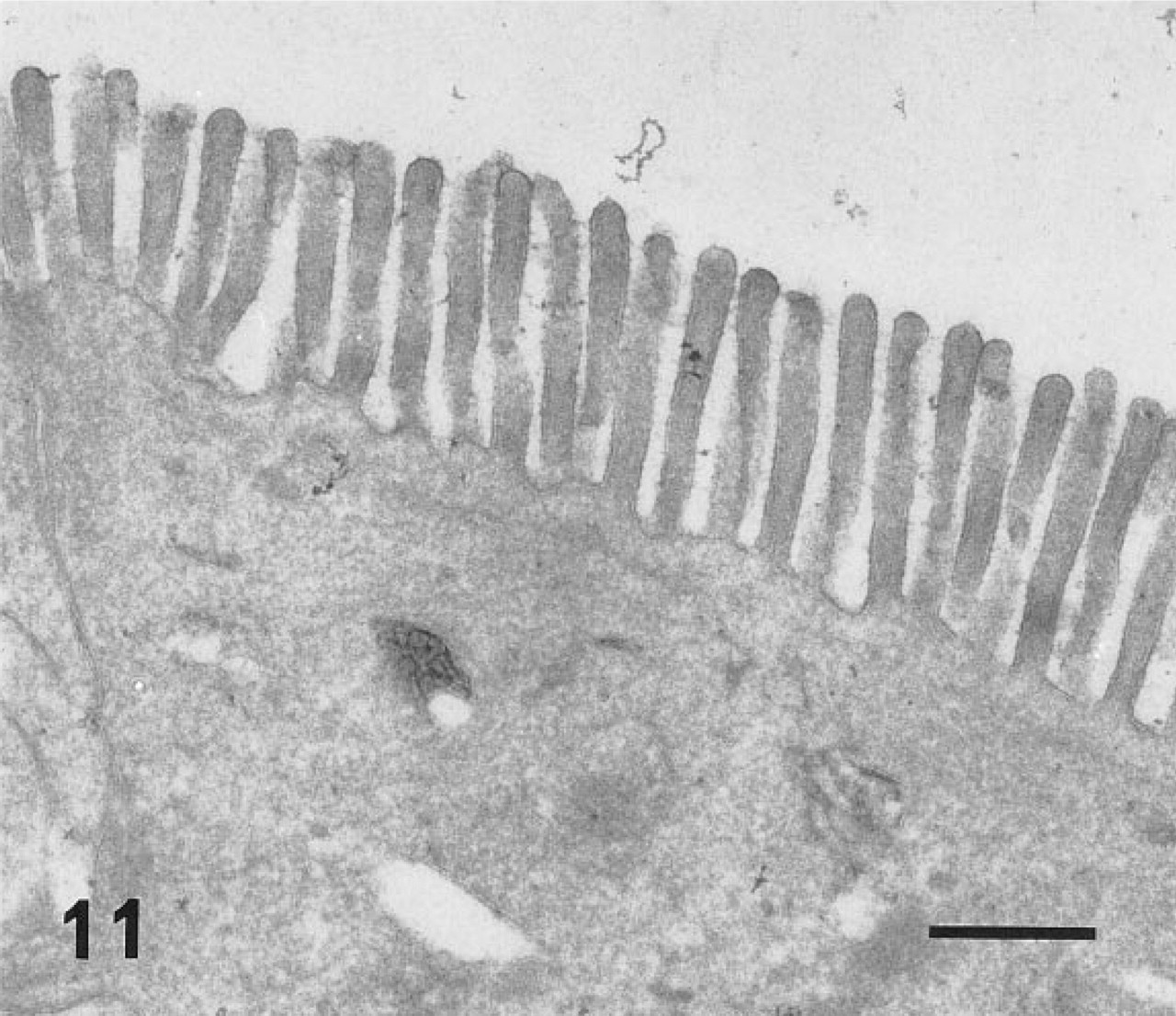
Control specimen. An enterocyte from the jejunum stained with the r-galectin-immunogold sequence, except that r-galectin-5-IAF was incubated with 10 mM TDG before staining. No specific gold labeling is seen on the microvilli. Bar = 500 nm.
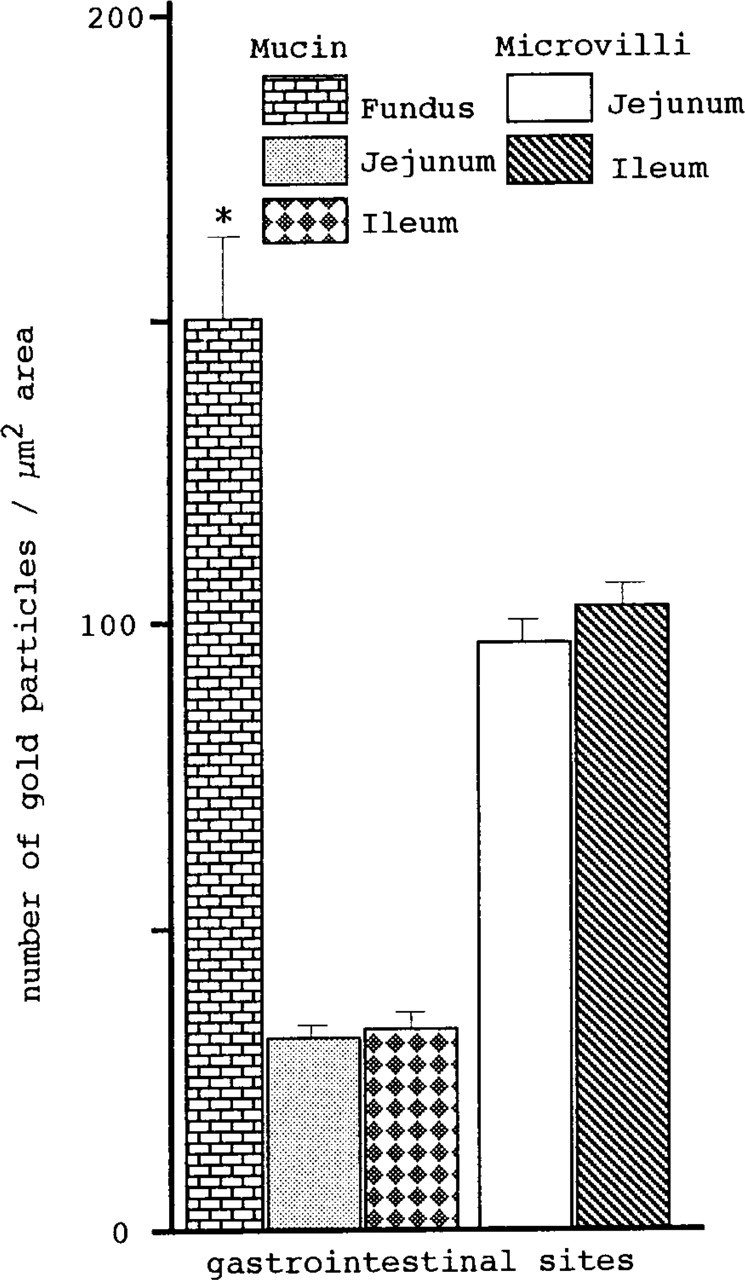
Different affinity of GI tract sites for galectin-1, evaluated by counting the number of gold particles. Total gold numbers on 0.25 × 4 μm2 areas within mucin granules randomly selected from 10 cells per tissue block per animal were counted. The fundic mucin exhibits significantly (*p<0.001 by Fisher's LSD) higher affinity for galectin-1 than does intestinal mucin. In contrast, mucin of goblet cells between two different intestinal regions does not show any significant difference. There is no significant difference (by t-test) in the labeling density of microvilli between two distinct intestinal regions. Data are means ± SEM (n = 90 for each Gltract site).
Are the mucin and ECSG natural ligands for galectin-1? Galectin-1 has been shown to be localized in muscle tissues (Poirier and Robertson 1993; Cooper et al. 1991; Wasano et al. 1990), in some neurons (Hynes et al. 1990), and in cultured fibroblasts (Roff and Wang 1983), but not in any epithelial tissue. These findings suggest that galectin-1, even if it is externalized into ECM (Barondes et al. 1994), is confined to the subepithelial connective tissue of the GI tract and therefore appears not to directly interact with luminal mucin or ECSG. In this regard, it is of interest that rat GI tract tissue has recently been shown (Lindstedt et al. 1993; Oda et al. 1993; Leffler et al. 1989) to contain two more galectins, galectin-3 and -4. Both of these galectins are postulated to be produced by GI tract epithelial cells and secreted extracellularly. All members of the galectin family exhibit a common property of strongest binding to oligosaccharides composed of a terminal galactosyl and penultimate glucosyl residue (Barondes et al. 1994; Leffler and Barondes 1986) among a battery of oligosaccharides tested. This suggests that the mucin and ECSG strongly recognized by galectin-1 may possess such oligosaccharide chains, which are also excellent ligands for another type of GI tract galectin such as galectin-3 or -4. This notion is partly supported by observations (Lindstedt et al. 1993; Sparrow et al. 1987) that mucin-type oligosaccharide chains inhibit binding of galectin-3 to lactose. It is possible, therefore, that galectin-3 or -4 externalized by the GI tract epithelia into the lumen may participate in crosslinking mucin to mucin and mucin to ECSG. Such crosslinking may efficiently prevent the loss of mucin into the lumen and protect the epithelial surface from the attack of luminal acid, digestive enzymes, and/or microorganisms. Indeed, the presence of such a system has been documented in chicken intestine (Barondes 1984; Beyer and Barondes 1980). Chicken intestine goblet cells secrete a kind of galectin called CLL-II (also called CL-16) into the intestinal lumen, along with mucin. This protein has high affinity for both intestinal mucin and ECSG and is thought to play a role in crosslinking these two glycoconjugates. Although CLL-II is 50% identical in its amino acid sequence with galectin-1 (Barondes et al. 1994), no functional counterpart to this protein has been established in mammalian GI tract.
The intracellular canaliculi of parietal cells were also stained with r-galectin. In the portion of the fundic glands where the parietal cells are located, there are neither r-galectin-positive secretory cells nor secretion adjacent to the cells. Therefore, the labeling appears to be caused by this cell type's own ECSG. Although the specialized intracellular structure of the parietal cells is known to be involved in acid secretion, it is unclear whether the r-galectin-positive ECSG is related to such function.