
Abstract
Central nervous system nuclei and circuits, such as the medial preoptic, ventromedial and paraventricular nuclei of the hypothalamus, play important roles in reproduction and parturition, and are influenced by estrogen. Peripheral autonomic and sensory neurons also play important roles in pregnancy and parturition. Moreover, the steroid hormone estrogen acts directly, not only on the reproductive tract organs (uterus and cervix), but also on the central and peripheral nerves by regulating expression of various neuronal genes. The peripheral primary afferent neurons innervating the uterine cervix relay mechanical and biochemical sensory information induced by local cervical events and by passage of fetuses, to the spinal cord and supraspinal centers. Consequently, the birth process in mammals is influenced by the combined action of neurons and hormones. Peripheral sensory stimuli, induced physiologically by fetal expulsion or mechanically by vaginocervical stimulation, alter behavior, as well as autonomic and neuroendocrine systems. Recent evidence indicates that primary afferent neurons innervating the cervix, in addition to their sensory effects, likely exert local “efferent” actions on the ripening cervix near term. These efferent effects may involve estrogen-regulated production of such neuropeptides as substance P and calcitonin gene-related peptide in lumbosacral dorsal root ganglia, and their release in the cervix. Collectively, these findings suggest an interrelationship among estrogen, cervix-related sensory neurons, and local cervical events near term.
Keywords
C
Until recently, little was known about the action of sensory neurons in parturition or estrogenic effects on peripheral sensory neurons innervating the female reproductive tract, even though a role for sex steroids in the peripheral neurons was proposed over seven decades ago (Nissen 1929). However, two sets of studies helped to direct research in this area. First, the remarkable studies in the early 1970s by Komisaruk et al. (1972) and Kow and Pfaff (1973), which demonstrated enlargement of peripheral sensory fields of genital nerves by 17β-estradiol in ovariectomized rats. Second, the studies by Carlson and De Feo (1965), Higuchi et al. (1987), and Burden et al. (1990a, b) showed that pelvic neurectomy (especially the sensory nerve) interfered with parturition. Subsequently, interest in this field has been growing steadily. We now know that estrogen exerts a broad range of biological effects on sensory neurons related to reproductive organs via ERs (Papka et al. 1997,2000,2001b; Papka and Storey-Workley 2002; Mowa et al. 2003a,b).
The “efferent” effects of primary afferent neurons on peripheral target organs is a well-established concept in tissues such as the skin and lungs, but relatively new in the female reproductive organs (reviewed by Holzer 1988; Maggi and Meli, 1988; also see Collins et al. 2002 and Mowa et al. 2003a, b, c). The present review provides an update on the relationship between estrogen and reproduction-related peripheral sensory neurons of the female, including (1) the distribution pattern of ERs and neuropeptides in reproductive organ-related peripheral sensory neurons during pregnancy and parturition, and (2) the influence of estrogen on neuropeptide synthesis and the effects of neuropeptides on local peripheral tissue changes, in particular the cervical ripening process. These observations point to an interrelationship among (1) the ER and estrogen system, (2) the uterine cervix-related sensory neurons of the lumbosacral (L6-S1) dorsal root ganglia (DRG), and (3) parturition.
Estrogen Receptors
Biology of Estrogen Receptors
Estrogen exerts its influence on target cells, including neurons, by acting principally through two ER subtypes, ER-α, the “classical” subtype cloned in 1986 (Koike et al. 1987) and ER-β, a more-recently cloned subtype (Kuiper et al. 1996). Various isoforms of ER-β have since been reported, namely ER-β2, ER-βcx, and ER-β variants that are altered by a deletion within the DNA-binding domain (ER-β1δ3 and ER-β283) (Moore et al. 1998; Ogawa et al. 1998).
ERs are members of the steroid/retinoid receptor superfamily that have structural and functional similarities (Mangelsdorf et al. 1995) and are divided into four structurally distinct, functional domains, namely (Figure 1): the N-terminal A/B region, which contains the activation function 1 (AF-1) and is responsible for transcriptional activation; the DNA-binding domain, which mediates specific DNA binding and is also called the mid- or C-region; the less-characterized hinge region or D domain; and the C terminus- or ligand-binding domain, which contains the ligand-binding pocket and sites for cofactor-binding, transactivation (AF-2), nuclear localization, and interactions with heat shock proteins (for review, see Weihua et al. 2003). When estrogen binds to its receptors, the ERs dimerize and regulate transcription by binding, in most cases, to the classical consensus estrogen response elements in the promoter region of the target genes, leading to a cellular response (Figure 2) (for review, see Weihua et al. 2003). ERs also can induce transcription by binding to AP-1 and Sp-1, where ER-α and ER-β have different and, in some cases, opposing effects (Gaub et al. 1990; Philips et al. 1993; Umayahara et al. 1994; Webb et al. 1995). Rapid effects of estrogen on neurons are exerted via the recently identified, but less characterized, membrane ER (McEwen 1991), which may be identified as ER-X (Toran-Allerand et al. 2002).
Peripheral Sensory Nerves of the Uterine Cervix
Sensory Neurons of the Lumbosacral Dorsal Root Ganglia
Sensory, sympathetic, and parasympathetic fibers are present in the uterus and cervix (Inyama et al. 1986; Traurig et al. 1988; Papka and Traurig 1993; Papka et al. 1995; Collins et al. 1999,2002) (Figures 3A–3D). A large segment of the sensory neural component of the cervix is supplied by the pelvic nerves, arising from the lumbosacral (L6-S1) DRG and entering corresponding segments of the spinal cord (Inyama et al. 1986; Berkley et al. 1988; Papka 1990), with additional inputs by the hypogastric and vagus nerves, arising rostrally from the T13-L4 and directly from the brainstem via the nodose ganglia, respectively (Berkley et al. 1988; Collins et al. 1999). The sensory neurons from L6-S1 DRG also innervate the urinary bladder and vagina, and distal gastrointestinal tract (Inyama et al. 1986; Nadelhaft and Vera 2001; Bennett et al. 2003).
Neurochemistry
The sensory neurons of the DRG synthesize multiple neurotransmitters, including the neuropeptides calcitonin gene-related peptide (CGRP), substance P (SP), secretoneurin (SN), neurokinin A, cholecystokinin, somatostatin, brain-derived nerve factor (BDNF), and pituitary adenylate cyclase-activating polypeptide (PACAP) (Papka and Traurig 1993; Yang et al. 1998; Collins et al. 1999,2002; Mowa et al. 2003a, b). SP, SN, and CGRP are of particular interest in the study of the birth process in that they: (1) are richly expressed in unmyelinated, small, capsaicin-sensitive sensory neurons, which have terminals in the reproductive tracts, including the cervix (Traurig et al. 1988; Papka et al. 1995; Collins et al. 2000,2002) (Figures 3A–3D); (2) coexist in neurons of the L6-S1 dorsal root ganglia (Ju et al. 1987); (3) are potent vasoactive neuropeptides with cooperative activities (Gamse and Saria 1985; Gamse et al. 1987; Green et al. 1992); and (4) have receptors in the cervix and are (peptides) released locally in the cervix (Collins et al. 2002; Pokabla et al. 2002; Mowa et al. 2003a, b). An established concept is that afferent nerves also have peripheral efferent effects when releasing transmitters from their terminals in target tissues (Holzer 1988; Maggi and Meli 1988; McDonald et al. 1996; Szolcsanyi 1996) (Figures 3A–3D).
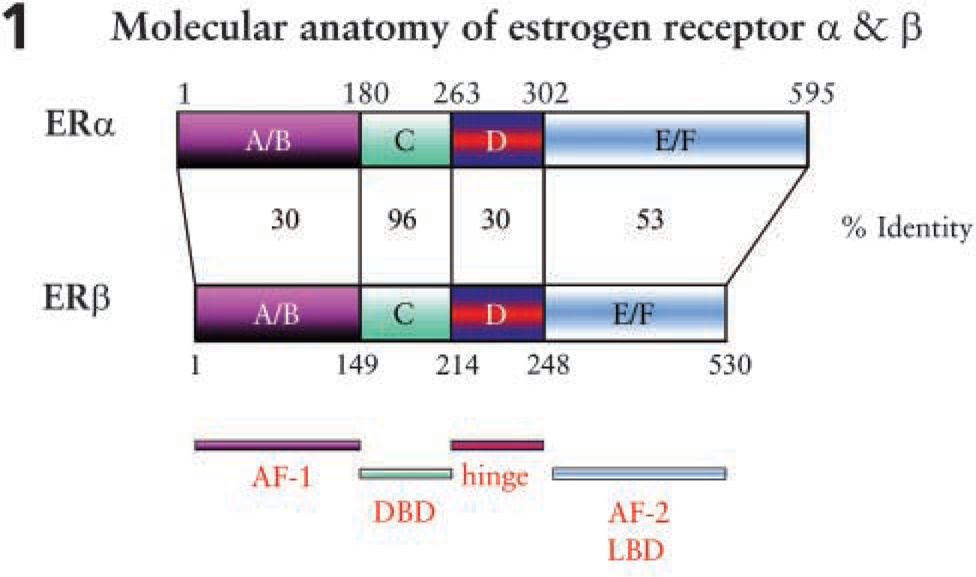
Model illustrating the molecular anatomy of estrogen receptor (ER) subtypes
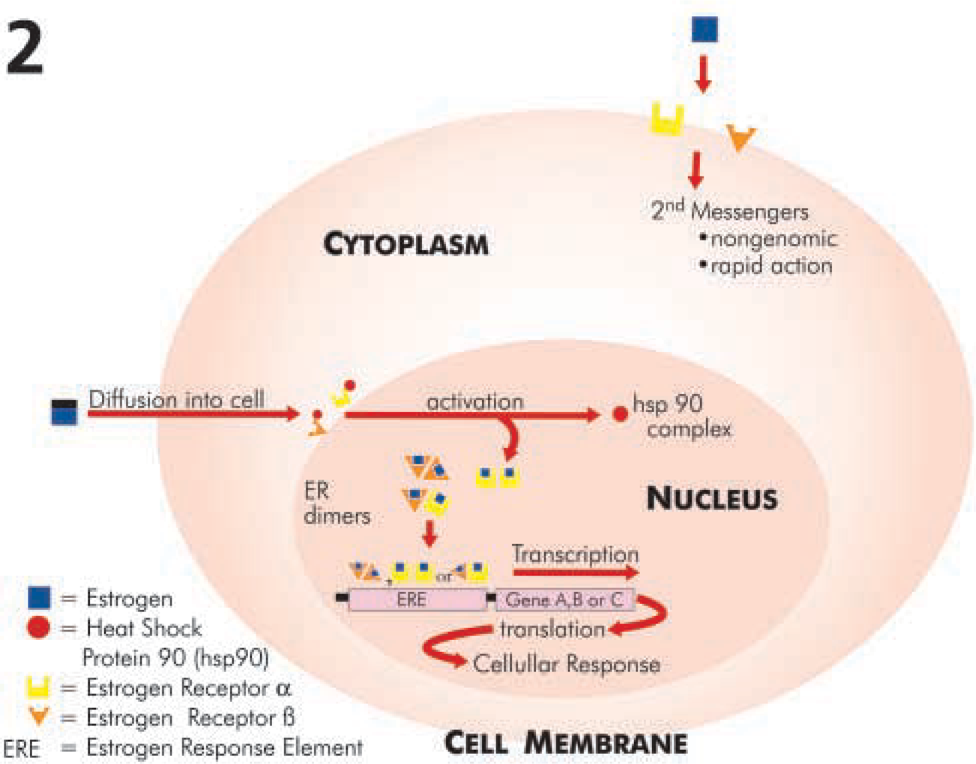
Schematic diagram of the ER and its mechanism of action in a model cell such as a sensory neuron. Estrogen diffuses into the cell and binds to the ER to initiate ER activation and dimerization (homo- and heterodimers). The ligand receptor complex in turn binds to the estrogen response elements on the promoter of the DNA to initiate transcription, translation, and, ultimately, cellular response. Rapid response to estrogen may be mediated by the recently described membrane ER (ER-X).
Estrogen Receptors in the Sensory Neurons of the Female Reproductive Organs
Distribution. The findings by Komisaruk et al. (1972) and Kow and Pfaff (1973) that 17β-estradiol treatment of ovariectomized rats led to the enlargement of the peripheral sensory fields of genital nerves served as the impetus for a series of studies. The initial studies were aimed at characterizing reproductive-related sensory neurons and their expression of ERs using several techniques, including retrograde axonal tracing (Figures 3E and 3F), immunohistochemistry (Figure 3G), in situ hybridization, and RT-PCR (Figures 4 and 5) (Sohrabji et al. 1994; Collins et al. 1999,2002; Taleghany et al. 1999; Papka et al. 2000; Shinohara et al. 2000; Papka and Storey-Workley 2002; Mowa et al. 2003a, b). Both ER-α and ER-β mRNA and protein are expressed at significant levels in a subpopulation of small- and medium-sized neurons of lumbosacral (L6-S1) DRG in both postnatal and adult animals, making them important targets of estrogen action (Figure 3G) (Sohrabji et al. 1994; Papka et al. 1997,2000,2001b; Taleghany et al. 1999; Shinohara et al. 2000; Papka and Storey-Workley 2002).
Estrogen Receptors in Neurochemically Identified Sensory Neurons of L6-S1 DRG
Many of the small- and medium-sized neurons represent visceral afferent and nociceptive neurons (Figures 3A–3D). Many of these neurons are responsive to capsaicin, a sensory neurotoxin, and express the capsaicin receptor, vanilloid receptor-1 (VR-1) (Figures 3H and 3I) (Papka et al. 2001a; Bennett et al. 2003); others express the P2 × 3 receptor and may respond to ATP (Figures 3J and 3K). A subpopulation of the small- and medium-sized DRG neurons innervating the female reproductive organs, ~30%, express either ER-α or ER-β, or both ER-α and ER-β, indicating the potential complexity and variability of estrogen action in these cells (Papka et al. 1997,2001b; Papka and Storey-Workley 2002; Bennett et al. 2003). These ER-expressing neurons can be further characterized into different subsets or subgroups on the basis of their neurochemical phenotype, an indicator of their likely function or the type(s) of stimuli to which they are likely to respond (Mowa et al. 2003a, b). The neurochemicals (and their receptors) coexpressed with ER in L6-S1 DRG neurons include SP, CGRP (Mowa et al. 2003a, b; Papka and Mowa 2003), BDNF, nitric oxide synthase, PACAP, and SN (Figures 3L and 3M). In addition, ER is coexpressed with the neurokinin 1 receptor (SP receptors) (Papka and Mowa 2003), VR-1 (Figures 3H and 3I), and P2 × 3 receptors (Figures 3J and 3K). These neurons innervate the cervix and could transmit sensory information, such as the noxious and mechanical stimuli producing neuroendocrine reflexes, from the cervix as well as the urinary bladder (Inyama et al. 1986; Papka et al. 1995; Nadelhaft and Vera 2001; Nadelhaft et al. 2002), and, importantly, the presence of ERs makes them a likely target of estrogen action. However, not all neurons of the L6-S1 DRG have been characterized yet, based on their coexpression of ER with local neurochemicals. These include neurons containing neuropeptides such as galanin, cholecystokinin, and somatostatin.
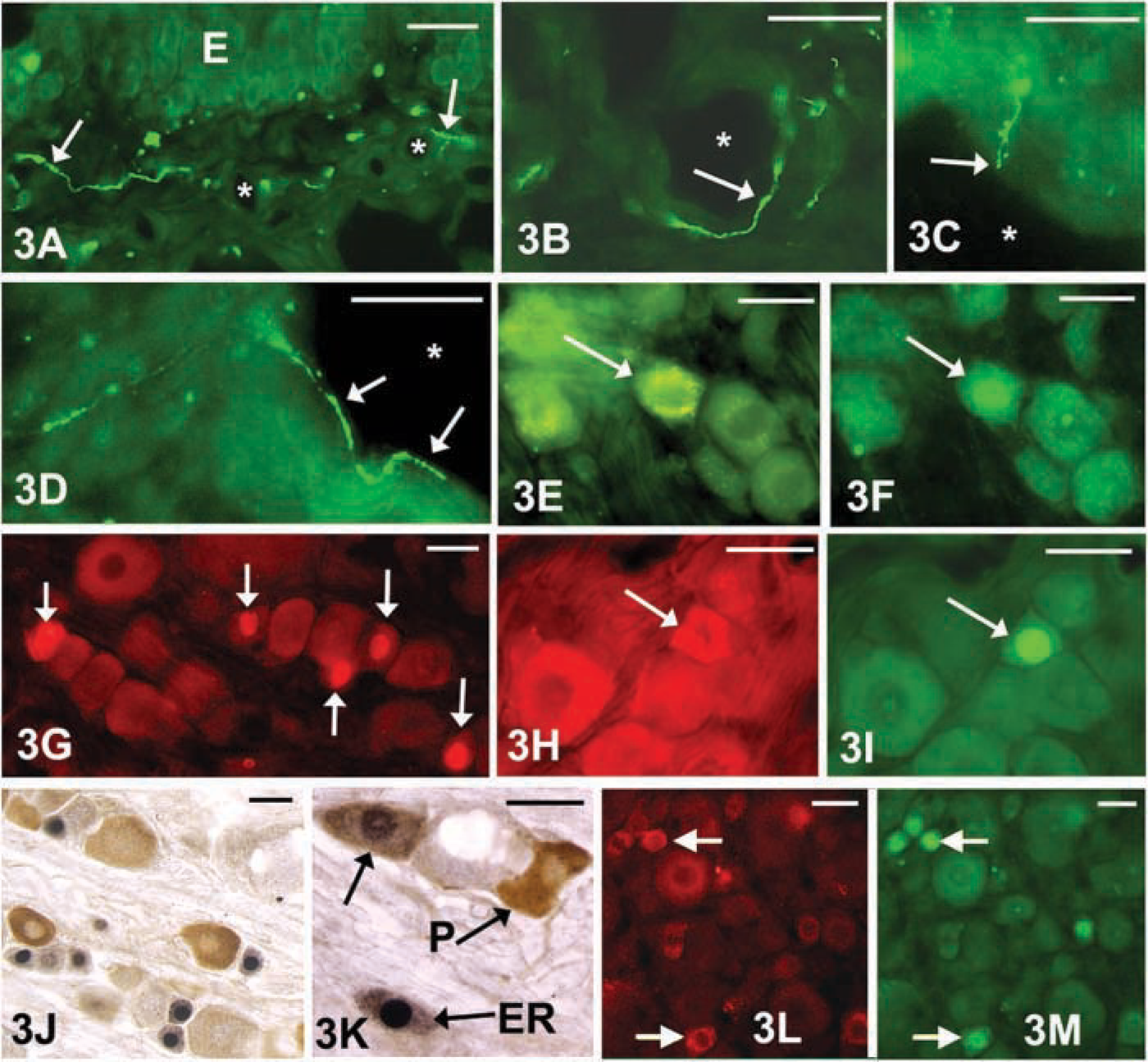
Photomicrographs of the uterine cervix (
Estrogen Modulates Synthesis of Neuropeptides in Sensory Neurons of Female Reproductive Organs
17β-Estradiol administered to ovariectomized rats in the short term (4 days) upregulates synthesis of neuropeptides such as SP, SN, and CGRP in a dose-related manner (Gangula et al. 2000; Mowa et al. 2003a, b; Mowa et al., unpublished data). This effect is reversed by pretreatment with the ER blocker, ICI 182 780 (Mowa et al. 2003a, b) (Figures 4 and 5). Such a result indicates that in sensory neurons in locales that innervate female pelvic organs, estrogen acts via its receptors to modulate synthesis of these neuropeptides (Figures 4 and 5). On the other hand, it appears that long-term treatment (90 days) with estrogen (Premarin) downregulated β-preprotachykinin (SP) mRNA (Liuzzi et al. 1999), implying that there may be several mechanisms through which estrogen acts, depending on the physiological conditions and duration of exposure to estrogen. The functional significance of the various subpopulations of L6-S1 DRG sensory neurons, based on the type of ER isoform expressed, and their regulation in pregnancy is as yet unclear: some neurons lack ERs, whereas others express only ER-α or ER-β, and others both ER-α and ER-β (Papka et al. 1997,2001b; Papka and Storey-Workley 2002; Bennett et al. 2003). Moreover, there is evidence that long-term treatment (21–28 days) of ovariectomized rats with exogenous estrogen (17β-estradiol) downregulates ER-α but upregulates ER-β (Taleghany et al. 1999). On the other hand, under the physiological conditions of pregnancy, levels of ER-β mRNA in the DRG remain largely unchanged throughout the course of pregnancy, whereas, in contrast, those of ER-α rise near term, in parallel with those of plasma estrogen and the neuropeptides SP and CGRP (Mowa et al. 2003a, b) (Figure 4). Thus, ER-α may play a dominant role in reproductive processes and during pregnancy, including mediating neuropeptide synthesis (Mowa et al. 2003a, b) (Figure 4).
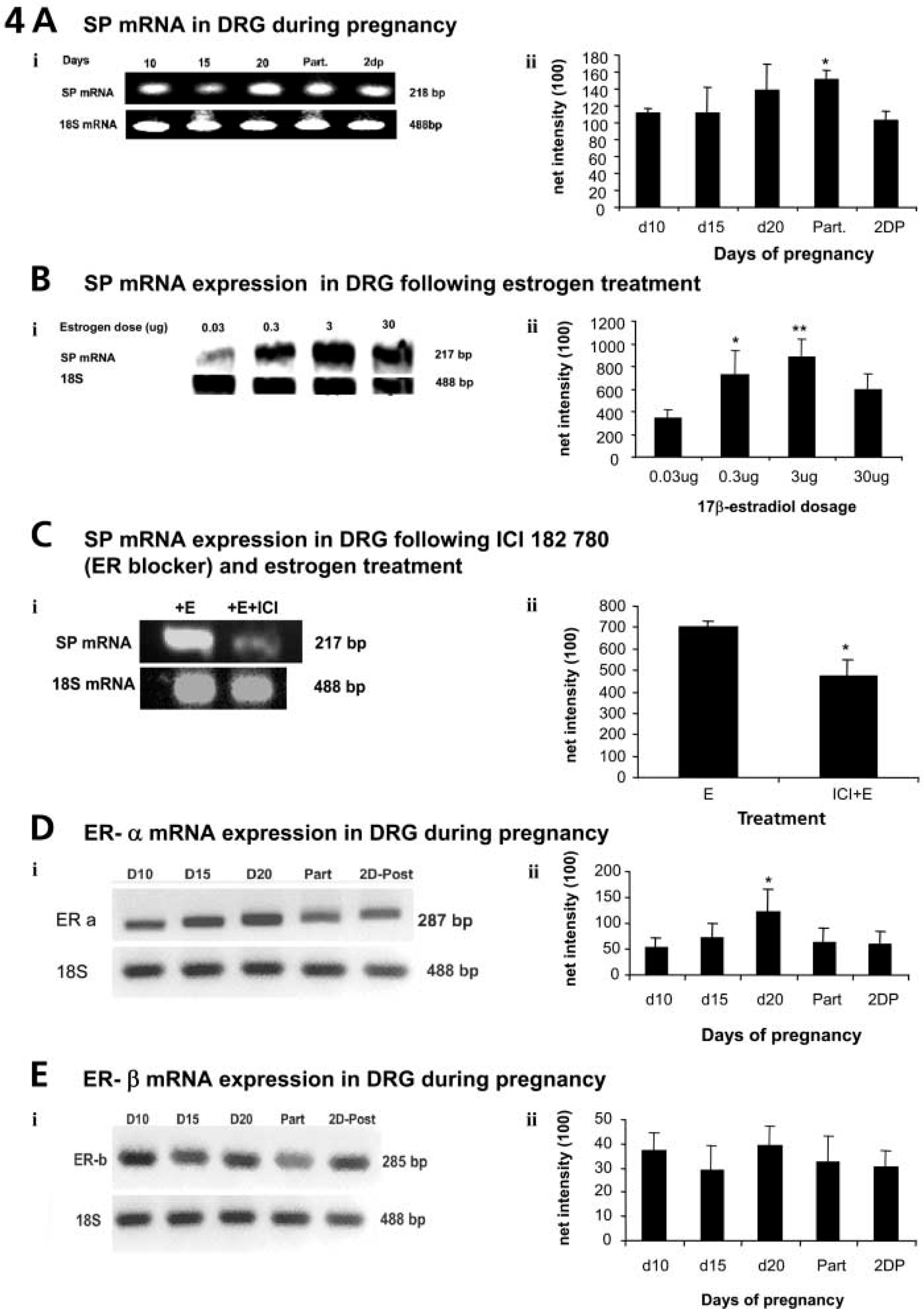
Data from RT-PCR analysis showing changes in SP, ER-α, and ER-β mRNAs in L6-S1 DRG at representative time points during pregnancy, the effect of estrogen treatment, and the effect of an ER blocker. SP (
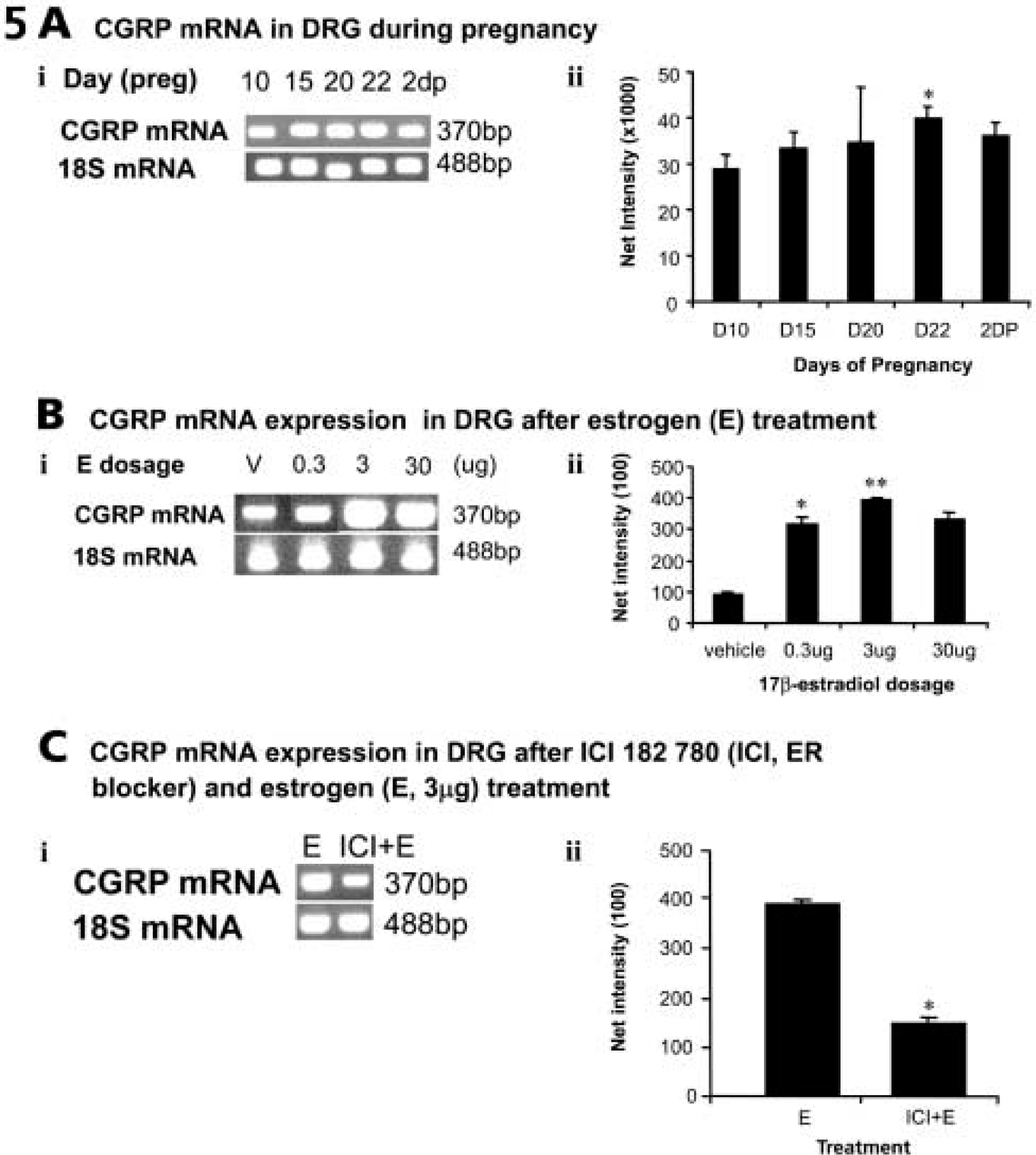
Data from RT-PCR analysis showing changes in CGRP mRNAs in L6-S1 DRG at representative time points during pregnancy (
Potential Interrelationship among Estrogen, Sensory Neurons, and Cervical Ripening
The Cervix: Basic Composition and Remodeling (Cervical Ripening)
The cervix forms the lower and narrower component of the uterus (Figure 6) and functions during pregnancy as a mechanical sphincter. The stromal matrix of the cervix is composed of structural proteins, including types I and II collagen and elastin, as well as proteoglycans and hyaluronan (Kelly 2002). During pregnancy, the quiescent uterus provides a favorable environment for the growing fetus, and the nonpliable cervix ensures retention of the fetus in utero. However, at term, the cervix undergoes tissue remodeling or cervical ripening, a progressive dissociation and disorganization of collagen fibers and bundles, leading to softening, dilatation, and loss of sphincteric function, whereas the uterus contracts to facilitate the timely passage of the fetus (Kelly 2002). A failure of either or both of these events leads to birthing problems such as preterm birth or protracted labor (Challis 2000).
Cervical ripening is complex and is regulated by numerous factors, many of which have been studied extensively, such as nitric oxide, cytokines, prostaglandin, relaxin, and steroid hormones (Taniguchi et al. 1991; Cunningham et al. 1993; Matsuzaki et al. 1993; Ali et al. 1997; Arntzen et al. 1998). However, the exact mechanism(s) that underlie the initiation and progression of cervical ripening remain elusive, implying that there may be other, as yet unidentified, factors involved in parturition. Indeed, data pointing to a role for sensory nerves in cervical ripening are strengthening.
Sensory Neurons and Parturition
The uterus and cervix are richly innervated by fibers of the pelvic nerves (Figures 3A–3D). The sensory fiber component of the pelvic nerves is critical for parturition; bilateral dorsal rhizotomy of the L6-S1 roots at days 8–10 of pregnancy induces dystocia, or difficult birth, whereas bilateral transection of ventral roots does not (Carlson and De Feo 1965; Higuchi et al. 1987; Burden et al. 1990b, b; Martinez-Gomez et al. 1998). Because the uterus, unlike the cervix, undergoes a physiological loss of innervation over the course of pregnancy (Haase et al. 1997), it is reasonable to state that the effect of bilateral pelvic neurectomy on parturition, as described above, could be attributed to the disruption of innervation to the cervix. Moreover, preliminary immunohistochemical data from our lab indicate that the neuronal activity of cervix-related sensory neurons increases during pregnancy, as indicated by the increase in expression of phosphorylated cyclic-AMP response element-binding protein immunoreactivity in Fluorogold retrogradely labeled sensory neurons in L6-S1 DRG (Figure 6) (Figures 7A and 7B). Interestingly, a trend toward peak activity in cervix-related sensory neurons at day 20 of pregnancy roughly coincides with the onset of cervical ripening at day 19.5.
The specific neuronal factors and exact mechanism(s) underlying the effects on the birth process of sensory nerve activity or its loss following bilateral dorsal rhizotomy of the L6-S1 roots are not yet completely clear. Recent evidence implicates the mediation of vasoactive and inflammation-inducing neuropeptides, such as SP, CGRP, and SN (Collins et al. 2000,2002; Mowa et al., 2003a, b). Indeed, the cervix is densely populated by nerve terminals immunoreactive to SP, CGRP, and SN and their respective receptors (at least those of SP and CGRP) (Collins et al., 2002; Pokabla et al. 2002), making the cervix a significant target of SP and CGRP action. Importantly, the inflammatory-like changes that occur during the cervical ripening process are strikingly similar to neurogenic inflammation induced by SP and CGRP in other organs and tissues (Liggins 1981; McDonald et al. 1996; Richardson and Vasko 2002). Moreover, there is a reduction in SP and CGRP in the cervix at about day 20 of pregnancy in the rat (which could be interpreted as increased release of SP and CGRP in late pregnancy), and administration of SP to nonpregnant rats replicates inflammatory-like changes evident in the ripening cervix (Collins et al. 2002; Mowa et al. 2003a, b). Taken together, these data implicate SP and CGRP as sensory neuronal factors or active “ingredients” that may mediate the effects of sensory neurons in the ripening cervix. Consequently, the dystocia, or difficult birth, induced by bilateral transection of pelvic nerves is, in part, likely to arise because of the interruption of the activity of these nerves and the resulting effects on SP and CGRP. Administration to pregnant animals of receptor blockers for SP and CGRP will help delineate their specific effect on cervical ripening.
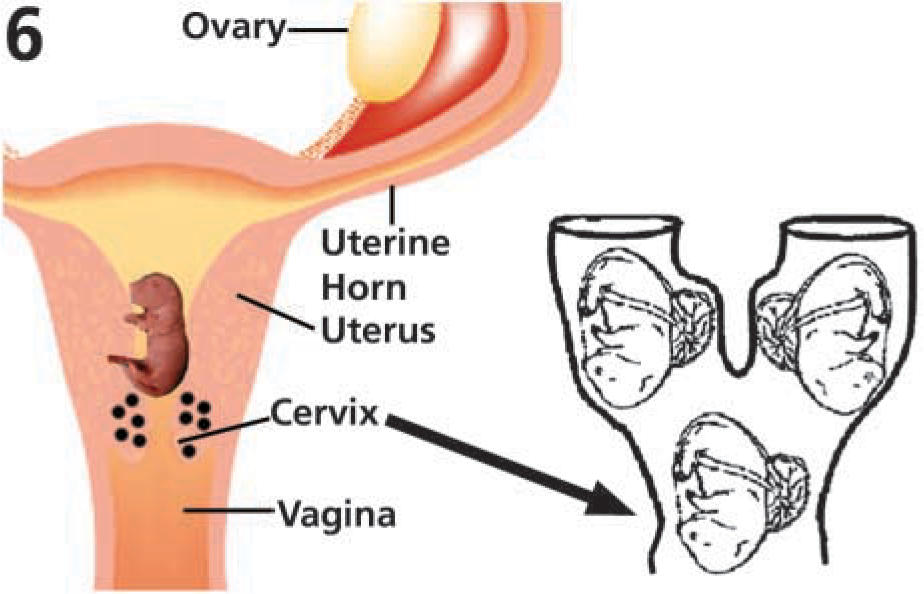
Schematic diagram of the female reproductive system (on the left) showing its major components: ovary, uterine horns, uterus, cervix, and vagina. The collagen-rich cervix (indicated by black dots) forms the lower and narrower component of the uterus and functions as a mechanical sphincter during pregnancy. The cervix dilates and softens by remodeling in late pregnancy to allow passage of the fetus at term (arrow to diagram on the right).
Sensory Neuronal Factors Induce Cervical Microvascular Remodeling
The exact mechanism(s) underlying the inflammatory-like effects of SP and CGRP on cervical remodeling are, as yet, unknown. However, the close association of SP and CGRP immunoreactive nerve terminals (Figures 3A–3D) and postcapillary venules of the cervix, and the localization of their receptors to the endothelia of these vessels (Collins et al. 2002; Pokabla et al. 2002) suggest that the SP/CGRP-induced inflammatory-like responses may be mediated by the microvas-culature. Indeed, SP administered to virgin rats invokes vasodilation and vascular leakage, leading to leukocyte egress and plasma extravasation in the cervix similar to those seen in parturient rats (Collins et al. 2002). These changes in the microvasculature may be essential to cervical ripening, in that they may ensure mobilization of leukocytes, the major source of collagenase, and a key catabolic enzyme that dissociates and disorganizes collagen, leading to cervical ripening (Luque et al. 1998).
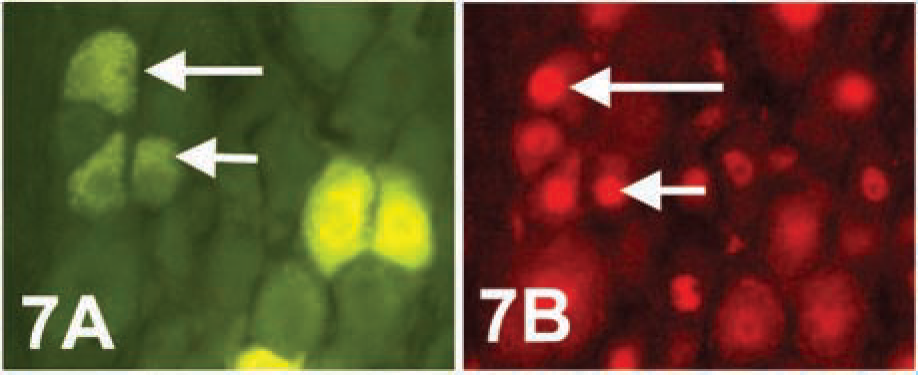
Micrographs from a rat injected with the retrograde tracer Fluorogold in the cervix at day 8 of pregnancy and then allowed to deliver a pup (parturient) before being killed for tissue harvesting. (
Angiogenic Factors in the Ripening Cervix
The angiogenic factor vascular endothelial growth factor (VEGF) has a broad spectrum of vascular effects resembling those that occur in the ripening cervix, such as vascular leakage, vasodilation, and vascular expansion (Bates and Curry 1997; Ferrara and Davis-Smyth 1997). Therefore, it is possible that VEGF could be one of the modulators of cervical microvascular remodeling in late pregnancy. To date, no study has reported direct or indirect effects of angiogenic factors (in general, and VEGF, in particular) on cervical microvascular remodeling. Nonetheless, Western blot analyses and immunoassays have shown that protein levels of VEGF and its receptors [VEGF R-1 (Flt-1) and VEGF R-2 (KDR)], and activity of VEGF's key intermediate mediators of angiogenesis [phosphorylated antiapoptotic kinase (pAkt) and nitric oxide (NO, measured indirectly as eNOS)], increase remarkably near term in the cervix (Mowa et al., unpublished data). Collectively, these findings imply that some of the microvascular changes associated with cervical ripening are also likely to be induced by VEGF, a potent vasodilator and vascular permeability– and inflammation-inducing factor (Bates and Curry 1997; Ferrara and Davis-Smyth 1997).
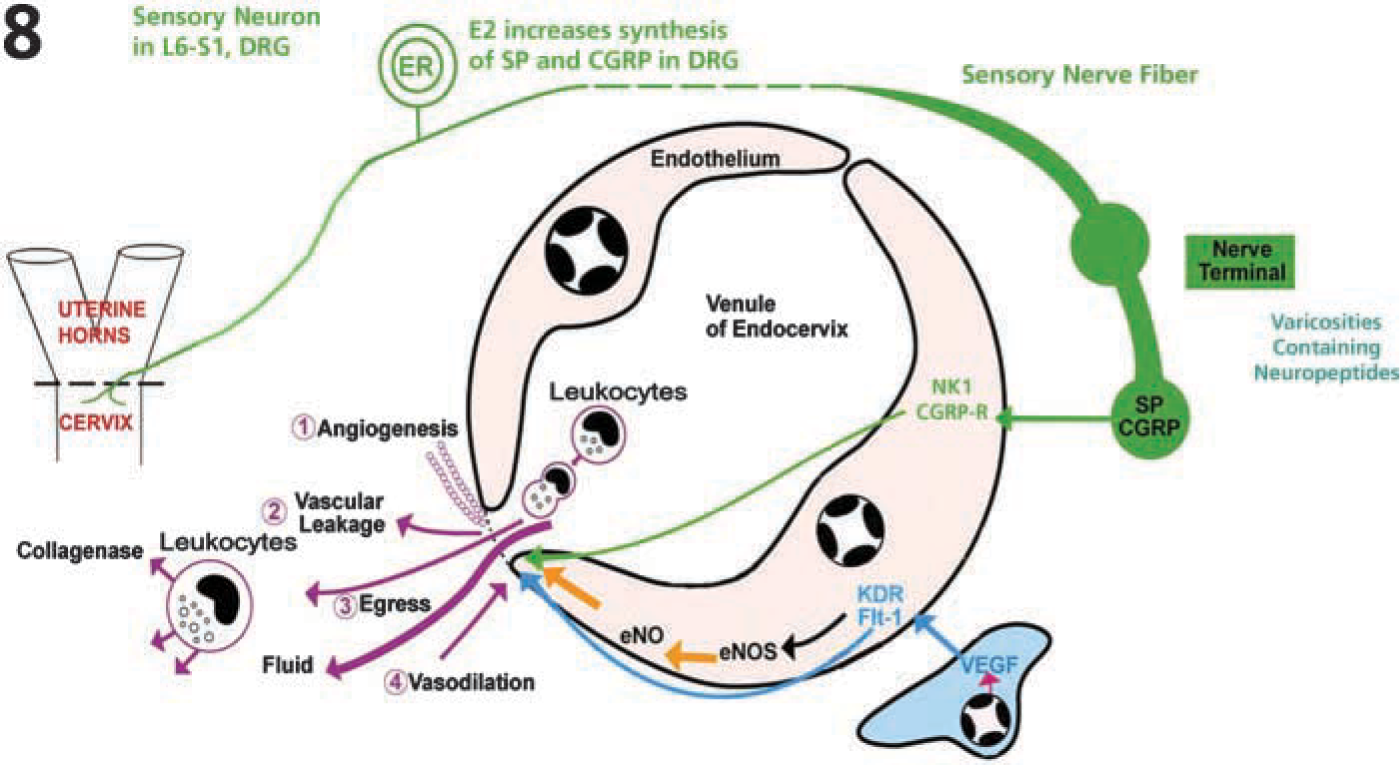
Summary diagram to illustrate potential functional relationships of sensory nerve terminals with the cervical microvasculature (illustrated above in Figures 3A–3D). A sensory neuron (green) has terminals in the uterine cervix (on the left, shown expanded on the right). Such sensory neurons express ERs, and estrogen stimulates increases in synthesis of SP and CGRP (Mowa et al., 2003a, b). SP and CGRP can be released from the terminals at the microvasculature (tan) to stimulate SP-NK1 and CGRP receptors (NK1, CGRP-R) in the endothelial cells and vascular endothelial growth factor (VEGF) in the stromal cells, respectively. Activation of NK1 and CGRP-R, and activation of VEGF receptors, KDR and Flt-1, may stimulate vascular changes directly or through the synthesis or activation of endothelial nitric oxide (eNOs-eNO system). Microvascular changes that could promote cervical ripening include (1) angiogenesis, (2) vascular leakage, (3) egress of leukocytes, and (4) vasodilation.
Link between Sensory Neuronal Factors and Angiogenic Factors in the Ripening Cervix
Exactly how endogenous neuropeptides such as SP and CGRP, released from peripheral terminals of sensory nerves, alter the cervical microvasculature remains to be elucidated. However, because levels of both neuropeptides (SP and CGRP) and angiogenic factors (VEGF) rise near term in the cervix (Mowa et al. 2003a, b, c; Mowa et al., unpublished data), it is possible that the action of these factors on the cervical microvasculature may be interconnected (Figure 8). If such a link does exist, transection of sensory nerves, described above, should alter levels of VEGF. Interestingly, bilateral pelvic neurectomy, which results in loss of sensory neuronal factors such as SP and CGRP, alters concentration of VEGF protein (Mowa et al., unpublished data) (Figure 8). These findings are very important, in that they may help unravel the potential mechanisms likely to mediate the effects of sensory neuronal factors, in general, and SP and CGRP, in particular, during parturition.
Future Directions and Conclusion
Dysfunctional cervices are among the most important etiologies of obstetrical complications, such as pre-term and protracted labor (Challis 2000). The lack of a comprehensive understanding of the precise factors triggering cervical ripening and the lack of effective treatments emphasize the need to explore and identify new targets for therapy. The recent advancements showing the interrelationship among sensory neurons of the cervix, the estrogen-ER system, and angiogenic factors in remodeling cervical microvasculature are welcome developments. Most importantly, these developments are therapeutically attractive, in that disruption or enhancement of the local vasculature has the potential to impact inflammation and influx of leukocytes, the ‘backbone’ or core of the cervical-ripening process.
Many questions remain unanswered, such as: (1) Are the effects of estrogen on L6-S1 DRG sensory neurons mediated by both membrane and genomic ER? (2) What are the specific neurochemical pheno-types of cervical sensory nerves, and what are their specific patterns of expression during pregnancy, when levels of plasma estrogen are increasing? and (3) Is there a common pathway mediating the effects of VEGF, SP, and/or CGRP and estrogen on cervical microvascular remodeling in the ripening cervix? These and other questions need to be addressed in future research.
Footnotes
Acknowledgements
The work reviewed here was supported by NIH Grant NS-22526, a NEOUCOM Research Challenge, and the NEOUCOM Foundation.