
Abstract
Glomerular expression of tensin was immunohistochemically studied in normal and diseased rat kidneys to determine whether tensin might be related to specific binding in individual glomerular cells. Normal rat kidneys displayed an intense immunofluorescence reaction for tensin along the basal aspects of proximal and distal tubule cells and parietal epithelial cells of Bowman's capsules. In glomeruli, a positive reaction for tensin was detected only in the mesangial areas. Immunoelectron microscopy revealed a positive reaction in the mesangial cell (MC) processes. RT-PCR and immunoprecipitation demonstrated mRNA and protein levels of tensin in cultured rat MCs. Mesangial tensin expression was decreased when the mesangium was injured by Habu snake venom. During the regenerative process after mesangiolysis, tensin expression was not detected in early-phase proliferating MCs that did not have extracellular matrix (ECM). The expression of tensin recovered in late-phase proliferating MCs, which became attached to regenerated ECM. It appears that tensin is related to MC attachment to surrounding ECM, which suggests that signal transduction regulated by tensin may be related to a specific mechanism of MC matrix regeneration. Furthermore, tensin can act as a marker for rat MCs because the expression of tensin was detected only in MCs in glomeruli.
T
Tensin is an actin-associated 220-kD phosphoprotein located between actin and integrin (Lo et al. 1994a; Chuang et al. 1995; Chen et al. 2000). The protein contains a functional Src homology 2 (SH2) domain (Davis et al. 1991), which is able to interact with certain tyrosine phosphorylated proteins (Salgia et al. 1995). In addition, it binds to barbed ends of actin filaments and is able to crosslink with actin filaments (Lo et al. 1994b; Chuang et al. 1995). Because of these properties, tensin has a dual role in cytoskeletal architecture and signal transduction (Lo et al. 1994c). A previous study using tensin-null mice showed that tensin was expressed in the tubule epithelial cells and was related to focal adhesion of these cells to the tubular basement membrane (Lo et al. 1997). However, expression of tensin in the glomeruli has not been examined.
In the present study, the localization of tensin was examined by immunohistochemistry (IHC) in rat kidneys, and selective localization of tensin in the glomerular MCs was identified. In addition, to explore the putative role of tensin in renal disease with mesangiolysis, the expression of tensin was analyzed in Habu snake venom (HSV) nephritis. Pathological lesions of HSV nephritis induced in Sprague-Dawley (SD) rats were characterized by mesangiolysis in the initial phase and MC proliferation in the late phase (Bradfield et al. 1977; Cattell and Bradfield 1977; Barners et al. 1994a).
Materials and Methods
Animals
All animal experiments were conducted in accordance with the National Research Council Guidelines for the Care and Use of Laboratory Animals. Adult male SD rats weighing ∼200 g were obtained from Japan Charles River Laboratories (Kanagawa, Japan).
Induction of Glomerular Disease Models
HSV nephritis was induced in adult SD rats by a single IV injection of HSV at 4.0 mg/kg body weight (Wako Pure Chemical Industries; Osaka, Japan). In addition to a control group (no injection), two experimental groups with animals sacrificed at 5 days or 10 days after the HSV injection were studied.
Fixation of Kidneys
After the rats were anesthetized with sodium pentobarbital (100 mg/kg body wt), they were perfused via the abdominal aorta with 4% paraformaldehyde (PFA) in PBS, pH 7.2, at 200 mmHg for 5 min. After perfusion of PFA, the left kidney was cut in half, dipped in 10% formaldehyde at 4C, and processed and embedded in paraffin for light microscopy. For electron microscopy, one half of the right kidney was cut into several pieces and fixed with 2% glutaraldehyde (GA) in 0.1 mol/liter phosphate buffer (PB) at 4C for 2 hr. After the tissues were washed in PB, they were postfixed with 2% aqueous osmium tetroxide at room temperature (RT) for 2 hr. Then they were dehydrated in a graded ethanol series at RT and finally embedded in Epon 812 by standard procedures. After perfusion of PFA, the right kidney was perfused with cryoprotectant 20% sucrose in PBS at 200 mmHg for 5 min. The kidney was removed and dipped in 20% sucrose solution at 4C for 2 hr. Finally, the kidney was cut into several pieces and frozen in melting isopentane cooled by liquid nitrogen to −160C for IHC.
Light and Electron Microscopy
For light microscopy, the paraffin-embedded tissues were cut into 3-μm-thick sections, mounted and stained with periodic acid-Schiff (PAS) by standard procedures. The sections were examined with an Olympus light microscope (Olympus; Tokyo, Japan).
For electron microscopy, the tissues were cut with a regular diamond knife into semi- and ultrathin sections. Semithin sections (1 μm thick) stained with 1% toluidine blue were used for light microscopy. Ultrathin sections (silver to gold) were collected on 100-mesh copper grids and double stained with 4% uranyl acetate and lead citrate. The sections were then examined with a Hitachi 7100 transmission electron microscope (Hitachi High Technologies; Tokyo, Japan) operated at 75 kV.
Immunohistochemistry
Frozen renal sections were cut into 3-μm-thick sections using the cryostat. As the primary antibody (Ab), a mouse monoclonal anti-tensin antibody (BD Transduction Laboratories; BD Biosciences, San Jose, CA) was used (Dugina et al. 2001; Kook et al. 2003). The second Ab was FITC-conjugated goat anti-mouse IgG antiserum (Cappel; ICN Pharmaceuticals., Aurora, OH). After the sections were blocked with PBS containing 2% bovine serum albumin, 2% fetal calf serum, and 0.2% fish gelatin for 15 min, they were incubated with primary Ab diluted 1:100 in blocking solution at room temperature for 60 min. The sections were then incubated with secondary Ab diluted 1:100 in blocking solution with 10% normal rat serum at RT for 30 min. After immunostaining, the sections were examined with a Leica fluorescence microscope (Leica Microsystems; Tokyo, Japan).
For immunoelectron microscopy, the frozen renal sections were cut into 3-μm-thick sections with a cryostat. After the sections were blocked with blocking solution for 15 min, they were incubated with primary Ab diluted 1:100 in blocking solution at 4C overnight. The sections were incubated with biotinylated goat anti-mouse IgG at 4C for 5 hr and were then incubated with avidin-peroxidase complexes from the Vectastain Elite ABC Kit (Vector Laboratories; Burlingame, CA) at RT for 30 min, followed by incubation with the peroxidase substrate solution diaminobenzidine in PBS for a few minutes. The sections were fixed with 2% GA at 4C for 60 min. They were then dehydrated in a graded series of ethanol at RT and finally embedded in Epon 812 by standard procedures.
For immunostaining of rat MCs (described below), two types of cells maintained in normal medium or FCS-free medium for 5 days were used. Primary rat MCs were placed on collagen type I-coated glass coverslips (Iwaki Glass; Tokyo, Japan) and cultured continuously with normal medium or FCS-free medium for 12 hr at 37C. After the cells were washed with PBS, they were fixed with 3% PFA containing 0.5% Triton X-100 (Sigma; St Louis, MO) for 5 min and then fixed with 3% PFA for 20 min. After the cells were incubated with blocking solution for 30 min, they were incubated with anti-tensin Ab diluted 1:100 in blocking solution at RT for 60 min. The cells were then incubated with Alexa-Fluor 488 goat anti-mouse IgG Ab (Molecular Probes; Eugene, OR) diluted 1:200 in blocking solution at RT for 60 min. After immunostaining, the cells were examined with a confocal microscope (Japan Bio-Rad Laboratories; Tokyo, Japan).
Preparation of Rat Glomeruli and Culture of Primary Rat MCs
After the rats were anesthetized, the kidneys were removed for RNA and protein extraction. Glomeruli were isolated by differential sieving methods (Striker et al. 1987). The glomeruli were incubated on type I collagen-coated 10-cm dishes to obtain MCs. Primary rat MCs were maintained in Dulbecco's modified Eagle's medium (Sigma) supplemented with 5% FCS, 100 U/ml penicillin G, and 100 mg/ml streptomycin at 37C.
RNA Preparation and RT-PCR in Rat MCs and Kidney Cortex
Rat MCs (1.2 × 106) were incubated on type I collagen-coated 10-cm dishes. Total RNA of rat MCs and kidney cortex were extracted with an RNeasy Mini Kit (QIAGEN; Hilden, Germany) for RNA preparation. One microgram of total RNA was reverse-transcribed using oligo (dT) primers (Life Technologies; Rockville, MD) and reverse transcriptase (Superscript II; Life Technologies). The single-strand cDNA product was denatured and amplified in the Gene Amp PCR System 9600 (Perkin-Elmer; Norwalk, CT) with two primers under the following conditions: 1 min at 95C; then 30 sec at 94C, 30 sec 54C and 1 min 72C for 35 cycles; and finally 72C 10 min and 4C. The cDNAs were amplified using various primers specific only for cDNA sequences of rat tensin (forward 5′-ACATTACCCCCTCAACACTG-3′ and reverse 5′-GTTCTGGATTCAAGGTGGTC-3′). The amplified products were confirmed by sequencing. The amplified products were electrophoresed with a 2% agarose gel, stained with ethidium bromide, and photographed under a UV lamp.
Immunoprecipitation of Rat MCs and Kidney Cortex
Rat MCs and kidney cortex were lysed in 1% NP-40 buffer [1% NP-40 = 300 mM NaCl, 100 mM Tris-HCl (pH 7.5), 0.1% sodium dodecyl sulfate (SDS), 2 mM ethylenediamine-tetraacetate] containing proteinase inhibitors (1 mM phenylmethylsulfonyl fluoride, 1 mM pepstatin A, 1 mM E-64C) on ice. Insoluble tissue lysates were removed by centrifugation at 15,000 × g for 5 min. The lysates were subjected to immunoprecipitation with a mouse monoclonal anti-tensin antibody and protein G-PLUS-Agarose (Santa Cruz Biotechnology; Santa Cruz, CA) at 4C overnight. The immunoprecipitates were collected on protein G-PLUS-Agarose, washed with NP-40 buffer, and then extracted from the beads by boiling in SDS sample buffer. They were analyzed by immunoblot analyses. Immunoblotting analyses were carried out according to standard protocols (Laemmli 1970; Towbin et al. 1979) and visualized by chemiluminescent methods with ECL plus immunoblot detection kits (Amersham Pharmacia Biotech; Poole, UK).
Results
Immunofluorescence
In the cortex of normal rat kidney, a positive reaction for tensin was detected in tubuli, Bowman's capsules, and glomeruli (Figure 1A). The expression of tensin intubuli displayed a continuous linear fluorescence at the basal aspects, but the intensity of the immunoreactivity was variable among tubule segments. Proximal convoluted tubuli were more intensely stained than other segments. In Bowman's capsules, the positive reaction was also detected at the basal aspects of parietal epithelial cells. The distribution of tensin was similar to that of actin fiber bundles in the cells. When the cells were cut vertically, linear immunoreactivity was detected in the cells. When the cells were cut obliquely, the immunoreactivity appeared as several short parallel lines. In glomeruli, a positive reaction for tensin was detected in the mesangium (Figure 1B). The intensity of the positive reaction was almost identical in each mesangial area of individual glomeruli. Other components in the cortex, i.e., blood vessels, interstitial cells and glomerular capillary walls, did not react with tensin.
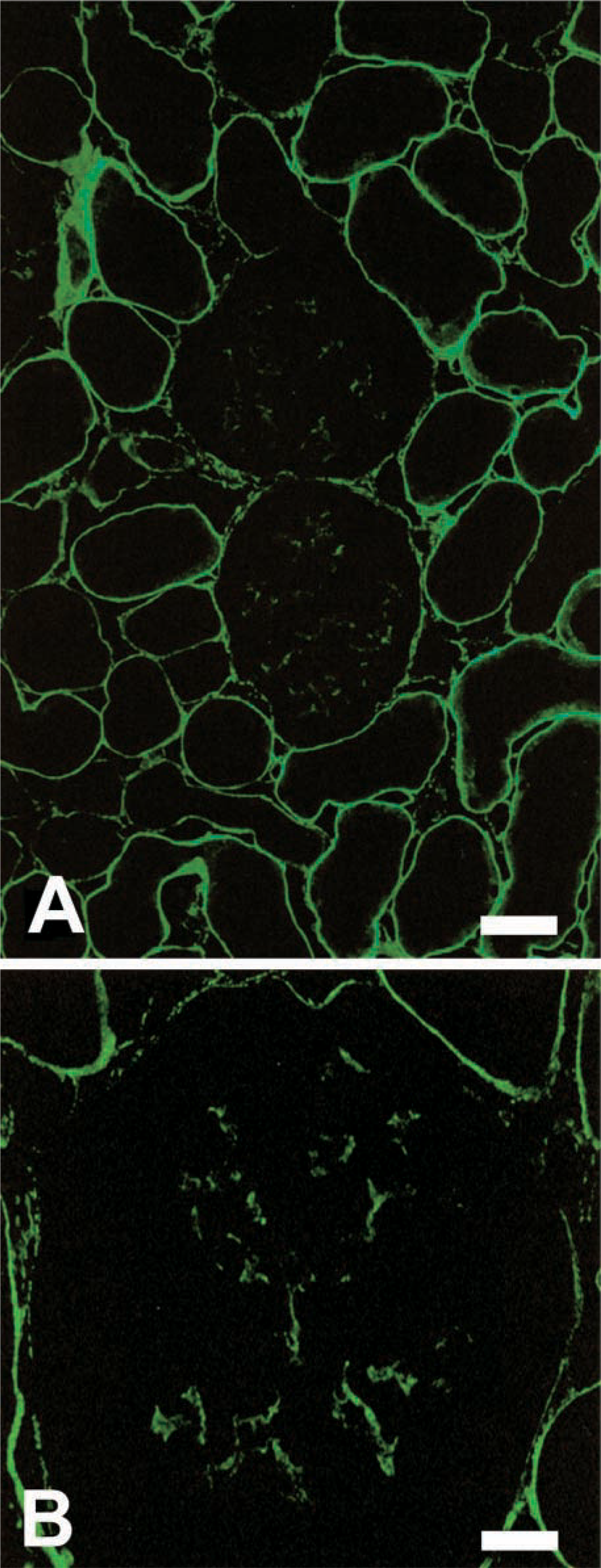
Immunofluorescence of normal rat kidney for tensin. (
Immunoelectron Microscopy
The presence of tensin in tubule epithelial cells and parietal epithelial cells of Bowman's capsules was clearly observed by immunoelectron microscopy. In each cell, immunoreactive products were localized at the inner aspects of the basal cell membranes attached to individual basement membranes (Figures 2A–2C). In the mesangium, immunoreactivity for tensin was localized in MCs (Figure 2D). Most of the immunoreactivity was localized in processes of MCs. Among individual processes, tongue-like processes extending toward the mesangial angles of the GBM exhibited the strongest reactivity (Figure 2E).
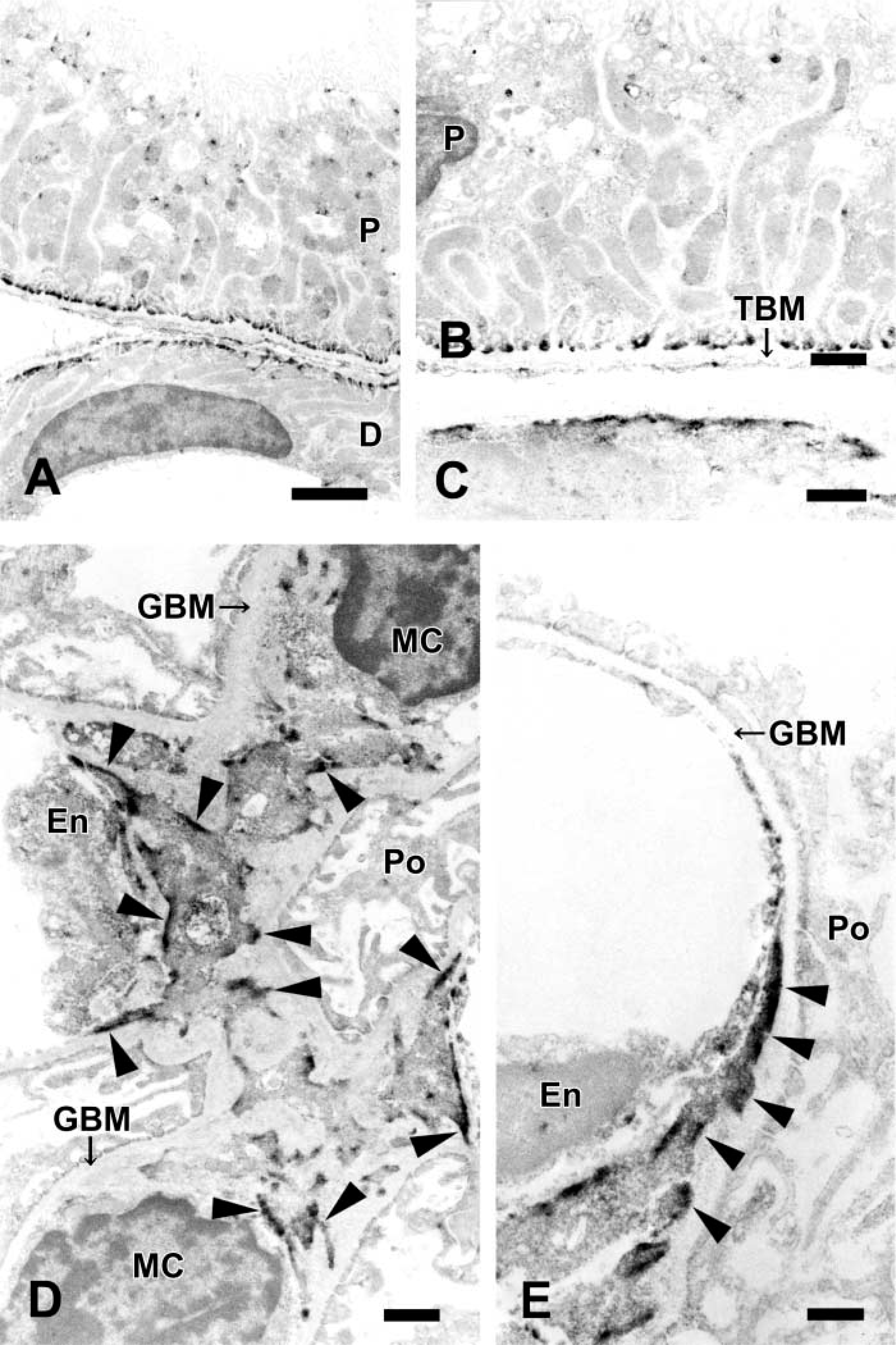
Immunoelectron microscopy of normal rat kidney for tensin. (
RT-PCR and Immunoprecipitation
Using RT-PCR, a single band of 324 bp for tensin was detected in cultured rat MCs and kidney cortex (Figure 3A). Immunoprecipitation demonstrated a single immunoreactive band at a position of 220 kD in extracts of cultured rat MCs and kidney cortex (Figure 3B).
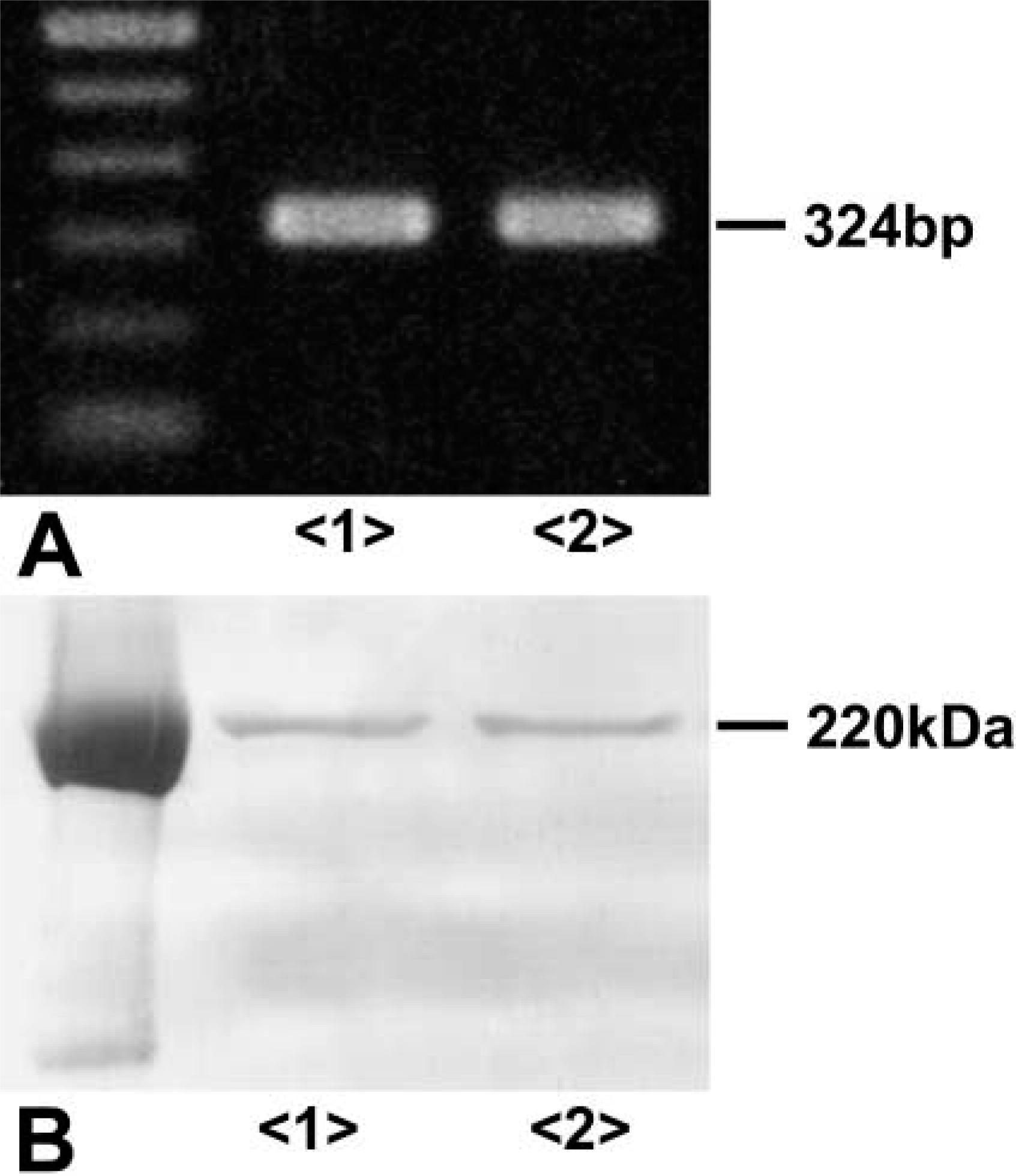
(
Cultured Mesangial Cells
In MCs cultured in normal medium, tensin was expressed in the ends of the actin filament where the cells were attached to the substrate (Figure 4A). Tensin was expressed as elongated or dot-like structures. MCs cultured in FCS-free medium also expressed tensin in the ends of the actin filament (Figure 4B). However, it appeared that the intensity was slightly increased and the elongated structures changed to broader structures.
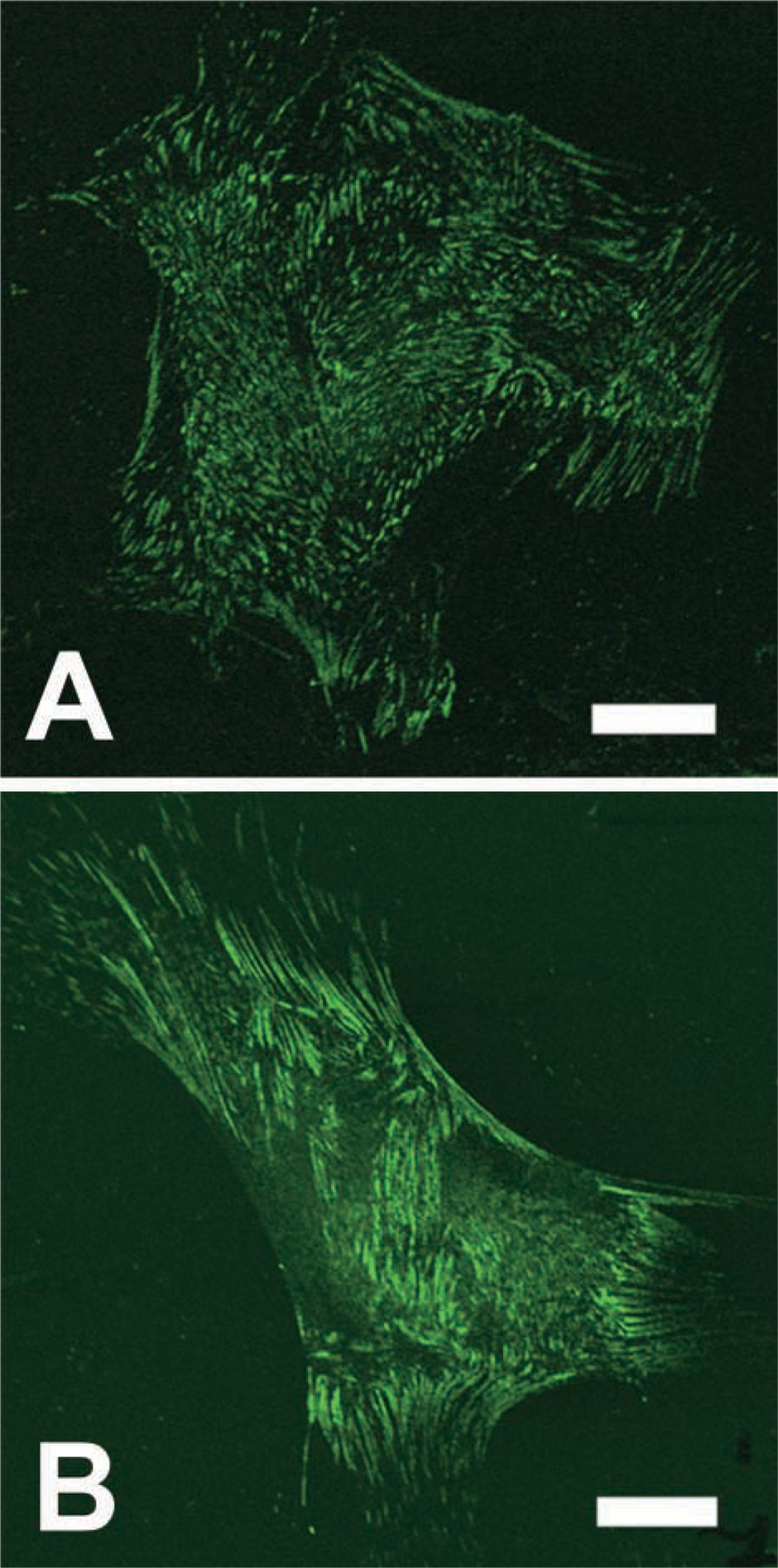
(
Habu Snake Venom Nephritis-induced SD Rat Kidney
The expression of tensin was decreased in the mesangial areas of mesangiolytic glomeruli, although a positive reaction for tensin similar to that in normal rat glomeruli was detected in most glomeruli. At day 5, the early phase of the regenerative process of the mesangium, the positive reaction for tensin was still decreased in mesangial areas (Figure 5A). However, at day 10, the late phase of the regenerative process, a strong reaction for tensin was observed in mesangial areas (Figure 5B). In both glomeruli, α-smooth muscle actin (α-SMA) was detected (data not shown) as previously reported (Barners et al. 1994a,b). In PAS-counterstained sections of the same IF sections, MC proliferation was observed in mesangial areas of each glomeruli (Figures 5C and 5D).
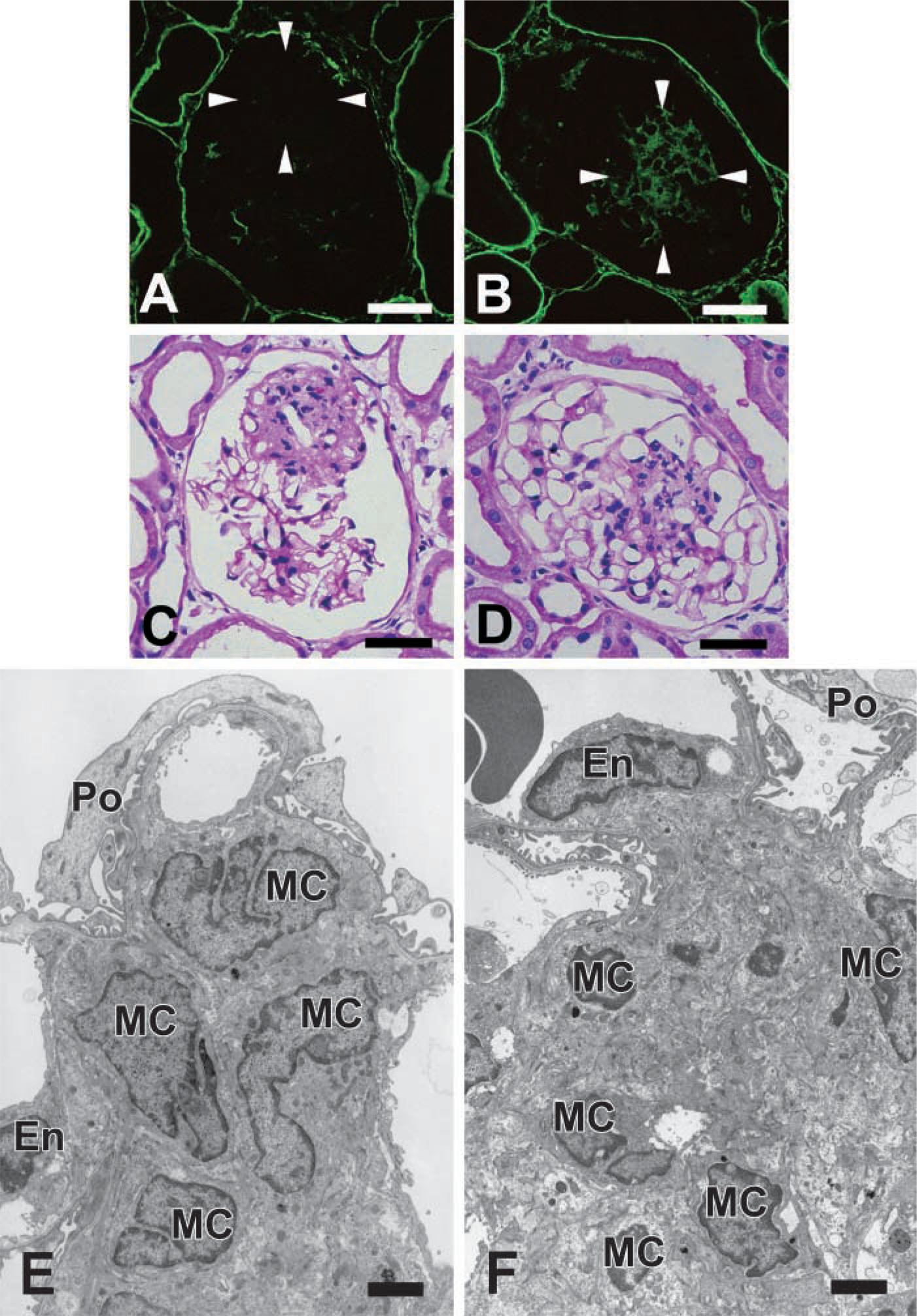
Changes in expression of tensin in the regenerative process after mesangiolysis by HSV nephritis on day 5 (
Proliferative MCs showed a significant difference in both phases by electron microscopy. At day 5, most proliferative MCs did not have surrounding extracellular matrix (ECM) and were in direct contact with neighboring MCs (Figure 5E). Some of the MCs had ECM but most of the ECM was basically immature. At day 10, proliferative MCs were also observed in mesangial areas. However, they had regenerative surrounding ECM and were in contact with other MCs through the mature ECM (Figure 5F).
Discussion
Tensin-1 is a 220-kD protein localized in the trans-membrane junctions between the ECM and the cytoskeleton (Lo et al. 1994a; Chuang et al. 1995; Chen et al. 2000). A recent study suggests that tensin-2 (170 kD), which has some homology to tensin-1, is also localized in focal adhesions in human tissues (Chen et al. 2002). Tensin is particularly interesting in the myriad of focal adhesion proteins because it can bind to actin microfilaments at multiple sites (Lo et al. 1994b), enabling tensin both to cap the growing ends of actin filaments and to crosslink actin filaments (Lo et al. 1994b; Chuang et al. 1995). In addition, tensin is phosphorylated on tyrosine residues (Salgia et al. 1995) and contains an SH2 domain (Davis et al. 1991), suggesting that tensin might participate in signal transduction cascades relevant to various cellular functions, including cell migration (Chen et al. 2002; Chen and Lo 2003), differentiation (Hiura et al. 1995), regeneration (Ishii and Lo 2001), and apoptosis (Kook et al. 2003). Several factors, including the ECM, platelet-derived growth factor, thrombin, angiotensin, and oncogenes, induce tensin-mediated signal transduction (Chen et al. 2002). The presence of these characteristics in a single molecule suggests that tensin may contribute to MC function.
The present study demonstrates that tensin, in addition to its localization in the tubuli and Bowman's capsules, is specifically expressed in the rat glomerular mesangial areas. Subcellular distribution of tensin examined by immunoelectron microscopy revealed that its protein is mostly localized in MC processes that have well-developed microfilament bundles and are in contact with the perimesangial GBM. Immunoprecipitation and RT-PCR using cultured rat MCs confirmed the presence of a protein of 220-kD molecular mass identical to that of tensin and the expression of mRNA for the protein. Although we did not check the specificity of the immunoreactivity by a preincubation test with a corresponding antigen, these findings strongly suggest that tensin itself is contained in MCs of rat glomeruli.
In this study, the expression of tensin was clearly localized in the MC processes where microfilament bundles terminate in the cell membrane to connect with the surrounding GBM. Among these processes, the most prominent expression of tensin was detected in juxtacapillary tongue-like processes that interconnect with the GBM at the two opposing mesangial angles of a single capillary. The distending forces of the glomerular capillary wall are balanced at the mesangial angles and at the mesangium-GBM interface by a complex anchoring system that connects the GBM to the mesangial processes (Sakai and Kriz 1987; Kriz et al. 1990a,b; Lemley et al. 1992). These findings point to the essential involvement of tensin in the mechanical role of MCs, i.e., stabilization of glomerular capillary architecture. On the basis of findings that tensin can bind to actin microfilaments at multiple sites, as described above, it is reasonable to assume that tensin expressed in the MC processes is an important molecule for the contractile and adhesion apparatus of MCs. In addition, a recent study reported that MCs organize the glomerular capillaries by adhering to the G- domain of laminin α5 in the GBM via integrin α3β1 and Lu, a member of the immunoglobulin superfamily (Kikkawa et al., 2003). Therefore, we suggest that MCs maintain the glomerular capillary architecture in various ways.
The transient strong expression of tensin in MCs during the recovery phase after mesangiolysis induced by HSV suggests that some specific mechanism of MC regeneration might be regulated by tensin-mediated signal transduction. Background data permitting speculation about specific function of the protein are lacking, but it is noteworthy that expression is detected not in early but in late regenerative processes after HSV injury. The same late appearance is also observed in the deposition of ECM such as fibronectin (Barners et al. 1994b,1995). As demonstrated in cell culture studies (Gauer et al. 1997; Prols et al. 1999), MCs and matrix proteins might interact to maintain cellular function, resulting in the recovery of normal structure and function of the mesangium.
Recent studies showed that ECM adhesion formed by cultured fibroblasts can be classified into two major types by molecular and structural criteria, i.e., classical focal contact (FC) and fibrillar adhesion (FA) (Zamir et al. 1999; Katz et al. 2000). These were characterized by distinct complements of integrins and cytoskeletal anchor molecules. FCs are located mainly at the cell periphery, bound mainly to vitronectin fibrils, and contain relatively high levels of vinculin, paxillin, α-actinin, talin, focal adhesion kinase, and αvβ3 integrin. On the other hand, FAs are located more centrally in cells, are characteristically elongated or dotlike structures, are rich in tensin and α5β1 integrin, and are bound to fibronectin fibrils (Zamir et al. 1999; Katz et al. 2000). These two types of adhesion appear to differ not only in their shape and molecular composition but also in their functions. FCs cannot translocate smoothly as a result of contractile forces and develop high tension. In contrast, FAs are moved dynamically by actomyosin-driven pulling (Zamir et al. 2000; Zamir and Geiger 2001). In cultured mesangial cells, differences in expression of tensin were observed under different culture conditions. In the quiescent condition, expression of tensin might be stronger with changes in the structures. Although we did not examine the presence of FAs in the MCs, these findings suggest that MCs may contain FAs and control cell functions, including cell adhesion and signal transduction, in different ways from other glomerular cells under all conditions. In the future, it will be necessary to examine the presence of FAs and the expression of tensin under abnormal conditions in the MCs.
At present, two proteins are used as markers of MCs, i.e., α-SMA and Thy-1.1. α-SMA is detected in MCs during proliferative and regenerative processes after severe injury but is not detected in normal MCs (Johnson et al. 1991; Alpers et al. 1992; Utsunomiya et al. 1999; Kaneko et al. 2001). Thy-1.1 is expressed in rat MCs but not in other mammalian MCs (Ishizaki et al. 1980; Harada et al. 1982; Yamamoto and Wilson 1987; Holthofer et al. 1995). The specific expression of the protein in rat MCs in this study suggested that tensin can be used as a marker for rat MCs. We also identified the localization of tensin in human MCs by IHC using renal biopsy specimens from patients with chronic glomerulonephritis with or without MC proliferation (Yamashita et al. 2004). This finding suggests that tensin can also act as a marker protein for human MCs. In comparison with these two proteins, tensin has the advantage of detecting rat and human MCs under both normal and proliferative conditions.
In summary, it appears that tensin is related to MC attachment to the surrounding ECM, by which MCs, together with the GBM, form the basic structural stabilizing system of the glomerular capillary architecture. It was also suggested that the signal transduction regulated by tensin may be related to specific mechanisms of MC and matrix regeneration. Furthermore, tensin can act as a marker for rat and human MCs because the expression of tensin was detected only in MCs.
Footnotes
Acknowledgements
We thank M. Yoshida, K. Sato, J. Nakamoto, T. Shibata, and T. Shigihara (Juntendo University; Tokyo, Japan) for their technical assistance.