
Abstract
Leishmania are protozoa that invade mononuclear phagocytes with the involvement of different ligand-receptor systems, including mannose receptors. Until now, scant data are available concerning the mechanisms that govern the infection of Leishmania in other host cell types such as fibroblasts. Our aim was to analyze the expression of mannose receptors in primary cultures of skin fibroblasts (SF) further characterizing their role during the invasion of promastigotes of Leishmania (L.) amazonensis. Both fluorescent, light, and electron microscopy assays revealed that SF have mannose receptors since they bound and internalized mannosylated ligands in addition to being positively labeled by fuc-BSA-FITC probes. D-mannose competition assays revealed the participation of mannose receptors during the parasite association with SF presenting upregulated receptor expression during the initial steps of the infection. After longer periods of Leishmania:fibroblasts contact, the modulation noted in the host mannose receptors was reverted concomitantly to the infection control, suggesting that the parasites were required for the alteration maintenance and providing evidences that the SF may display microbicidal mechanisms to control the Leishmania infection.
T
It is known that the parasites invade phagocytic cells of the vertebrate host with the involvement of different ligand-receptor systems, including the man-nose receptors (MR) (Blackwell et al. 1985; Palatnik-de-Sousa et al. 1993; de Almeida et al. 2003). Inside the phagolysosomal compartment, the promastigotes differentiate to amastigotes and multiply until the lyses of infected cell, releasing large amounts of amastigotes, which can be taken up by other host cells. The life cycle is completed by the bite of the female phle-botomine sand fly when it takes the infecting blood meal containing amastigotes that again differentiate into promastigotes in the insect midgut (Grimaldi Jr. et al. 1983; Mauël 1990; Peters et al. 1995; Sacks and Kamhawi 2001).
The macrophage mannose receptors are C-type lectin, transmembrane glycoproteins (175 kD), which belong to a multilectin receptor protein family displaying eight carbohydrate recognition domains (CRDs) (Wileman et al. 1986; Taylor et al. 1990; East and Isacke 2002). The receptor is expressed on the surface of several cellular types and mediates the binding and internalization of mannosylated glycoproteins (Stahl et al. 1976; Lane et al. 1998; Leteux et al. 2000; Linehan et al. 2000; Lee et al. 2002) as well as participates in the endocytosis of different pathogens enriched with mannose residues at their surface such as Trypanosoma cruzi (Soeiro et al. 1999), Mycobacterium tuberculosis (Noorman et al. 1997), Candida albicans (Stahl 1990) and Leishmania donovani (Wilson and Pearson 1988).
Although most analysis concerning Leishmania invasion has been done using macrophages as host cells due to their high in vivo infection rates, some studies clearly show the infection of other cellular types by the parasites of the Leishmania genus in both in vivo (Zuckerman 1953) as well as in vitro assays (Belle 1958; Lewis 1974), including the infection of cultured human skin fibroblasts (Chang 1978; Schwartzman and Pearson 1985). Since fibroblasts are localized in close proximity to the parasite inoculation area, they can represent a potential cell target for Leishmania early infection and spreading as already observed by others in hamster (Zuckerman 1953) and canine infections (Hervas Rodriguez 1996). Our present aim is to characterize the mannose-receptor expression in primary cultures of skin fibroblasts, examining its possible role in Leishmania (L.) amazonensis infection.
Materials and Methods
Reagents
The following reagents were purchased from Sigma Chemical Co. (St. Louis, MO): horseradish peroxidase (HRP),
Parasites
Leishmania (L.) amazonensis (MHOM/BR/77/LTB0016) isolated from a human case of cutaneous leishmaniasis was used in all experiments and was kindly supplied by Dr. Gabriel Grimaldi, Jr. (Department of Immunology, Fundação Oswaldo Cruz, RJ, Brasil). Amastigote forms were isolated from the lesions of heavily infected animals and maintained in NNN blood agar medium (Novy, McNeal and Nicolle) with an overlay of modified LIT medium (liver infusion-Trypticase) at 25C (Jaffe et al. 1984). The promastigotes obtained on the 7th day of cultivation were harvested by centrifugation at 1500 × g for 10 min at 4C and washed twice in 0.1 M phosphate-buffered saline (PBS) immediately before the experiments.
Primary Cultures of Skin Fibroblasts
Primary cultures of skin fibroblasts (SF) were obtained by the dissociation of mouse skin embryos. The tissues were dissected, minced and incubated for 30 min at 37C in a dissociation solution containing 0.03% collagenase diluted in Dulbecco's modified Eagle medium (DMEM). After three dissociation steps, the enzyme digestion was interrupted by adding 10% fetal bovine serum, the suspension was centrifuged and resuspended in DMEM supplemented with 5% fetal bovine serum, 1 mM
Parasite-Host Cell Infection Assays
Promastigote forms of Leishmania (L.) amazonensis were resuspended in PBS to achieve a ratio of 10:1 parasite/host cell. After 2 hr of interaction at 34C, the SF were rinsed with PBS to remove extracellular parasites, fixed or maintained for 24 hr/34C. In some assays, the infection was followed for 7 days. For light microscopy assays, the SF were fixed with Bouin's solution (75 ml 1.2% aqueous picric acid solution, 25 ml formalin (40% formaldehyde) and 5 ml glacial acetic acid) and stained with Giemsa. The percentage of skin fibroblasts containing associated parasites (parasite association rates) was determined by examining at least 400 randomly selected cells at ×63 magnification under a Zeiss Axioplan microscope (Carl Zeiss Inc., Thornwood, NJ). The randomization permits the evaluation of the whole cover-slips avoiding non-representative analysis of the cell culture infection. All the assays described here were run three to five times at least in duplicate, as described (Soeiro et al. 1999).
Transmission Electron Microscopy Analysis of Mannose Receptors in SF
The expression of mannose receptors in skin fibroblasts was analyzed by two different ultrastructural approaches using the HRP as mannosylated probe (Soeiro et al. 1999). HRP was coupled to 15 or 10 nm colloidal gold particles (HRP-Au) according to Goodman et al. (1979). In the first approach, SF were incubated for 30 min/4C with HRP-Au in the presence of 5 mM calcium chloride, washed to remove unbound ligands and then chased for 24 h/37C. After the incubations, the cultures were fixed for 1 hr/4C with 2.5% glutaraldehyde (GA) in 0.1 M sodium cacodylate buffer and postfixed with 1% osmium tetroxide diluted in the same buffer. The monolayer was then peeled off from the plastic dish in the washing buffer, harvested and centrifuged. The pellet was dehydrated in a graded series of acetone and embedded in Poly/bed 812. In the second approach, the samples were fixed for 30 min/4C with 4% PFA 0.01% GA diluted in 0.1 M sodium cacodylate buffer containing 3.5% sucrose. The monolayer was then peeled off from the plastic dish in the washing buffer, harvested and centrifuged. The pellet was dehydrated in a graded series of methanol and embedded in Lowicryl resin. Sections were hydrated in PBS buffer and incubated for 30 min in 50 mM ammonium chloride. After washing with 0.1 M PBS 3% BSA, the sections were incubated for 10 min in a blocking solution containing 0.1 M PBS 3% BSA 0.2% Tween 20 and then incubated for 1 hr at room temperature with 50 μg/ml HRP-Au. In both protocols, the grids were stained with uranyl acetate and lead citrate and finally observed at Zeiss EM 10C Transmission Electron Microscope. We performed competition assays by incubating the samples with HRP-Au in the presence of 250 mM
The Endocytotic Activity of Mannose Receptors in SF
The ability of skin fibroblasts to incorporate large mannosylated particles was further investigated by using zymosan A particles (Zy) as probe. For light microscopy studies, uninfected cultures were formerly incubated for 10 min-24 hr at 37C with 106 Zy particles, washed and fixed with Bouin's solution followed by staining with Giemsa. The analysis of skin fibroblasts with internalized Zy particles was performed using a Zeiss photomicroscope (Zeiss Inc., Thorn-wood, NY). Fluorescent studies were conducted by staining Zy for 30 min/37C with low doses of Concanavalin-A-TRITC (5 μg/ml), exhaustively washed and then incubated for 10 min and 24 hr/37C with the skin fibroblasts. After the incubations, the SF were washed to remove unbound Con-A-treated Zy and fixed for 20 min/4C with 2% PFA diluted in 0.1 M PBS. The DNA was stained with 10 μg/ml 4,6-diamidino-2-phenylindole (DAPI) to enable the visualization of SF nuclei, the coverslips were mounted over the sections with 2.5% 1.4-diazabicyclo-(2.2.2)-octane (DABCO) and examined immediately using a Zeiss photomicroscope equipped with epifluorescence. All the assays described here were run two to four times at least in duplicate.
D -Mannose Competition Assays
To ascertain the participation of mannose receptors during the Leishmania invasion, SF were incubated for 30 min with increasing doses of
Statistical Analysis
Student's t-test was used to determine whether differences between means in the
Analysis of Mannose Receptors by Neoglycoprotein Binding
To analyze the expression of mannose receptors through fluorescent approaches, skin fibroblasts were allowed to interact for 2, 24, 48 and 72 hr/34C with L. (L.) amazonensis promastigote forms (parasite:host cell ratio of 10:1). After washing, both uninfected and Leishmania-infected SF were immediately fixed for 20 min/4C in 2% PFA. The cultures were then incubated for 1 hr/37C with 50 μg/ml α-L-fucopyranosyl-albumin-FITC (fuc-BSA-FITC), a tracer for mannose receptors (Stahl et al. 1976; Avrameas et al. 1996; Lane et al. 1998). After washing with PBS, the uninfected and Leishmania-infected cultures were further incubated with DAPI for visualization of SF and parasites nuclei and kinetoplast of the parasites, mounted with DABCO and examined as described above. In all experiments, optimal lectin or neoglycoprotein concentrations were determined by preliminary titration assays. To assure labeling specificity, some assays were performed with the addition of 250 mM
Results
The Expression of Mannose Receptors in SF and Their Endocytotic Activity
The expression of mannose receptors in skin fibroblasts was initially investigated by fluorescent microscopy assays using related neoglycoproteins. Our data showed that fuc-BSA-FITC bound to the mannose receptors present in skin fibroblasts, which displayed a fluorescent labeling throughout the cell surface (Figures 3A and 3B). The expression of mannose receptors in skin fibroblasts was further characterized by following the binding and internalization of HRP-Au particles, another well-known mannosylated ligand (Shepherd et al. 1994; Marzolo et al. 1999; Soeiro et al. 1999) by transmission electron microscopy. The tracer could be localized at the surface of SF after their incubation for 30 min/4C (Figure 1A) as well as during the labeling of the ultrathin sections with HRP-Au (Figure 1B). Following the internalization at 37C, we found the tracer both within intracellular vesicles in close proximity to the cell surface (Figure 1C inset) and inside perinuclear endosomes, which displayed different sizes and electrondensity (Figure 1C). The competition assays performed by the addition of
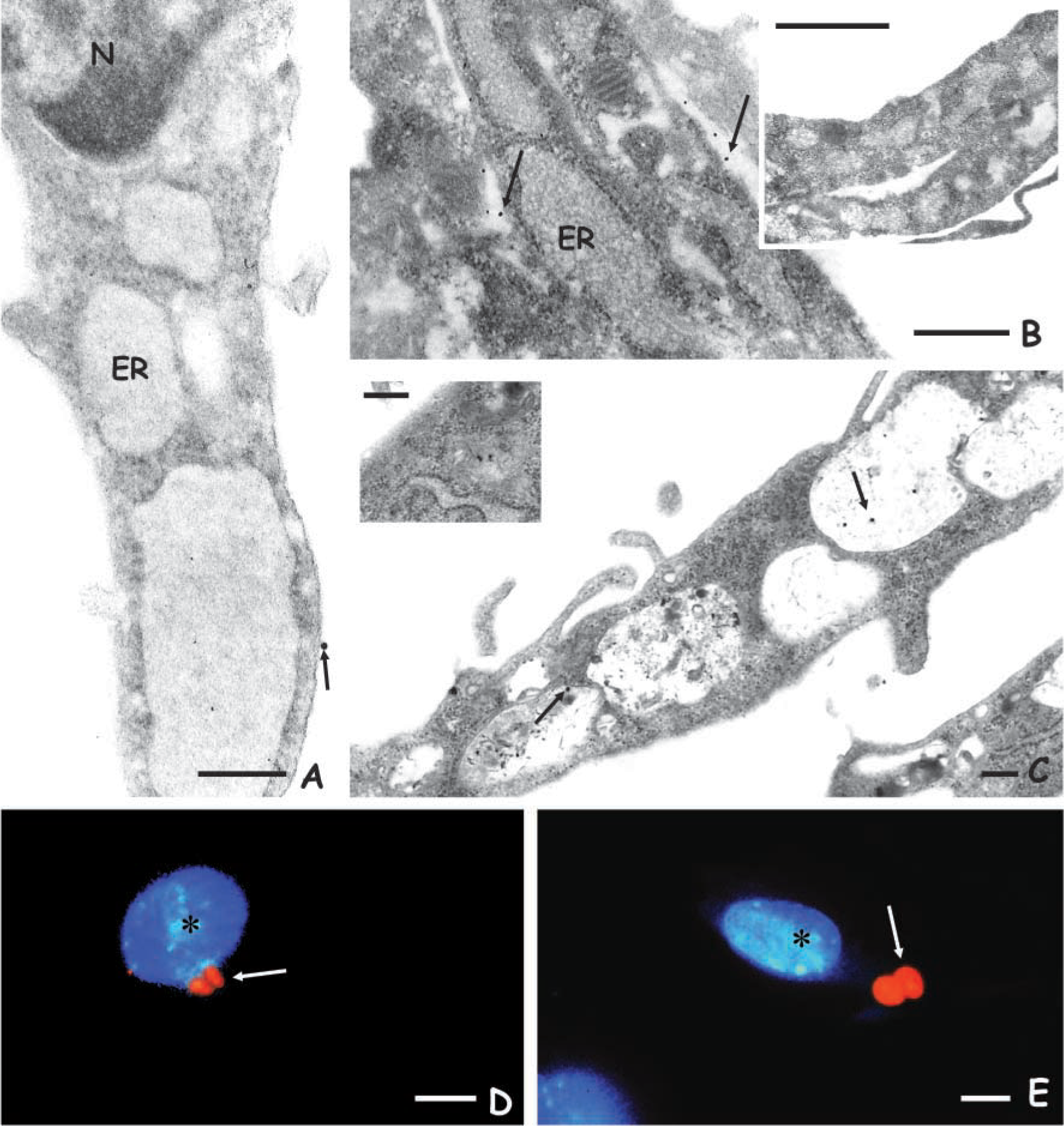
Transmission electron microscopy analysis showing mannose receptors at the surface of skin fibroblasts during incubation with HRP-Au both at 4C (
The Role of Mannose Receptors during the Invasion of Skin Fibroblasts by Promastigotes of L. (L.) amazonensis
To investigate the involvement of mannose receptors during the interaction of SF with L. (L.) amazonensis, competition assays were done employing different doses of
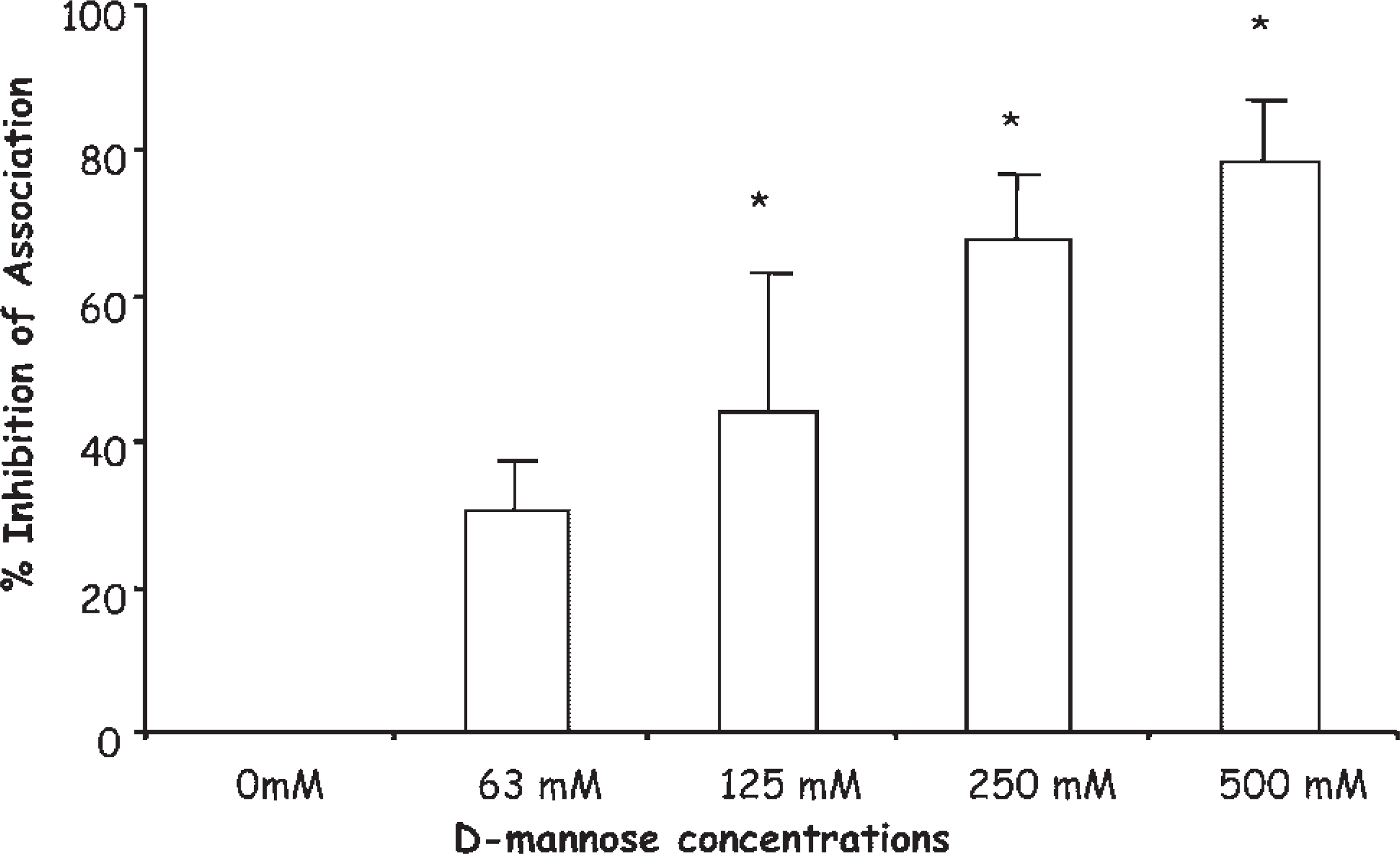
Competition assays showing the participation of man-nose receptors during the association of promastigotes with skin fibroblasts. The assays were realized by adding increasing doses of
The Mannose Receptors Are Regulated during the Infection of SF by Promastigotes of L (L.) amazonensis
Finally, we analyzed the expression of mannose receptors in skin fibroblasts infected by Leishmania. Our results using fuc-BSA-FITC probes showed that after interaction for 2–48 hr with promastigote forms, a higher expression of mannose receptors was noted both at the surface of the infected SF as well as within intracellular vesicles (Figures 3C, 3E and 3G) as compared with uninfected cells (Figure 3A). The addition of 250 mM
Discussion
To successfully invade the host cells, the promastigotes of Leishmania species must first interact with molecules present at the surface of the host cells both directly and/or indirectly through their opsonization by host soluble molecules (Guy and Belosevic 1993; Kelleher et al. 1995; Handman and Bullen 2002). In vitro data described the participation of different receptor-ligand systems during the internalization of promastigotes by macrophages, including 1. mannose receptors (Wilson and Pearson 1986, 1988); 2. complement receptors type 1 (CR1) and type 3 (CR3) (Da Silva et al. 1989; Russell and Talamas-Rohana 1989; Kedzierski et al. 2004), receptor for advanced glycosylation end products (Mosser et al. 1987); and 3. receptors for extracellular matrix molecules, including fibronectin (Rizvi et al. 1988; Russell and Talamas-Rohana 1989) and heparan sulfate (Butcher et al. 1992).
Although macrophages are considered the major host cells for Leishmania infection in vivo, the infection of other phagocytes has been reported both in vivo as well as in vitro, including neutrophils (Laskay et al. 2003) and dendritic cells (Colmenares et al. 2004; Prina et al. 2004). Regarding the infection of non-professional phagocytic cells, Belle (1958) described the infection of epithelial cells by L. (L.) donovani in vitro and later Lewis (1974) demonstrated the ability of L. (L.) mexicana parasites to invade MDCK epithelial cells. Likewise, the infection of fibroblasts by Leishmania species has been reported both in in vivo (Zuckerman 1953; Hervas Rodriguez 1996; Tarantino et al. 2001) and in vitro studies (Chang 1978; Schwartzman and Pearson 1985; Côrte-Real et al. 1995).
The MR are expressed on the surface of several cellular types, including mononuclear phagocytes, especially tissue macrophages (Shepherd et al. 1982; Basu et al. 1991), retina epithelial cells (Lane et al. 1998), dendritic and Langerhans cells (Noorman et al. 1997), tracheal smooth muscle cells, kidney mesangial cells and Kaposi sarcoma cells (reviewed in Leteux et al. 2000), fibroblasts (Straus 1983) and cardiomyocytes (Soeiro et al. 1999). It has been implicated in clearing glycoconjugates ending in mannose,
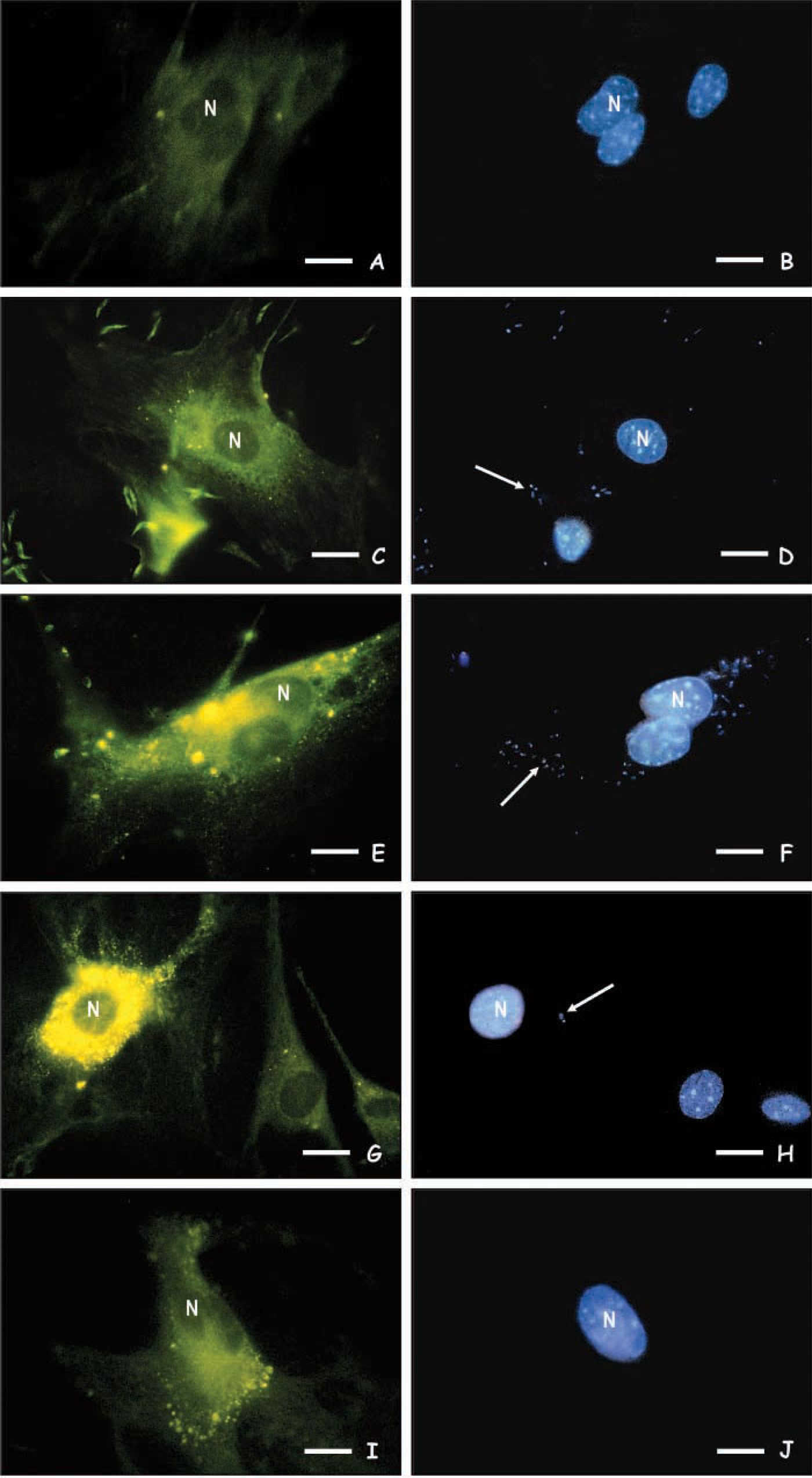
Fluorescence microscopy analysis of mannose receptors expression in uninfected SF (
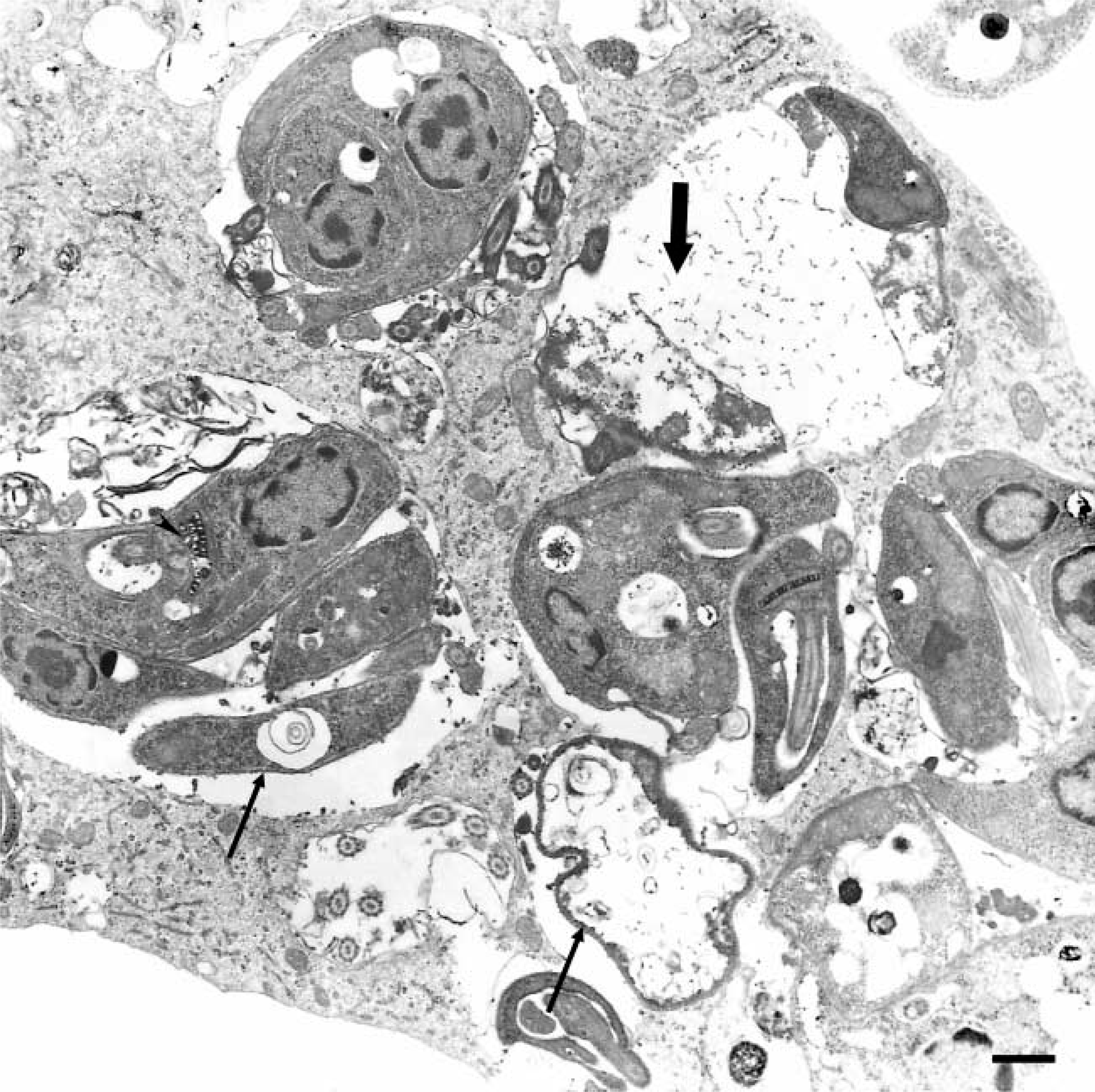
Transmission electron microscopy image showing SF after 72 hr of interaction with promastigotes of L. (L.) amazonensis presenting non-viable parasites (arrow) and parasite debris (large arrow) within large parasitophorous vacuoles. Note the alterations in the Leishmania kinetoplast (arrowhead). Bar = 0.5 μm.
In the present work, we characterized the expression of MR in primary cultures of skin fibroblasts evaluating its role during the invasion by L. (L.) amazonensis promastigotes. Our data revealed that uninfected skin fibroblasts express mannosyl binding sites because 1. they bound a neoglycoprotein that specifically bind to mannose receptors (Avrameas et al. 1996); 2. they are able to internalize zymosan A, which are mannosylated particles frequently used in phagocytic assays (Speert and Silverstein 1985; Lombard et al. 1994); and 3. ultrastructural assays evidenced the association of HRP-Au to the SF surface, another mannosylated ligand commonly used for MR analysis (Shepherd et al. 1994; Marzolo et al. 1999; Soeiro et al. 2002). The addition of d-mannose efficiently blocked the binding of both HRP-Au and α-
After characterizing the MR in skin fibroblasts, we next evaluated its role during the invasion of L. (L.) amazonensis promastigotes. Competition experiments conducted by the addition of increasing concentrations of
Finally, as 1. the modulation of MR has been described in host cells infected with various microorganisms, including Bacillus Calmette-Guérin (Ezekowitz and Gordon 1982), L. (L.) donovani (Basu et al. 1991), Candida albicans (Shepherd et al. 1997; Gelderman et al. 1998), Pneumocystis carinii (Ezekowitz et al. 1991), Trypanosoma cruzi (Soeiro et al. 1999), and 2. our present data pointed to the participation of MR during the parasite invasion, and our next aim was to analyze the expression of MR in Leishmania-infected SF. Our results showed a gradual increase in the expression of MR in infected SF, reaching maximal expression after 48 hr of infection, with the labeling localized mostly at the cellular surface as well as intracellularly. The reversion of the up-regulation was evidenced after 72 hr of infection concomitantly to the presence of non-viable parasites and cellular debris visualized and confirmed by transmission electron microscopy, which was suggestive of abortive infection of L. (L.) amazonensis in SF. The mechanisms that govern the MR modulation in Leishmania-infected skin fibroblasts were not presently characterized, but some possibilities can be envisaged including alterations in the receptor degradation levels and/or transportation of recycled receptors (and/or newly synthesized) for the cellular surface and/or reduced MR synthesis mediated by the parasite and/or host-infected-cell factors. Our results demonstrated that the modulation in the expression of host mannose receptors induced by Leishmania invasion was reverted concomitantly to the loss of parasite viability, suggesting that the presence of viable parasites is required for the alteration maintenance noted in the MR.
The modulation of MR has been reported during other parasitic infections (Ezekowitz and Gordon 1982; Basu et al. 1991; Shepherd et al. 1997; Gelderman et al. 1998; Soeiro et al. 1999). Then the modulation of MR during the invasion of SF by Leishmania promastigotes could be expected by analogy with MR modulation by Bacillus Calmette-Guérin (Ezekowitz and Gordon, 1982), L. (L.) donovani (Basu et al. 1991), Candida albicans (Shepherd et al. 1997; Gelderman et al. 1998) and T. cruzi (Soeiro et al. 1999). At the present time, assays are underway to better investigate the factors involved in this up-regulation.
In summary, our present results pointed to the role of mannose receptor during the interaction of L. (L.) amazonensis promastigotes and skin fibroblasts in a similar way to the already reported system occurring during the invasion of macrophages by Leishmania promastigotes.
Footnotes
Acknowledgements
We thank Bruno Ãvila, Genézio Lopes Faria and José L. Faria for their excellent technical support. CNPq, FAPERJ, Fundação Oswaldo Cruz (FIOCRUZ) supported this investigation.