
Abstract
Cyclosporine A (CsA) is the first immunosuppressant used in allotransplantation. Its use is associated with side effects that include nephrotoxicity. This study explored the anatomic structures involved in CsA nephrotoxicity and the effect of heme oxygenase (HO) in preventing CsA injury. Rats were divided into four groups, which were treated with olive oil, CsA (15 mg/kg/day), CsA plus the HO inhibitor (SnMP; 30 μM/kg/day), and with the HO inducer (CoPP; 5 mg/100 g bw). Renal tissue was treated for morphological, biochemical, and immunohistochemical studies. CsA-treated rats showed degenerative changes with renal fibrosis localized mainly around proximal tubules. Collapsed vessels were sometimes seen in glomeruli. No HO-1 expression and increased expression of endothelin-1 (ET-1) were observed in CsA-treated rats compared with controls. In CsA plus SnMP-treated rats, HO-1 expression was further reduced and the morphology was not changed compared to the CsA group, whereas CsA plus CoPP-treated animals again showed normal morphology and with restoration and an increase in HO-1 levels. HO activity and immunohistochemical data showed similar alterations as HO expression. No changes were observed for HO-2 analysis. The observations indicate that HO-1 downregulation and ET-1 upregulation by CsA might be one mechanism underlying CsA-induced nephrotoxicity. Therefore, attempts to preserve HO levels attenuate CsA nephrotoxicity.
T
The most common feature of CsA use is the development of interstitial fibrosis, which can be patchy or “striped” (Verpooten et al. 1986; Rezzani et al. 2001b). This may promote development of chronic renal failure in organ transplant recipients, although long-term studies have shown that, with the dose regimens now employed, renal function can remain stable but impaired for many years (Bantle et al. 1990). The mechanism of CsA-induced interstitial fibrosis has been attributed to an angiotensin-dependent upregulation of molecules that promote scarring, such as TGF-β1 (Ahmed et al. 2004) and osteopontin (Lim et al. 2004). CsA may also impair the regenerative capacity of microvascular endothelial cells and induce apoptosis (Li et al. 2004). To this end, thrombotic microangiopathy is a distinct form of CsA-induced vascular toxicity that may result from a direct effect on vascular endothelium, possibly interfering with generation of prostacyclin (Rezzani et al. 2001b).
It is unknown whether renal tissue possesses a defense mechanism against CsA-induced injury. Heme oxygenase (HO), the rate-limiting enzyme in heme catabolism, has been shown to attenuate the extent of various forms of renal injury, most notably injury due to ischemia and to inflammation (Wolf et al. 1994; Datta et al. 1999). HO is a microsomal enzyme that catalyzes the NADPH-, O2-, and CYP450 reductase-dependent oxidation of heme to carbon monoxide (CO), iron, and biliverdin, which is reduced to bilirubin by biliverdin reductase (Abraham et al. 1996). Two catalytically active HO isoenzymes have been characterized, HO-1 and HO-2. HO-1 is induced by a host of stimuli that have in common the ability to cause oxidative stress. HO-2 is a constitutive form and, to date, only the adrenal glucocorticoids have been identified as inducers of its gene (Abraham et al. 1996). In the rat kidney the HO-1 isoform is distributed in proximal convoluted tubules and loops of Henle (Da Silva et al. 2001). HO controls the initial and rate-limiting step in heme ca-tabolism, thereby regulating cellular heme levels and generation of CO. In this dual capacity, HO controls both the availability of heme for synthesis of heme-containing enzymes and the generation of CO, which binds to these enzymes and thus modulates their activity. Examples of heme-containing enzymes that have been implicated in the pathophysiology of CsA-induced nephrotoxicity are CYP450 (Mayer et al. 1989), the arachidonate cyclo-oxygenases (COXs) (Jenkins et al. 2001), TxA2 synthase (Perico et al. 1986), and NOS (Amore et al. 2000). It follows that changes in HO expression or enzyme activity in response to CsA would impact on the activity of heme-containing enzymes. The ensuing changes in product synthesis, i.e., prostaglandin, TxA2, and NO, might modulate the extent of CsA-induced renal vasoconstriction and cell injury.
The hypothesis tested is that HO expression is associated with the attenuation of CsA-induced nephrotoxicity. Moreover, we examined by electron microscopic analysis whether the negative CsA effects on intracellular structures might be related to their major susceptibility to drug metabolism and the anatomic structures that were considered to be the intrarenal target for CsA damage.
Materials and Methods
Animal Treatment
Forty male Sprague-Dawley rats (average weight 200–250 g) were used. The animals were divided into four groups and treated for 21 days. Group I (n = 10) was injected SC with olive oil (the CsA vehicle) and served as control. Group II (n = 10) was treated with CsA alone (15 mg/kg/day) for 21 days. Group III (n = 10) was treated with CsA and the inhibitor of HO activity stannous mesoporphyrin (SnMP; 30 μmol/kg). Group IV (n = 10) received one dose of the HO inducer cobalt protoporphyrin (CoPP; 5 mg/100 g bw) after 3 days of CsA treatment. All animals were sacrificed at the end of each treatment period. Kidney cortical sections from each rat were homogenized for Western blot analysis of HO-1 and HO-2 expression and for enzyme activity assay. A portion of the kidney tissue was fixed in formalin and was processed for sectioning (5-μm sections) and staining for identification of collagen using the Azan method and for HO and ET-1 immunohistochemistry (IHC). Another portion of kidney was fixed in 2.5% glutaraldehyde, dehydrated, and treated by standard procedures for electron microscopic analysis.
Renal Morphology
For tubulointerstitial fibrosis, the quantitative scorings used were similar to those reported by Li et al. (2004). A minimum of 20 fields per section was assessed and graded using a color image analyzer (Image-Pro Plus, Media Cybernetics Inc.; Silver Spring, MD). The extent of tubulonterstitial fibrosis was estimated by counting the percentage of areas injured per field using a scoring scale of 0 to 3+, where 0 = normal in-terstitium, 0.5 = <5% injured area, 1 = 5–15% injured area, 1.5 = 16–25% injured area, 2 = 26–35% injured area, 2.5 = 36–45% injured area, and 3 = >45% injured area.
Western Blot Analyses
All cortical homogenates (protein lysates), obtained by transverse cutting near the renal apex, were processed for Western blot analysis and protein levels were visualized by immunoblotting with rabbit antibodies against human HO-1 or HO-2 (Stressgen Biotechnologies; Victoria, BC, Canada). Briefly, 30 μg of lysate was separated by SDS-PAGE and transferred to a nitrocellulose membrane (Amersham; Piscataway, NJ) using a semidry transfer apparatus (Bio-Rad; Hercules, CA). The membranes were incubated with 5% milk in 10 mM Tris-HCl (pH 7.4), 150 mM NaCl, 0.05% Tween-20 (TBST) buffer at 4C overnight. After washing with TBST, the membranes were incubated with a 1:2000 dilution of anti-HO-1 or anti-HO-2 antibodies for 1 hr at room temperature (RT) under constant shaking. Membranes were then washed and probed with horseradish peroxidase-conjugated donkey anti-rabbit IgG (Amersham) at a dilution of 1:2000. Chemiluminescence detection was performed with the Amersham ECL detection kit according to the manufacturer's instructions. The bands were analyzed densitometrically using an image analyzer (Photo-Capt V.99 software).
Statistical Analysis
The results are presented as mean ± SEM of the number of triplicate determinations. Statistical significance of differences between the experimental groups was estimated using the ANOVA and Bonferroni test, with p<0.01 considered as significant.
Measurement of HO Enzyme Activity
Kidneys were perfused with ice-cold HEPE's solution, sliced, and cortical tissue was homogenized in 10 mM Tris buffer, pH 7.5, containing 0.25 M sucrose. Homogenates were centrifuged at 27,000 × g for 20 min at 4C. The supernatant was centrifuged at 105,000 × g for 1 hr at 4C and the resulting microsomal pellet was resuspended in 0.1 M potassium phosphate buffer, pH 7.6. Protein concentration was measured according to the method of Bradford (BioRad). Homogenates were used for measurement of HO activity, which was assayed with an NADPH generating system as previously described (Morimoto et al. 2001). The amount of bilirubin generated was determined by a scanning spectrophotometer (Lambda 17 UV/VIS; Perkin-Elmer Cetus Instruments; Norwalk, CT) and was defined as the difference between 460 and 530 nm (extinction coefficient 40 mM−1 for bilirubin). Results were expressed as nanomoles of bilirubin per milligram of protein per 30 min.
Immunohistochemical Analysis for HO-1, HO-2, and ET-1 Proteins
The serial sections were immersed in 3% hydrogen peroxide in methanol for 30 min to block endogenous peroxidase activity. The sections were then incubated with goat serum (Dakopatts, Glostrup, Denmark; diluted 1:5) for 40 min, and successively with rabbit polyclonal anti-HO-1 and HO-2 antibodies (diluted 1:100) and rabbit polyclonal anti-ET-1 antibody (diluted 1:50) for 2 hr. The sections were washed in Tris-buffered saline (0.1 M, pH 7.4) and incubated with biotinylated goat anti-rabbit immunoglobulin and avidin-biotin-horseradish peroxidase complex according to the manufacturer's instructions (ABC kit; Dakopatts).
Specificities of antibody labeling were investigated using appropriate controls, incubating the tissue sections with Tris-buffered saline instead of the primary or secondary antibody.
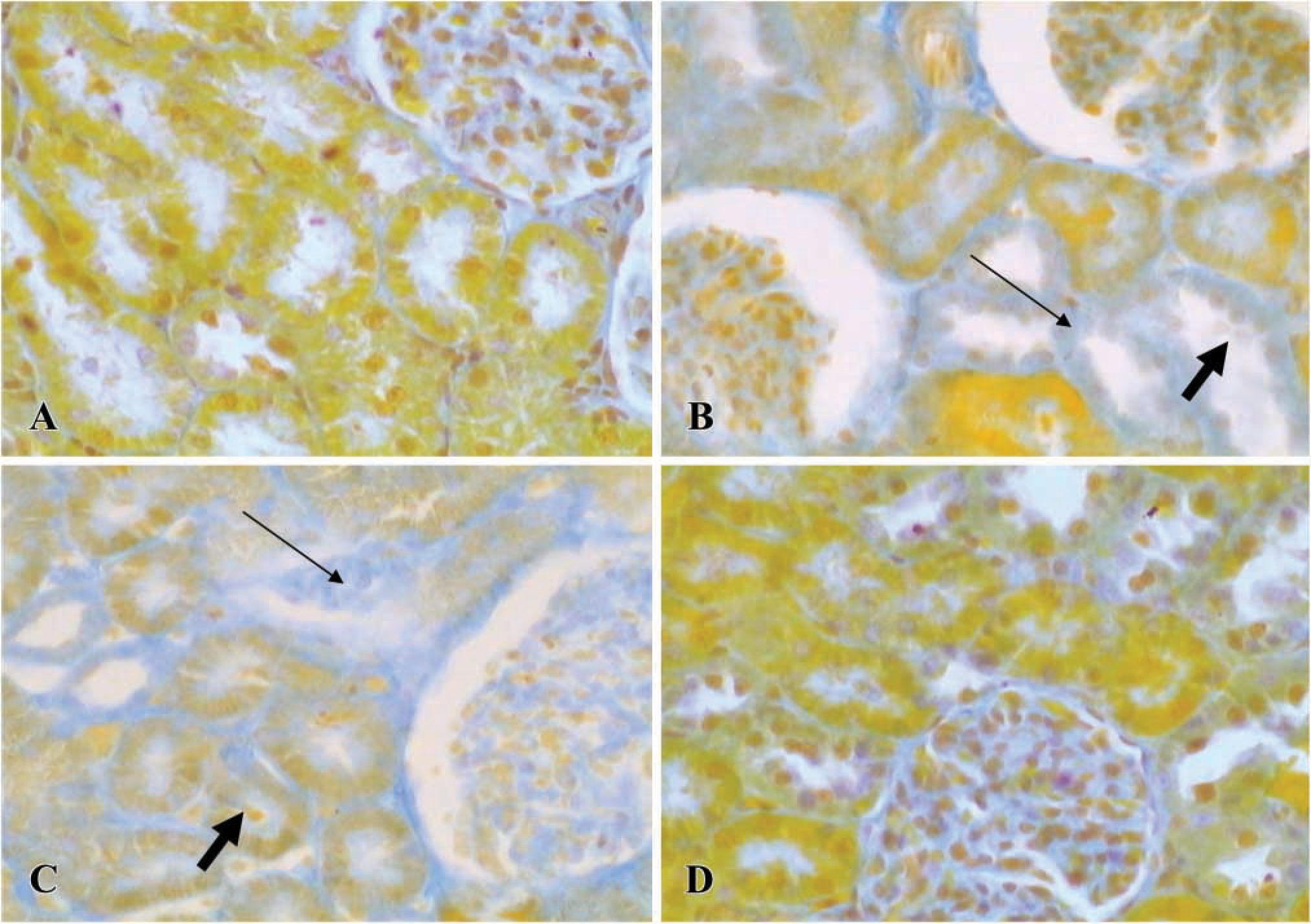
Photomicrographs showing kidney, stained by Azan staining, in control (
Semiquantitative Analysis of HO-1, HO-2, and ET-1 Immunostaining
Intensity of the immunostaining within the glomeruli, proximal tubules, and distal tubules was evaluated separately and blindly. Proximal tubules were differentiated from distal tubules as follows: proximal tubules had larger diameters than distal tubules. The nuclei of proximal tubule epithelial cells were usually arrayed at the base of the cells. Most proximal tubule epithelium had obvious brush borders. Based on the intensity and distribution of HO staining, the degree of HO staining was graded as ± when the staining was very weak, + when the staining was weak, + + when the staining was moderately positive, and + + + when the staining was strong.
Results
Morphological Findings in Control, CsA Alone, CsA plus SnMP-, and CsA plus CoPP-treated Rats
Figure 1A shows that the kidneys had a normal morphology. The cortex showed a large number of glomeruli with proximal and distal tubules located around the latter. The electron microscopic analysis revealed that epithelial cells of proximal and distal tubules showed normal mitochondria near the nucleus. They were also randomly distributed in the cytoplasm or concentrated at the sites of high energy utilization (Figure 2A). The cristae and the membrane were clearly evident and were normal, as shown in the inset of Figure 2A. Renal injury was evident mainly in the cortex of kidneys from the CsA alone and CsA plus SnMP- treated animals. The injury consisted of tubulointerstitial changes visible as narrow stripes of fibrosis localized mainly around the proximal tubules, as well as lack or swelling of epithelial cell nuclei (Figures 1B and 1C). On our scoring system, a significant increase in fibrosis was observed in CsA-treated rats compared with controls (1.8 ± 0.2 vs 0.22 ± 0.04; p<0.01). The fibrosis increase in CsA-treated animals was similar to that observed in the CsA plus SnMP-treated rats (1.8 ± 0.2 vs 1.6 ± 0.3). These data are reported in Table 1. Moreover, glomerula showed changes in morphology. In particular, we sometimes observed a collapse of vessels in glomerula, as previously reported by other authors (Kim and Suh 1995) with an experimental treatment of CsA at the same dose used here. The most evident negative effects consisted of injury mainly in the proximal aspect of distal tubules. Electron microscopic analysis of tubule structures in kidneys treated with CsA and CsA plus SnMP showed giant mitochondria that were round or oval, rarely cigar-shaped, and which lay adjacent to the nuclei; the morphology of these giant mitochondria lacked any specificity (Figure 2B). They usually exhibited few cristae (Figure 2B, inset), and sometimes contained paracrystalline inclusions. The animals treated with CsA plus CoPP showed a morphological pattern similar to that of controls (Figure 1D). In addition, the quantitative analysis for CsA plus CoPP-treated rats was similar to that of control rats (0.3 ± 0.09 vs 0.22 ± 0.04). These data are reported in Table 1. The data showing normal renal cytoarchitecture were also confirmed with the electron microscopic analysis (data not shown).
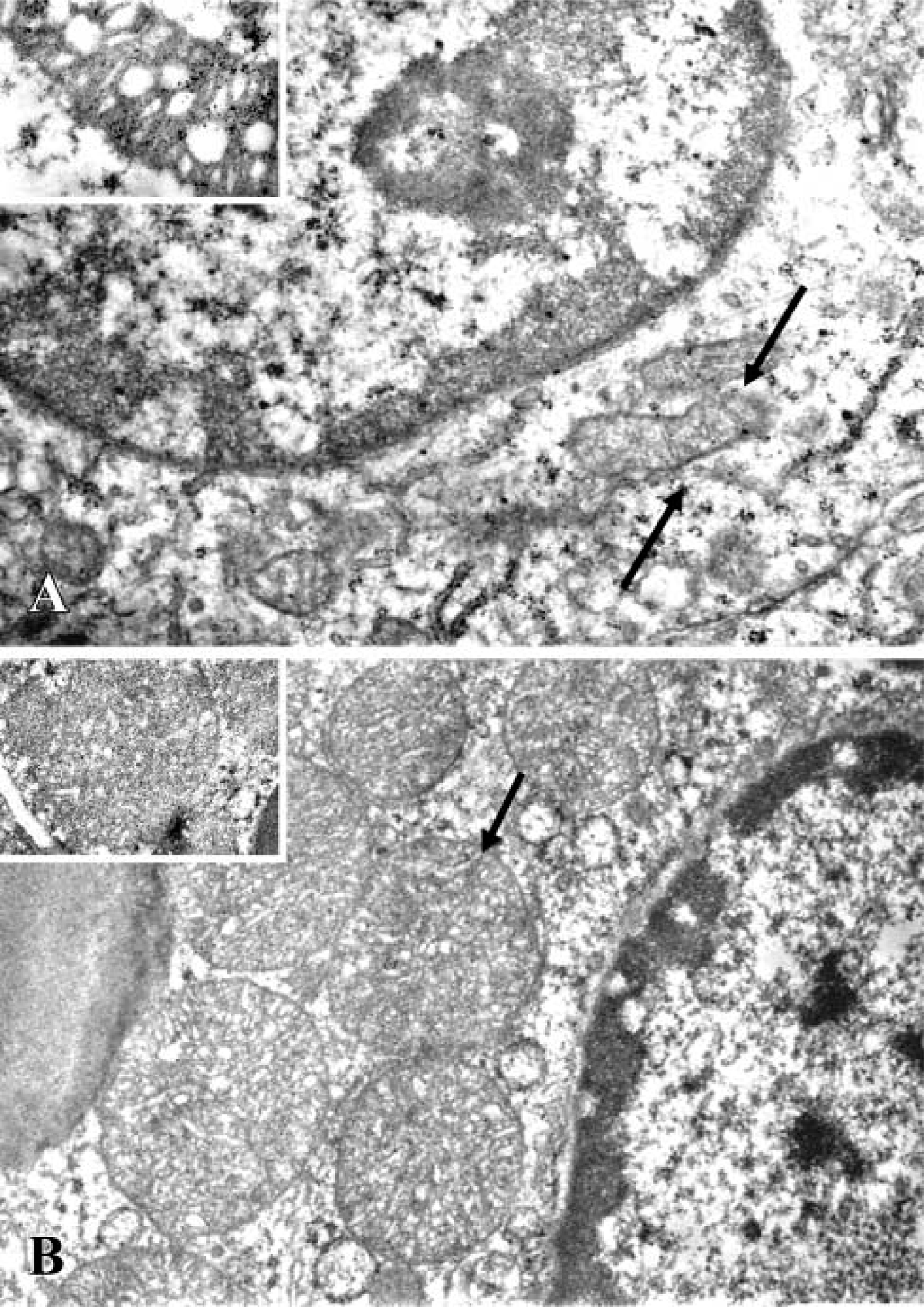
Electron photomicrographs showing (
Effects of CsA Alone, CsA plus SnMP, and CsA plus CoPP Treatment on HO-1 and HO-2 Protein Expression Levels
We studied the effect of CsA levels of HO-1 and HO-2 protein in cortical tissue homogenates by Western blot analysis. Basal levels of HO-1 protein were detectable in control (Figure3A, Lane 3) but not in CsA-treated rat kidneys (Figure 3A, Lane 1). In animals treated with CsA followed by administration of the HO inducer CoPP, renal HO-1 protein levels were restored and also increased (Figure 3A, Lane 4; compare with Lanes 1, 2, and 3). SnMP treatment induced a decrease in renal HO-1 protein levels (Figure 3A, Lane 2). In contrast, there was no significant difference in HO-2 levels among the different experimental groups (Figure 4A). The HO quantitative analyses are shown as histograms in Figures 3B and 4B.
Quantitative score of tubulointerstitial fibrosis of control, CsA, CsA plus SnMP, and CsA plus CoPP-treated rats a
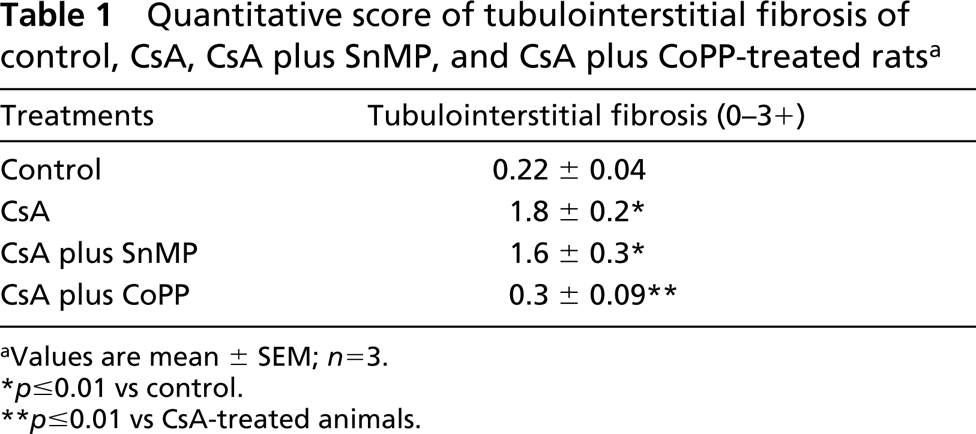
Values are mean ± SEM; n=3.
p≤0.01 vs control.
p≤0.01 vs CsA-treated animals.
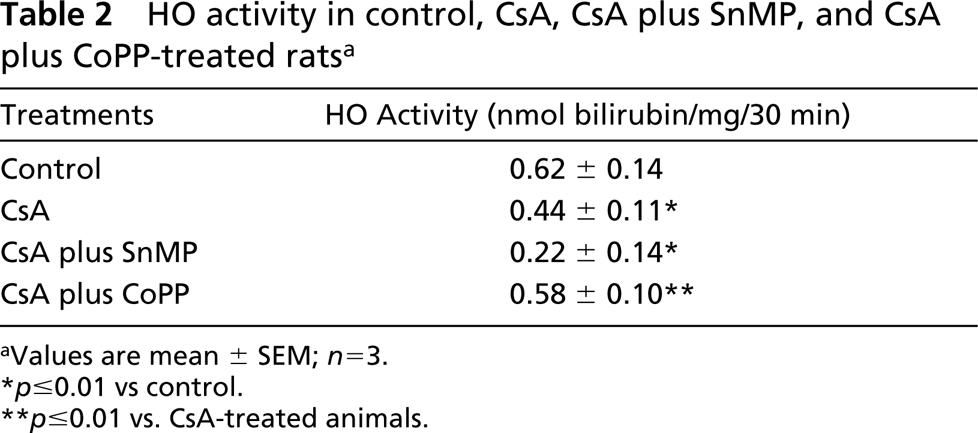
Effects of CsA Alone, CsA plus SnMP, and CsA plus CoPP Treatment on HO Enzyme Activity
In CsA-treated animals, renal cortical HO activity was reduced. To determine the extent of this reduction, a separate group of CsA-treated rats received the HO activity inhibitor SnMP. In these animals renal cortical HO activity was further reduced compared with levels in animals treated with CsA alone. The animals treated with CsA and CoPP showed HO activity levels similar to those of the control group (Table 2).
Effects of CsA Alone, CsA plus SnMP, and CsA plus CoPP Treatment on HO-1, HO-2 Protein Immunostaining
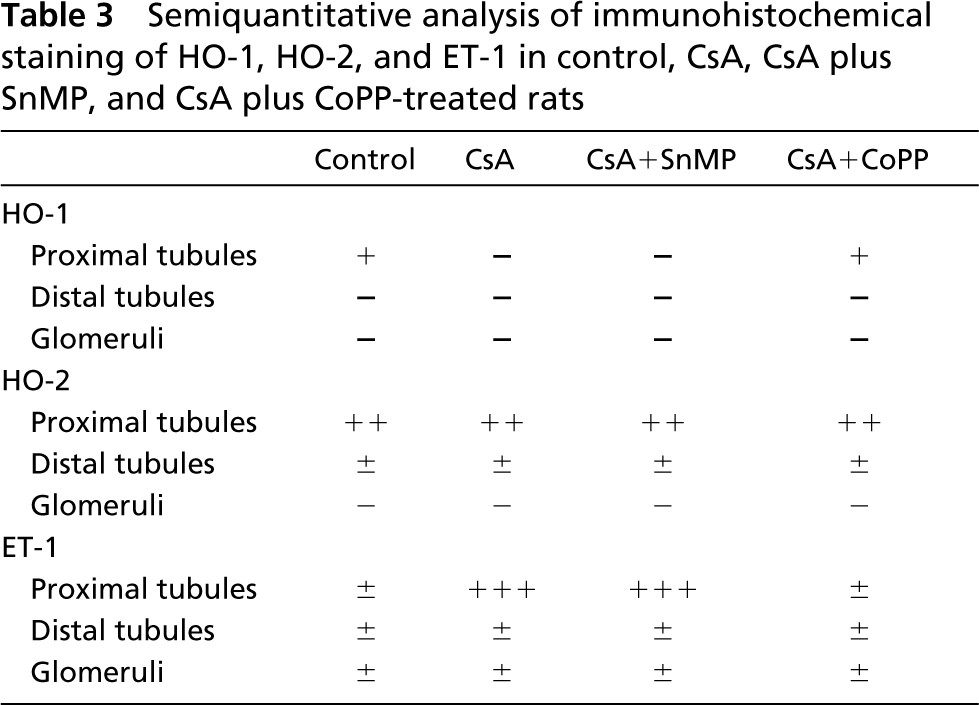
We examined by IHC analysis the effect on the expression of HO-1 and HO-2 protein. In control rats, HO-1 protein showed weak positivity (+) in the epithelial cells of proximal tubules; glomerula and distal tubules were negative (-) (Figure 5A). Conversely, we observed that HO-1 expression decreased in renal parenchyma and was undetectable both in CsA alone and CsA plus SnMP-treated rats (Figures 5B and 5C). HO-1 expression was again similar to control in CsA plus CoPP-treated animals (Figure 5D).
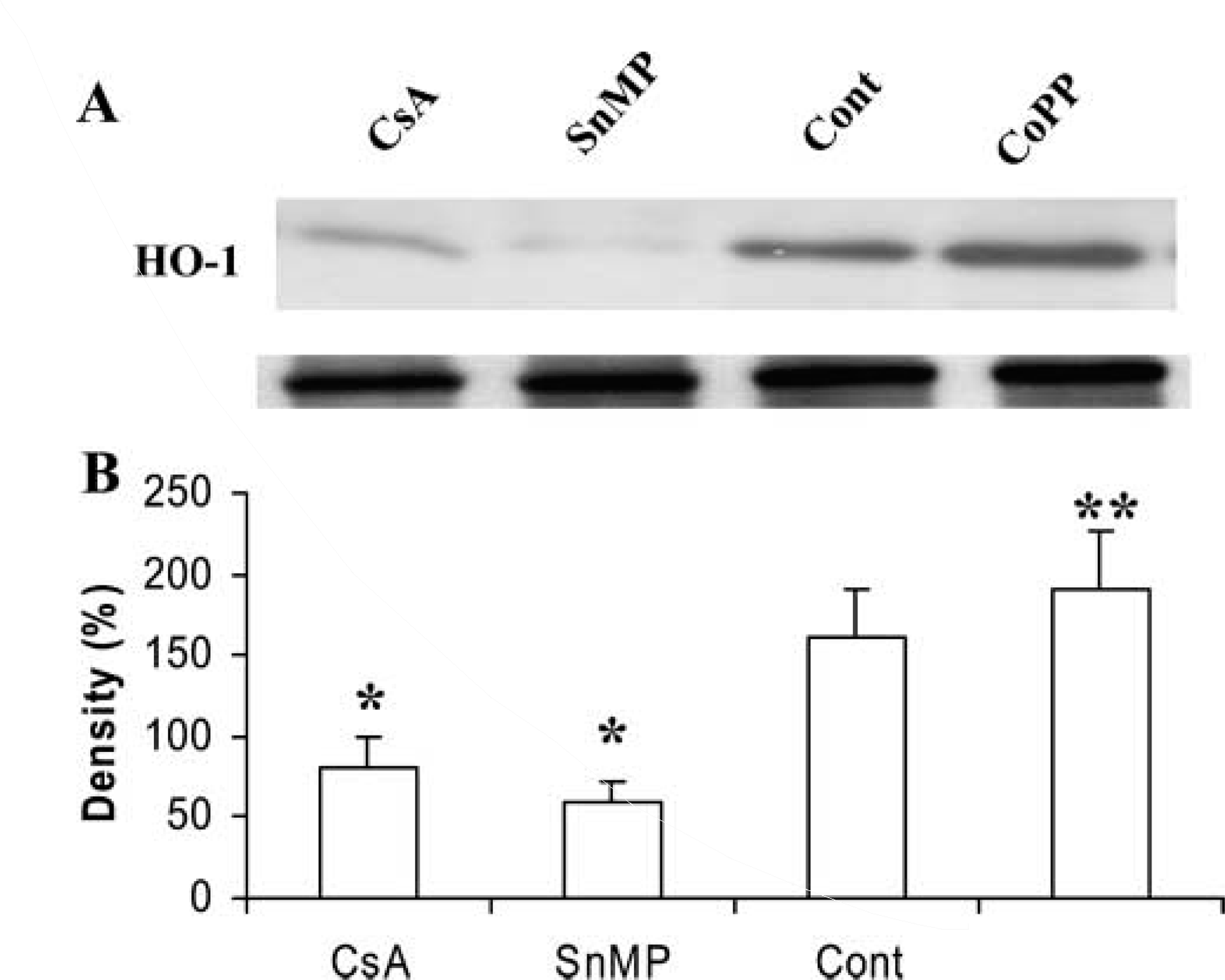
Western blot analysis of HO-1 in renal tissue from CsA-treated rats (Lane 1), CsA plus SnMP (Lane 2), control (Lane 3), and CsA plus CoPP (Lane 4)-treated rats (with β-actin as control). Representative blots with densitometric readings are expressed as percent of control. Significant differences: ∗p<0.01 compared with control; ∗∗p<0.01 compared with CsA-treated group.
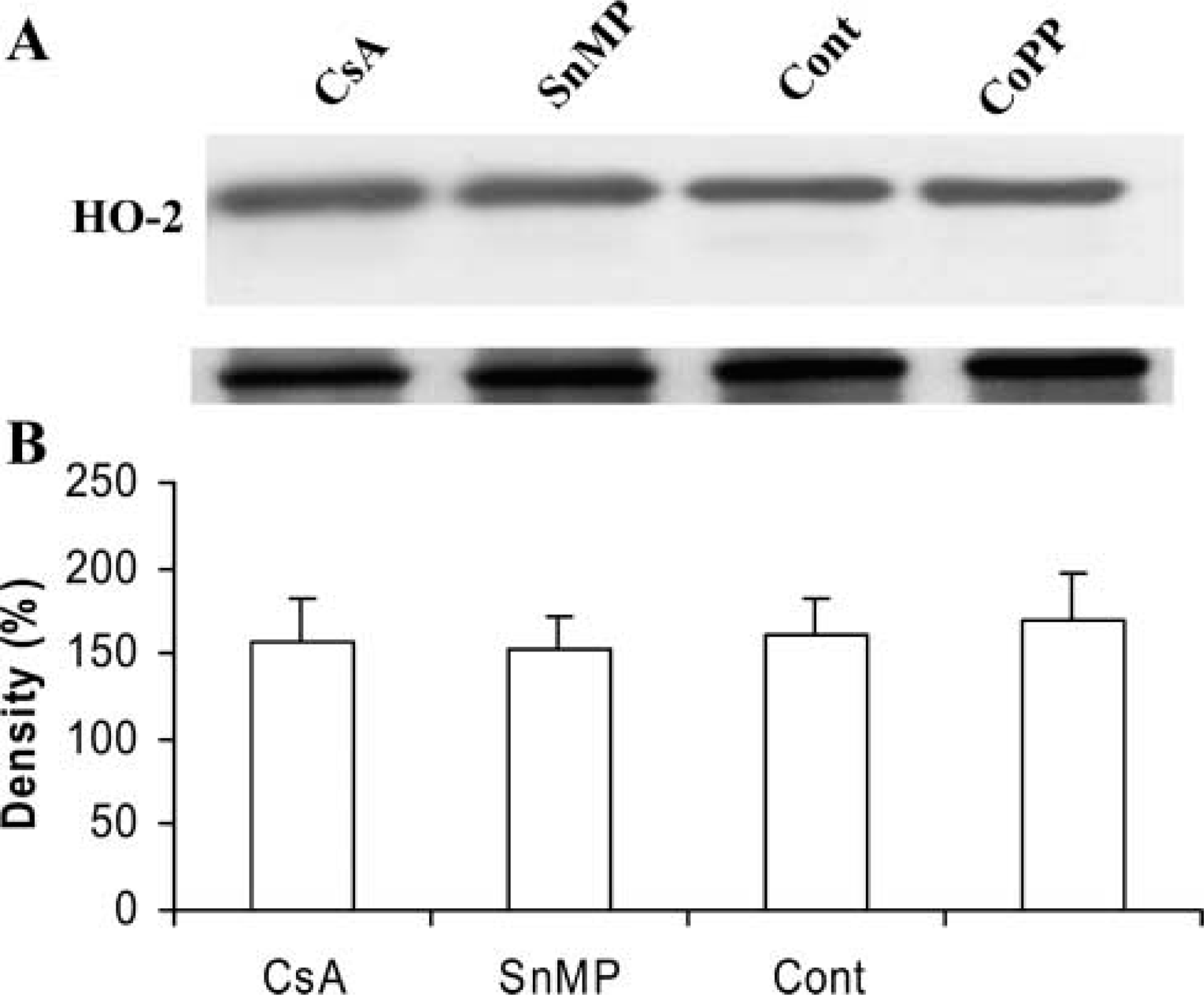
Western blot analysis of HO-2 in renal tissue from CsA-treated rats (Lane 1), CsA plus SnMP (Lane 2), control (Lane 3), and CsA plus CoPP (Lane 4)-treated rats (with β-actin as control). Representative blots with densitometric readings are expressed as percent of control. Significant differences: ∗p<0.01 compared with control; ∗∗p<0.01 compared with CsA-treated group.
HO-2 expression did not differ among the different experimental groups. In fact, HO-2 expression showed moderate positivity (+ +) in proximal tubules and very weak positivity (±) in distal tubules, whereas glomeruli were negative (-) (data not shown).
All semiquantitative data are reported in Table 3.
Effects of CsA Alone, CsA plus SnMP, and CsA plus CoPP Treatment on ET-1 Protein Immunostaining
In control rats, ET-1 protein showed very weak (±) positivity in proximal and distal tubules and in glomerula. Conversely, in CsA-treated rats we observed evident differences in the pattern of ET-1 positivity compared with controls. Proximal tubules were strongly (+ + +) positive, whereas ET-1 staining was very weak (±) in glomeruli and distal tubules. These results confirmed the data reported in our previous work (Rezzani et al. 2001a). No IHC variations compared to CsA-treated and control animals were found, respectively, after treatment with SnMP or CoPP. All semiquantitative data are reported in Table 3.
HO activity in control, CsA, CsA plus SnMP, and CsA plus CoPP-treated rats a
Values are mean ± SEM; n=3.
p≤0.01 vs control.
p≤0.01 vs. CsA-treated animals.
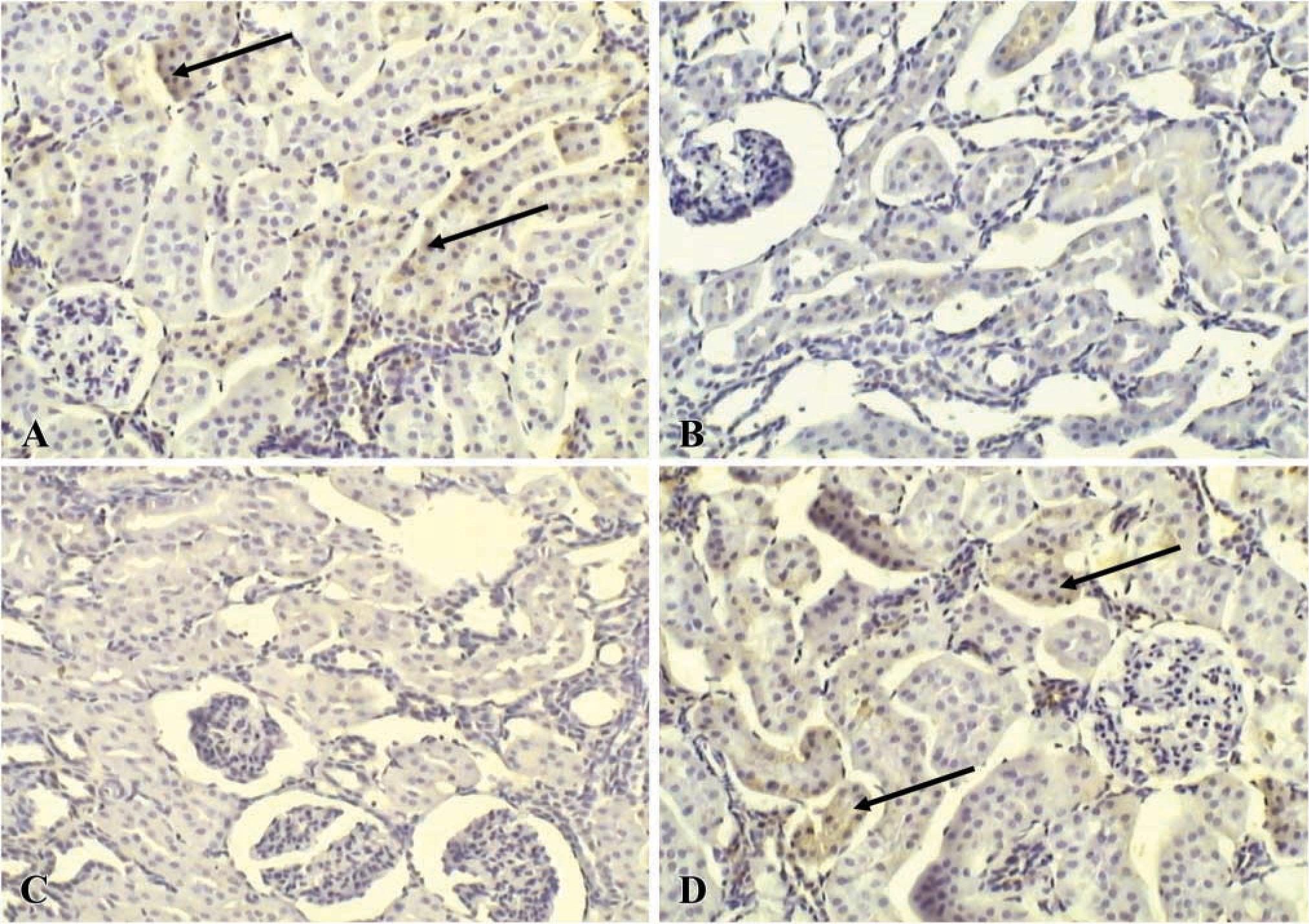
Immunolocalization of HO-1 protein in kidney of control (
Discussion
Nephrotoxicity is an important side effect of CsA treatment and constitutes the major detriment of this otherwise remarkable immunosuppressant and anti-rejection drug. CsA-induced renal vasoconstriction is due in part to alterations of arachidonic acid metabolism in favor of the vasoconstrictor TxA2 and to increases in ET-1 expression (Rezzani et al. 2001a; Bianchi et al. 2003).
The vexing issue of CsA nephrotoxicity has prompted clinical and experimental strategies to attenuate this toxicity, particularly the vasoconstrictor component. Approaches such as low-dose dopamine infusion (Sabbatini et al. 1989), use of calcium channel blockers (Rodicio 2000), and omega-3 fatty acid (Torras et al. 1994)-enriched diets to reverse the CsA-induced imbalance between synthesis vasodilator and vasoconstrictor prostaglandins have been employed without apparent benefit in long-term studies. Prostaglandin agonists, such as misoprostol (a PGE analogue; Nast et al. 1995) and TxA2 synthase inhibitors (Kim et al. 1997), are also ineffective.
Semiquantitative analysis of immunohistochemical staining of HO-1, HO-2, and ET-1 in control, CsA, CsA plus SnMP, and CsA plus CoPP-treated rats
Our studies point to HO both as an underlying mechanism of CsA-induced nephrotoxicity and as a target for novel strategies to ameliorate this toxicity. In our model, 21 days of daily CsA treatment produced histopathological lesions, tubule interstitial fibrosis, and lack or swelling of epithelial cell nuclei resembling those seen in kidneys of organ recipients after long-term CsA treatment. In CsA-treated rats, renal HO-1 protein levels and HO renal enzyme activity were reduced. This observation, coupled with the well-established renoprotective effect of HO in various forms of renal injury, i.e., ischemia and inflammation, indicates that CsA depletes the kidney of an important defense system. The beneficial effects of HO activation in renal injury have been attributed to production of heme degradation products, mainly the vasodilator CO and bilirubin-biliverdin, which scavenge reactive oxygen radicals. With respect to CO, its cellular production is regulated by HO. Therefore, the depletion of HO caused by CsA might reduce CO production, thereby allowing action by unopposed vasoconstrictors, such as TxA2 and ET-1, according to the data reported in this work. On the other hand, a reduced production of biliver-din-bilirubin may allow accumulation of reactive oxygen radicals.
Our observations also show that the CsA-induced decrease in HO expression and activity is partial and that it can be restored by established HO inducers such as CoPP. This indicates that attempts to preserve or augment HO activity during administration of CsA treatment is a reasonable strategy to attenuate CsA-induced nephrotoxicity. To this end, augmentation of HO-1 activity would be beneficial by increasing production of CO and biliverdin-bilirubin and by decreasing levels of heme. Reduction of cellular heme levels would reduce activity of heme-containing enzymes such as inducible NOS and TxA2 synthase. The byproduct of inducible NOS, high-output NO production, can be toxic, whereas the byproduct of TxA2 was shown to mediate the CsA-induced constriction of the renal mi-crovasculature.
In summary, our observations demonstrate that down-regulation of HO is a putative mechanism of CsA-induced nephrotoxicity. This effect is partial and reversible. Strategies to preserve or augment HO activity during CsA treatment may attenuate its nephrotoxicity.
With regard to the structures considered targets for CsA nephrotoxicity, we showed that the proximal tubules were mainly involved in these negative effects. These data were confirmed by our previous works (Rezzani et al. 2001a,b) and by the results of other authors (Dekant and Vanvakas 1996) showing that proximal tubules were the first intrarenal targets for most nephrotoxic compounds. Proximal tubules, in fact, were responsible for drug biodegradation and for accumulation of its metabolite (Schaaf et al. 2001). CsA degradation was due, in part, to its oxidation by CYP 450 mixed-function oxidases, which were strongly present in mitochondria of proximal tubules (Burke and Whiting 1986; Pahan et al. 1997; Rezzani et al. 1999). The metabolites obtained from CsA biodegradation in the mitochondria were altered and toxic (Quesniaux et al., 1987). Therefore, we suggest that the metabolites produced alterations in the same organelles with lack of normal morphological cytoarchitecture and then fibrosis in the epithelial cells of proximal tubules, leading to enhanced Na+ reabsorption and thus decreasing fractional excretion rates (Mason 1989).