
Abstract
When rat glomerular mesangial cells (MCs) are cultured with IGF-1 they accumulate intracellular lipid and take on foam cell morphology. These changes were characterized by electron microscopy and Nile red staining. To define the mechanism responsible for IGF-1-mediated lipid uptake, MCs were evaluated for endocytosis, scavenger receptor activity, and receptor-mediated uptake by the LDL receptor. Lipid accumulation was markedly increased when MCs were cultured with IGF. The primary route of uptake was through enhanced en-docytosis. Lipid-laden MCs have decreased phagocytic capacity and disrupted cytoskeletons. These data show that IGF-1 induces MC to take on a foam cell morphology and that lipid-laden MCs have impaired phagocytic function.
Lipid Accumulation in the vessel wall is considered to be an early step in atherosclerosis. Lipoproteins bind to matrix proteins in vessel walls, where they become oxidized (Chisolm 1991; Scoble 1999). Oxidized lipoproteins activate endothelial cells to release chemoattractants that recruit macrophages into the vessel wall. Macrophages take up modified lipoproteins via scavenger receptors and, as this material accumulates, they become foam cells (Aviram et al. 1988). As the lesion progresses, vascular smooth muscle cells proliferate and also accumulate intracellular lipid. The pathogenesis and progression of the lesion is fueled by various factors, including hyperlipidemia, abnormalities in low-density lipoprotein (LDL) and scavenger receptors, regulatory cycles of lipid metabolism and clearance, and inflammatory cytokines (Bayes-Genis et al. 2000). Similar mechanisms may contribute to progressive sclerosis in the glomerulus.
In the glomerulus, the mesangial cell (MC) functions as a pericyte with properties of a vascular smooth muscle cell. Like the smooth muscle cell, it similarly expresses LDL and scavenger receptors (Ruan et al. 1999), secretes increased amounts of extracellular matrix (Wheeler et al. 1994) and monocyte chemoattractant protein 1 (Lee and Kim 1998) in response to oxidized LDL, and has mitogenic responses to LDL (Grone et al. 1992). In some diseases, e.g., focal and segmental glomerulosclerosis and hyalinosis, MCs exhibit a foam cell morphology, which suggests that mechanisms that are operative in atherosclerosis may also contribute to some forms of glomerulosclerosis (Hirano 1999; Miyata et al. 1999; Scoble 1999; Bayes-Genis et al. 2000).
Considerable work has focused on the details of LDL and scavenger receptor biology (Chait and Heinecke 1994), as well as the role of growth factors in the progression of atherosclerosis (reviewed in Bayes-Genis et al. 2000). In our recent studies of the effects of insulin-like growth factor 1 (IGF-1) on MC cytoskeletal organization (Berfield et al. 1997) and migration (Abrass et al. 1997), we observed that rat MCs cultured in IGF-1 (100 nM) for longer than a week take on the appearance of foam cells. The present studies were undertaken to confirm that the material within the IGF-1-treated MCs is lipid, to evaluate the role of IGF-1 in intracellular accumulation of lipid, and to examine the functional consequences of lipid accumulation to the MCs. These studies show that IGF-1 stimulates MCs to take up lipid by generalized endocytosis and that lipid-laden MCs have impaired phagocytic function.
Materials and Methods
Materials
The following reagents were purchased: phagocytosis assay kit, FITC-dextran (10,000 MW), Nile red, Bodipy FL cholesteryl ester, Bodipy phalloidin, Texas Red phalloidin and Bodipy acetylated low-density lipoprotein (AcLDL) (Molecular Probes; Eugene, OR); trypan blue (Sigma; St Louis, MO); chambered tissue culture glass slides (NUNC; Naperville, IL); Fluoromount (Fisher Scientific; Pittsburgh, PA); poly/bed 812 resin (Ted Pella; Redding, CA); human recombinant IGF-1 (Collaborative Research; Waltham, MA).
Experimental Design
MCs were cultured in routine medium (20% FCS-RPMI) with and without IGF-1 (100 nM) for 7 days and in growth arrest medium (2% FCS-RPMI) for 24 hr. To confirm that IGF-1-treated MCs accumulate lipid, MCs were examined by routine electron microscopy and stained with Nile red. To examine the mechanisms whereby IGF-1 stimulates lipid uptake, particle uptake by fluid-phase pinocytosis, scavenger receptors, and LDL receptors were measured as described below. To examine the impact of intracellular lipid accumulation on function, MCs were tested for their ability to phagocytose particles and to reorganize their cytoskeleton.
Cell Culture
Rat glomerular MCs were prepared by modification (Abrass et al. 1994,1995) of routine methods (Kreisberg and Karnovsky 1983). Minced rat kidney cortex was sieved. Isolated glomeruli were plated in medium containing a 1:1 mix of 20% FCSRPMI 1640 and previously collected glomerular conditioned medium. MC outgrowths were harvested and passed in this medium for an additional week, after which the conditioned medium was omitted. The supplemental insulin routinely added to MC cultures was omitted. MCs were cloned and studied at passages 8–12. Medium was replaced every 72 hr.
Lipid Accumulation and Lipid Uptake in MCs
MCs (2 × 104/ml) were plated in glass chamber slides, grown for 7 days in 20% FCS-RPMI medium with or without IGF-1 (100 nM), then placed in 2% FCS-RPMI for 24 hr. For measurement of accumulated neutral lipids, 2.5 μg Nile red in glycerol was added to MC cultures for 5 min and processed for fluorescence microscopy. Other slides were incubated for 4 hr with 20 μg/ml FITC-dextran to measure fluid-phase endocytosis, with fluoresceinated acetylated LDL for scavenger receptor function and fluoresceinated Bodipy cholesteryl esters for the LDL receptor-mediated and other nonspecific endocytosis. Trypan blue was used to quench particles that were not endocytosed. Each condition was measured in triplicate.
Phagocytosis
MC phagocytosis was measured using the phagocytosis assay kit in untreated cells and those grown in IGF-1 (100 nM). Phagocytosis was determined after 1 hr by uptake of fluoresceinated E. coli. Complete internalization was confirmed by quenching with trypan blue and slides were processed for fluorescence microscopy. Other slides were counterstained with Texas red phalloidin and processed for scanning confocal microscopy.
Cytoskeletal Organization
F-actin organization was determined by Bodipy phalloidin staining on permeablized MCs and examined by fluorescence microscopy.
Fluorescence Microscopy
Chamber slides of various experiments were rinsed in PBS and fixed in 2% paraformaldehye for 20 min. For Bodipy or Texas red phalloidin staining, MCs were permeabilized with 0.05% Triton X-100 for 3 min, rinsed in PBS, and incubated with primary antibody for 20 min at RT. MCs were mounted in Fluoromount and observed with a Leitz microscope equipped for epi-illumination. The fluorescent images were recorded on Kodak DX 400 black-and-white film or on Ektachrome film.
Confocal Microscopy
Phagocytosis slides counterstained with Texas red phalloidin were scanned on a Leica TCS-SP confocal microscope using absorptions of 596 nM and 488 nM. Images were collected in sequential xz-series and yz-series, as well as overlay images. Acquired images were exported to Adobe Photoshop for further processing.
Electron Microscopy
MCs were rinsed with PBS, fixed in 2% gluteraldehyde for 2 hr at 4C, treated with 1% osmium tetroxide for 30 min at RT, and washed three times with cacodylate buffer. They were sequentially dehydrated through ascending concentrations of alcohol (35–100%) to resin and polymerized. A Sorvall MT6000 ultramicrotome was used for sectioning and specimens were examined on a JEOL TEM S100.
Results
IGF-1-induced Foam Cell Formation
Compared to MCs cultured for 7 days in growth medium alone, cells supplemented with IGF-1 (100 nM) displayed morphological changes consistent with a foam cell. These include a convoluted nucleus and an expansive cytoplasm with extended Golgi and endoplasmic reticulum. The cytoplasm contained multiple lipid- and myelin-filled vacuoles and distorted fat-laden mitochondria (Figure 1).
To confirm that MCs with foam cell morphology contain increased amounts of lipid, untreated MCs or those treated with IGF-1 were stained with Nile red. As shown in Figure 2, IGF-1-treated MCs had increased neutral lipid content compared to untreated cells. FCS contains 200 mg/dl of total lipid, including cholesterol, HDL, LDL, triglycerides, and phospholipids. Therefore, medium containing 20% FCS has 40 mg/dl of lipid. Intracellular lipid accumulation may result from uptake from the medium and/or intracellular synthesis.
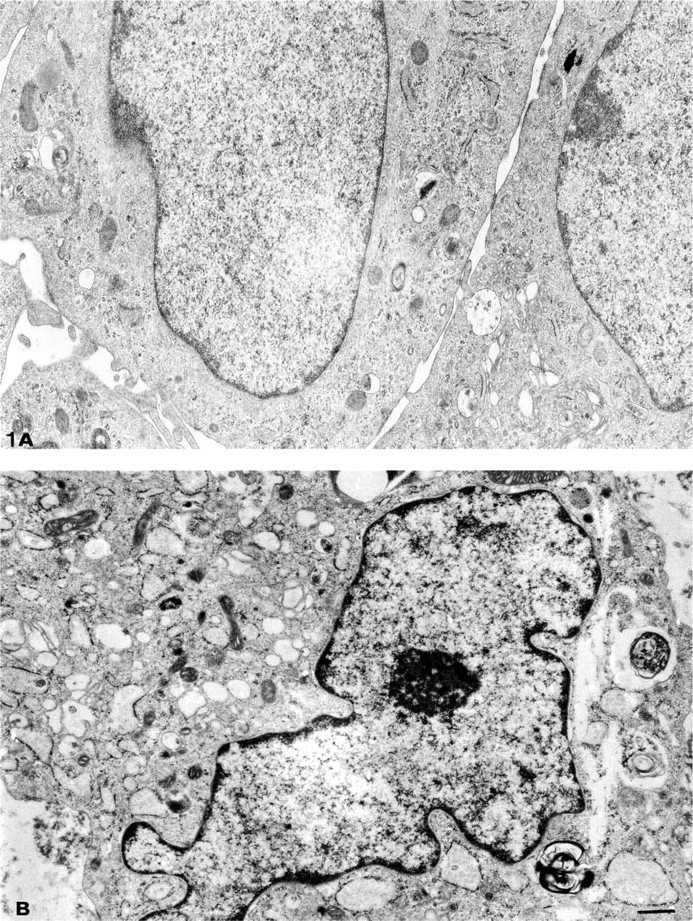
Electron micrograph of MCs grown in 20% FCS-RPMI 1640 (growth medium) (
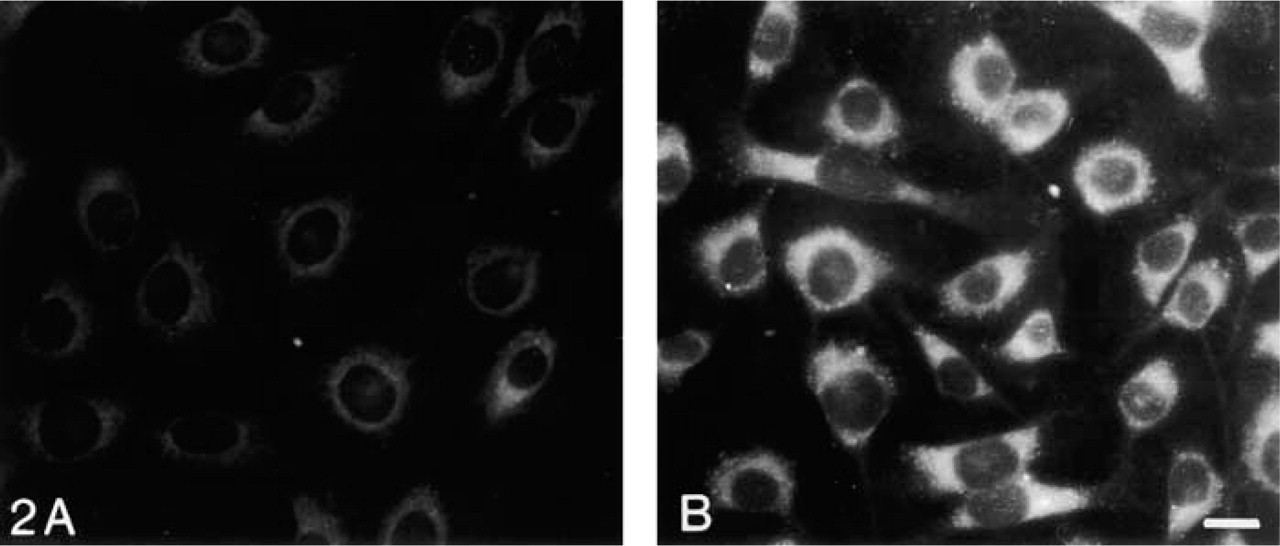
Lipid accumulation as detected by Nile red staining. MCs grown in 20% FCS-RPMI 1640 (growth medium) for 1 week (
IGF-1 Stimulation of Lipid Uptake by MCs
Having shown that IGF-1 treatment of MCs leads to lipid accumulation in MCs, we analyzed IGF-1-induced uptake of various test substances to determine the pathways involved. As shown in Figure 3, FITC-dextran was scarcely seen within untreated MCs even after 7 days in growth medium (Figure 3A, whereas uptake was increased in IGF-1-treated cells in regular growth media (Figure 3B. This is consistent with the known effect of IGF-1 in increasing membrane inter-nalization, turnover, and rates of general endocytosis (Berfield et al. 1997). FITC-dextran is taken into the cells via non-coated flask-shaped membrane invaginations or caveolae (Goldstein et al. 1985; Van Deurs et al. 1993).
Within an hour, small amounts of labeled AcLDL were taken up by both untreated and IGF-1-treated MCs (data not shown). AcLDL initially taken up by untreated MCs is no longer present by 7 days (Figure 3C. However, IGF-1-treated MCs show slightly increased intracellular accumulation of AcLDL (Figure 3D. These results suggest that IGF-1 has no direct effect on scavenger receptor-mediated uptake of AcLDL. However, IGF-1 may affect scavenger receptors after chronic exposure or may influence the turnover of intracellular lipid such that oxidized LDL accumulates within the cell.
To investigate lipid uptake by receptor-mediated endocytosis in clathrin-coated pits, where the LDL receptor is located, as well as “selected transport of lipo-protein” (Reaven et al. 1995), MCs were incubated with labeled cholesteryl esters. Within 1 hr of treatment, very little receptor-mediated uptake of cholesteryl esters was detected in untreated or IGF-1-treated MCs (not shown). After 7 days in growth medium, a few control MCs showed diffuse cholesteryl esters with distinct cytoplasmic and perinuclear staining (Figure 3E. Intracellular accumulation of lipid was increased in IGF-1-treated MCs (Figure 3F. It is possible that long-term exposure to IGF-1 modulates the LDL receptor or alters lipid efflux and turnover. Alternatively, various forms of lipids may be internalized by “selective transfer of cholesteryl ester,” and by other endocytosis of soluble as well as receptor-bound particles (Reaven et al. 1995), which becomes cumulative over time.
Impact of Lipid Accumulation on Phagocytosis
Phagocytosis is the process by which cells recognize and engulf large particles. MCs are known to take up large particles, including immune complexes (Hora et al. 1992). Phagocytosis is receptor- and actin-dependent. Therefore, it is possible that lipid accumulation within MCs might alter phagocytosis. Untreated MCs actively took up labeled E. coli particles (Figure 4A, confirming phagocytosis of E. coli. Complete engulf-ment of particles was confirmed because the particles continue to fluoresce after quenching with trypan blue. In contrast, most MCs cultured in IGF-1 for 1 week showed decreased ability to phagocytose E. coli particles (Figure 4B. To determine if E. coli particles were actually ingested, laser scanning confocal microscopy was performed. MCs were counterstained with Texas red phalloidin to examine the relationship of actin fibers and the phagocytosed FITC-labeled E. coli. Although when unquenched both untreated (Figure 5A and IGF-1-treated MCs (Figure 5C appeared to have phagocytosed E. coli (green), YZ sections reveal that untreated MCs have ingested the bacterial particles (Figure 5B, whereas IGF-1-treated MCs have not (Figure 5D.
Cytoskeletal Organization
MC phagocytosis, migration, and contraction all require an intact cytoskeleton that can rapidly reorganize in response to a variety of stimuli (Camussi et al. 1993). In the present study, untreated MCs are well-spread and have distinct criss-crossed stress fibers (Figure 6A. We previously showed that IGF-1 induces rapid reorganization of the cytoskeleton (Berfield et al. 1997) and that MCs become kite-shaped or bipolar, with stress fibers parallel to the polarity. After 1 week of treatment with IGF-1, stress fibers are lost and condensates of actin are seen throughout the cell body (Figure 6B. These findings indicate that, in association with lipid accumulation, MCs chronically exposed to IGF-1 show disruption of the actin cytoskeleton.
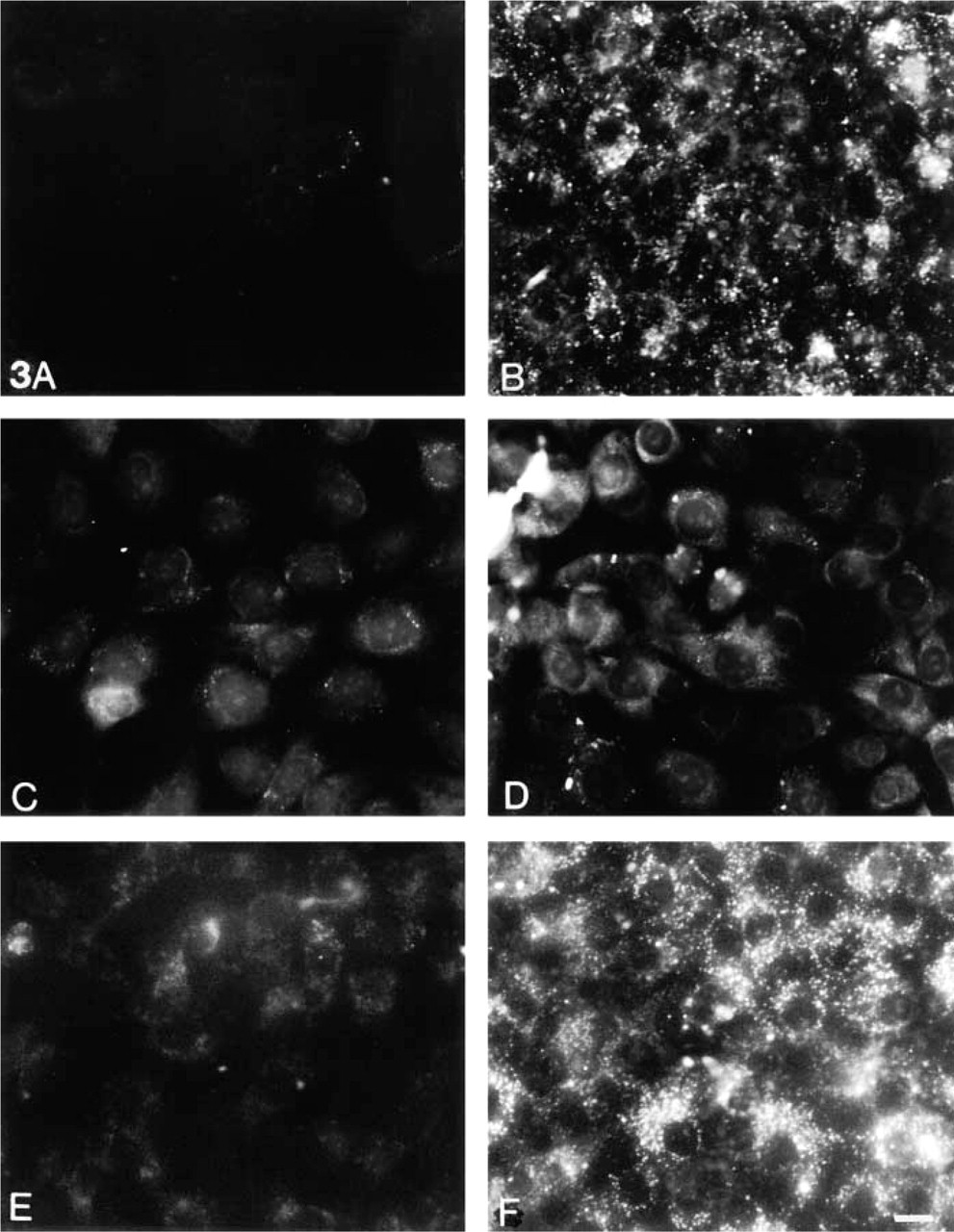
Particle uptake by MCs. Fluid-phase and non-clathrin-mediated endocytosis as measured by uptake of FITC-dextran (
Discussion
In this study we show that long-term exposure of MCs to IGF-1 induces foam cell formation, as indicated by ultrastructural characteristics and accumulation of neutral lipids. This occurs in routine medium with relatively low concentrations of lipid. Studies of particle uptake indicate that a primary effect of IGF-1 is to enhance endocytosis. Although this mechanism can lead to generalized particle uptake from the medium, lipids accumulated to a marked degree. This suggests that lipid disposal mechanisms may be impaired or may become overloaded. Once MCs become lipid-laden and assume the appearance of foam cells, they are no longer able to phagocytose particles. Mesangial foam cells have a disrupted cytoskeleton, which may in part be responsible for impaired cell function because an intact cytoskeleton that can be rapidly reorganized is required for phagocytosis.
IGF-1 and the Mechanisms of Intracellular Lipid Accumulation
IGF-1 stimulated MCs uptake and accumulation of lipid by several mechanisms. Using FITC-labeled dextran,
Phagocytosis. Phagocytosis was assessed by uptake of FITC-labeled E. coli-untreated MCs (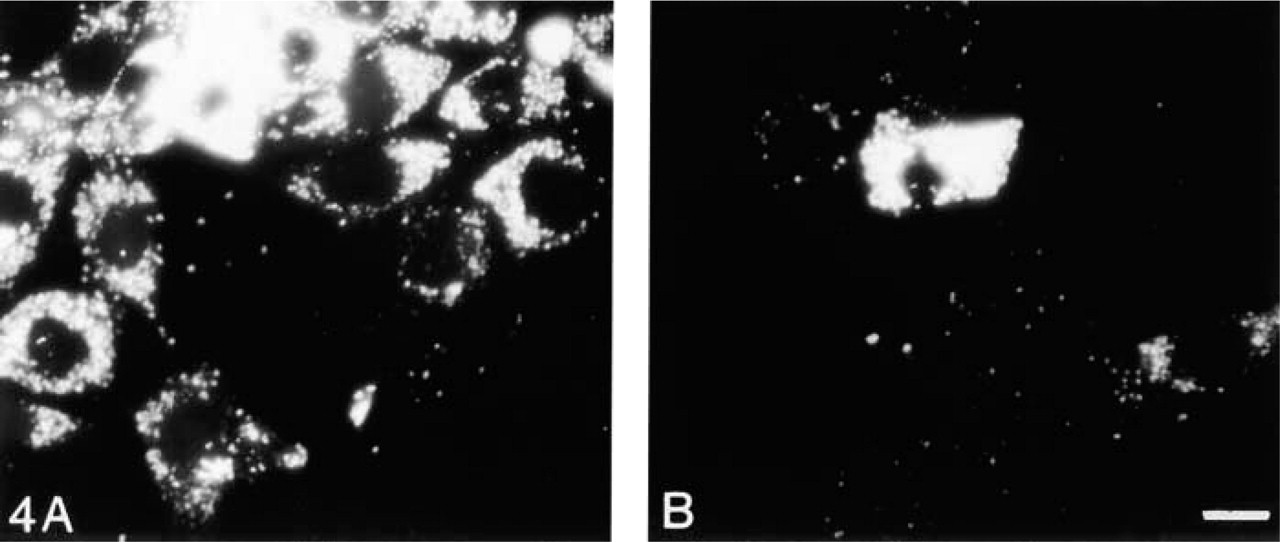
MCs express scavenger receptors and take up modified LDL by this pathway (Coritsidis et al. 1991). In keeping with those observations, we found that AcLDL was taken up by MCs. Because the amount of AcLDL uptake was comparable in control and IGF-1-treated MCs, IGF-1 did not appear to have an effect on scavenger receptor activity. This is consistent with previous studies in which IGF-1 failed to acutely alter expression or activity of scavenger receptors in smooth muscle cells (Gong and Pitas 1995). Although IGF-1 had no effect on AcLDL uptake, as with FITC-dextran, intracellular AcLDL persisted in IGF-1-treated cells for 7 days. This may be the result of IGF-1-mediated alterations in lipid efflux, late effects of IGF-1 on scavenger receptor activity, or secondary effects of IGF-1 that may stimulate the release of cytokines such as PDGF or TGFβ, which upregulate scavenger receptor activity (Gong and Pitas 1995). Additional studies are needed to distinguish between these possibilities.
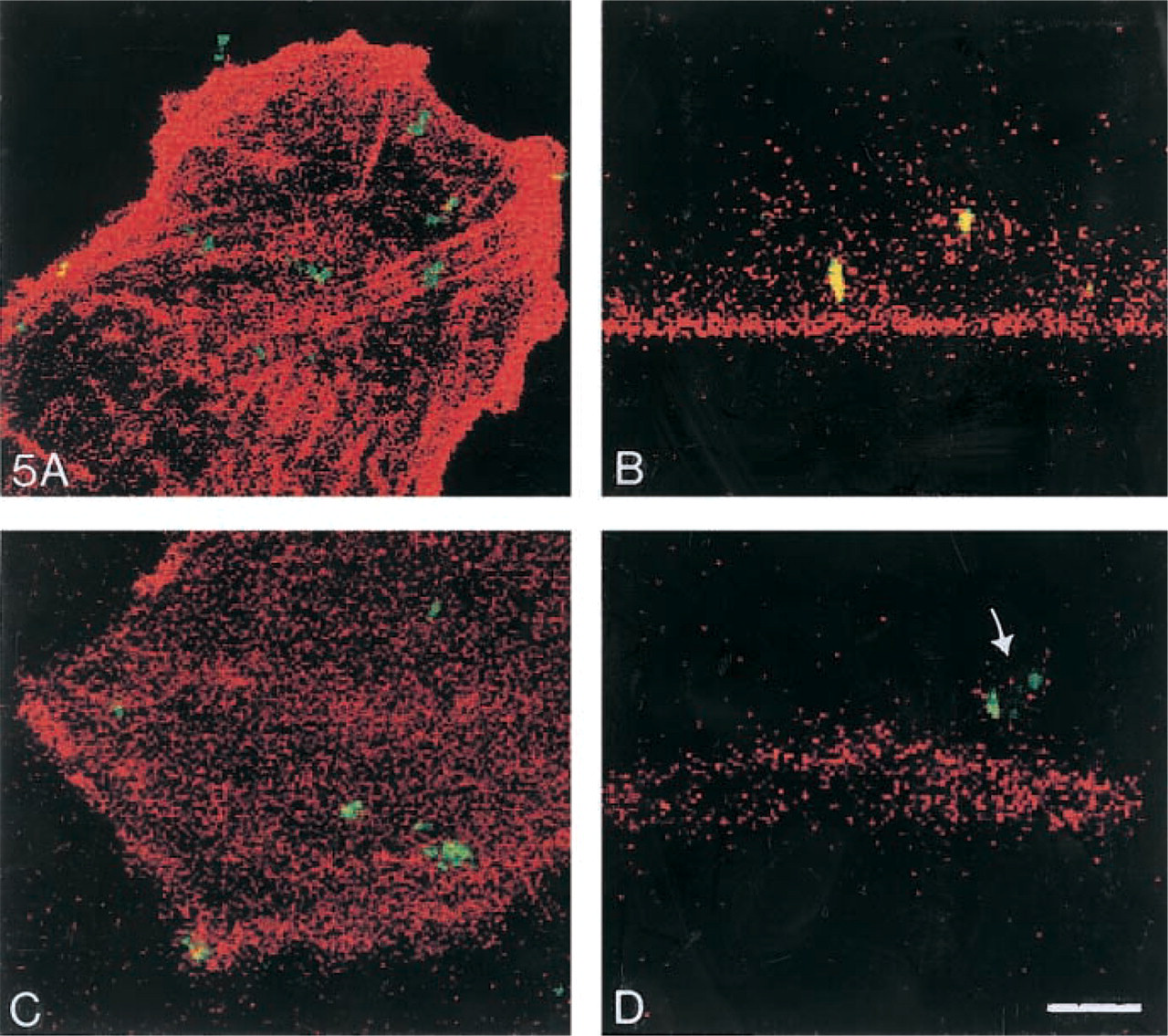
Confocal images of phagocytic MCs. In the MCs, f-actin and stress fibers were stained with Texas red phalloidin (red) while the E. coli was fluoresceinated (green). Untreated MCs (
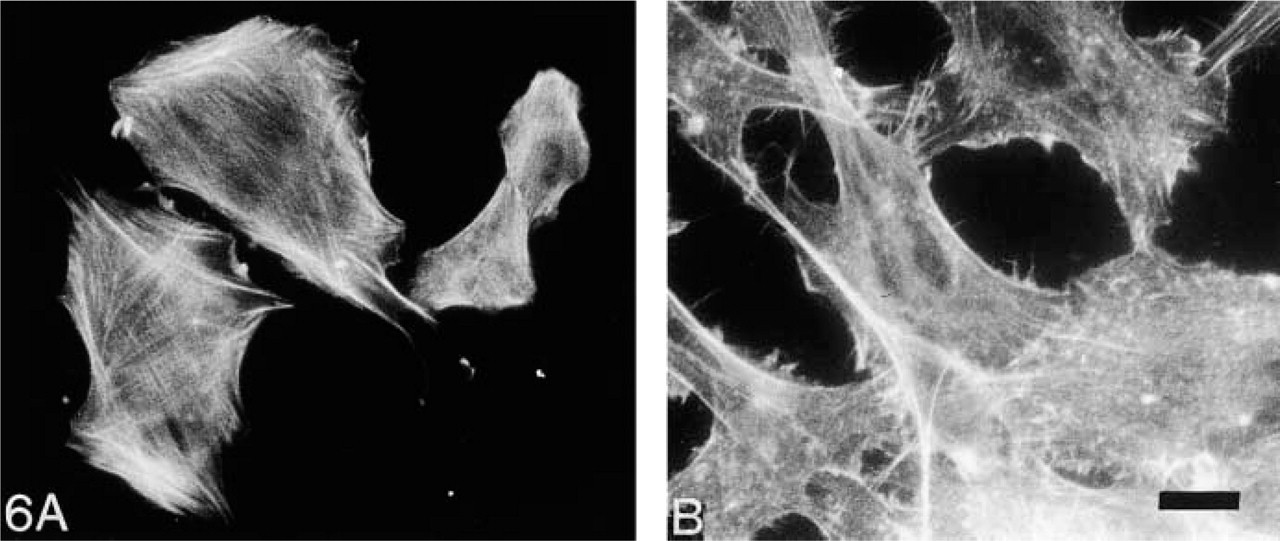
Cytoskeleton organization. Staining for f-actin in untreated MCs (
Long-term treatment of MCs with IGF-1 increased cholesteryl ester uptake compared to control MCs. Bo-dipy-labeled cholesteryl ester has classically been used to measure LDL receptor binding, which suggests that IGF-1 modulates LDL receptor expression. Recent data suggest that uptake of labeled cholesteryl ester may also define a “selective” pathway of vesicles or intracellular membrane sheets by which lipids are imported into the cell (Reaven et al. 1995). Although not well-defined, this pathway is similar to the endocytotic pathway in which substances are deposited on the cholesterol rafts of endosomes and are transported into the interior. Both insulin and IGF-1 decrease neutral cholesteryl esterase activity, thereby increasing cholesteryl ester accumulation in vascular smooth muscle cells and fibroblasts (Fujiwara et al. 1995), which may have contributed to lipid accumulation and foam cell formation in IGF-1-treated MCs. In summary, these studies show that IGF-1 stimulates an increase in endocytosis, which enhances lipid uptake. Lipid accumulation in IGF-1-treated MCs appears to be exaggerated by alterations in lipid degradation and efflux. Additional studies are needed to evaluate delayed effects of IGF-1 on scavenger and LDL-receptor function and to define the mechanisms whereby IGF-1 alters lipid egress from the cell.
Altered Phagocytosis and Cytoskeletal Reorganization
MCs are known to phagocytose both particulate matter and immune complexes (Hora et al. 1992), which mediate intraglomerular inflammation and limit glomerular injury. In previous short-term studies, IGF-1 treatment enhanced fluid-phase pinocytosis and membrane turnover (Berfield et al. 1997), which would be expected to enhance phagocytosis. However, when MCs become lipid-laden with long-term IGF-1 treatment, they are unable to completely phagocytose E. coli particles. During phagocytosis, lipid-laden MCs have increased membrane ruffling and partially enveloped particles. However, open connections to the exterior were demonstrated by the quenching of fluorescence by trypan blue. Failure of lipid-laden MCs to fully engulf E. coli was confirmed by confocal microscopy, which showed that E. coli particles were surrounded by actin protrusions (ruffles) but not engulfed. These observations suggest that the cytoskeleton cannot reorganize sufficiently to complete the closure of the phagocytic vesicle. Previously, we showed that IGF-1 induces rapid reorganization of the MC cyto-skeleton (Berfield et al. 1997), resulting in MCs with a bipolar phenotype as actin stress fibers reorganized from a criss-cross orientation to parallel alignment. After chronic (1 week) exposure to IGF-1, some cells retained this bipolar morphology. However, in many other cells actin was congealed and disorganized. The absence of f-actin stress fibers was correlated with the degree of lipid accumulation. Lipid and lipid oxides have previously been shown to disrupt actin microfilament organization (Palladini et al. 1996). During phagocytosis, an intact and responsive cytoskeleton is required to move the membrane around the particle, complete the zippering process, and draw the lyso-some into the interior of the cell (Swanson and Baer 1995). Therefore, it appears that the abnormalities in the cytoskeleton contribute to the inability of lipid-laden MCs to phagocytose E. coli particles.
Foam Cells and Renal Disease
Information supporting a role for lipids and progressive renal disease is emerging. Lipid deposits can be found in the glomeruli and renal interstitium of patients with focal and glomerulosclerosis, diabetic nephropathy, membranoproliferative glomerulonephritis, and hemolytic uremic syndrome (Lee et al. 1991; Moorhead 1991; Ravid et al. 1995; Lee 2000). High levels of circulating lipids accelerate the rate of progression of renal disease (Ravid et al. 1995), and treatment with low-fat diets or lipid-lowering agents slows the rate of progression of renal disease (Kasiske et al. 1988). In culture, lipids increase MCs release of cytokines, including monocyte chemoattractant protein-1 (Lynn and Siow 2000), and increase the rate of synthesis of extracellular matrix proteins (Feld et al. 1995; Neugarten and Schlondorff 1991). In turn, the extracellular matrix becomes a reservoir for binding lipids (Lee 2000), which may become oxidized. Oxidized lipoproteins are taken up by MC scavenger receptors, where they further contribute to foam cell formation (Coritsidis et al. 1991). Moreover, MCs oxidize LDL, which may promote renal injury (Rifici et al. 1994; Wheeler et al. 1994). Growth factors and cytokines act as co-factors in facilitating lipid-associated vascular injury (Bayes-Genis et al. 2000). MCs synthesize and respond to IGF-1 (Abrass et al. 1988; Conti et al. 1988; Berfield et al. 1997). Because IGF-1 synthesis is increased in certain diseases, particularly diabetic nephropathy (Elliot et al. 1993; Landau et al. 1995), IGF-1 action within the mesangium in the setting of hyperlipidemia may contribute to MC foam cell formation and impairment of MC function. MCs are similar to vascular smooth muscle cells in handling of and response to lipids. Therefore, the mechanism responsible for lipid-mediated injury may be similar in atherosclerosis and some forms of glomerulosclerosis.
Footnotes
Acknowledgements
Supported by the Medical Research Service of the Department of Veterans Affairs.