
Abstract
Substance P contributes to the physiological homeostasis of pulmonary airways and vasculature. During pneumonia, alterations in substance P production and receptor expression can influence bronchoconstriction and vascular perfusion. The distribution of substance P receptor [neurokinin-1 receptor (NK-1R)] in lungs of normal sheep and sheep with acute (1 day), subacute (15 days), and chronic (45 days) bronchopneumonia caused by Mannheimia haemolytica was determined by immunohistochemistry (IHC). Three rabbit polyclonal antibodies generated to the same cytosolic C-terminal portion of NK-1R (residues 393-407) were tested. NK-1R immunoreactivity was traced in digital images and quantified with IPLAB software. There were no significant differences in NK-1R protein density between normal and infected lambs. Antibody 1 had the broadest distribution and intensity, and stained alveolar septae, smooth muscle cells of airways and vessels, epithelial cells of airways and alveoli, and submucosal glands. When all animals from the study were included, there was a trend towards decreased NK-1R immunoreactivity over time. The work suggests that (a) the density of NK-1R does not change during progression of bacterial (M. haemolytica) bronchopneumonia, (b) NK-1R is widely distributed in ovine lung and decreases with age, and (c) antibodies to the same NK-1R cytosolic region can vary in specificity and affinity.
T
Among TKs, SP is considered to be the main sensory neurotransmitter in mammalian airways (Corcoran 1996). The distribution of capsaicin-sensitive SP-like immunoreactive (SP-Li) nerve fibers in the respiratory tract has been examined in cattle (Nishi et al. 2000), sheep (Corcoran 1996; Ramirez-Romero et al. 2000,2001), and horses (Sonea et al. 1994a,b). These studies indicate that SP-Li nerve fibers are much more numerous in young animals than in adults and are mainly seen in the epithelium (cattle, horses), in connective tissue beneath the epithelium and around blood vessels (cattle, horses, sheep), submucosal glands (cattle, horses, sheep), airway smooth muscles (neonatal foals, sheep), and bronchial and pulmonary vessels (horses). A similar distribution has been found in the human respiratory tract (Bai et al. 1995). Together with NKA, SP produces a proinflammatory effect, that contributes to the physiological homeostasis of pulmonary airways and vasculature and is called “neurogenic inflammation” (Geppetti and Holzer 1996). It stimulates mucus secretion from submucosal glands and goblet cells (Rogers 2000), mucociliary clearance (Wong et al. 1990), plasma protein extravasation (Gamse and Saria 1985; Keith 2000), vasodilatation (Pedersen et al. 2000), leukocyte adhesion to the vascular endothelium (Matis et al. 1990), alveolar macrophage and monocyte release of inflammatory cytokines, and mast cell degranulation (Barnes 2001). SP also enhances cholinergically mediated broncho-constriction, angiogenesis, and proliferation and chemotaxis of human lung fibroblasts (Barnes 2001; Rice et al. 2001). However, there are some exceptions, such as SP-induced contraction of pulmonary arteries in rabbits (Shirahase et al. 1995) or relaxation of bronchial smooth muscles in rats (Bodelsson et al. 1999). These species differences might be attributed to different pathways (direct and indirect) of neurokinin receptor activation (Bodelsson et al. 1999; Amadesi et al. 2001) and distribution, indicating the precautions needed during interpretation of interspecies diversity.
Three mammalian NK receptors have been cloned from several species, and including NK-1R, NK-2R, and NK-3R (Joos et al. 1994). All belong to the super-family of guanine nucleotide-binding, protein-coupled receptors, with seven putative transmembrane-spanning α-helices, an extracellular amino-terminus, and an intracellular carboxyl tail (Piedimonte 1995; Joos et al. 2000). Although, when present at sufficiently high concentrations, each of the naturally occurring TKs can act as a full agonist on all three receptors (Regoli et al. 1987), SP has the highest binding affinity for NK-1R (Regoli et al. 1988). In human subjects, positive immunostaining for NK-1R is present in the bronchial smooth muscles, myoepithelial cells of bronchial glands, in the endothelium and smooth-muscle layer of bronchial vessels and pulmonary arteries, and occasionally in nerves (Mapp et al. 2000). NK-1R is also present in the apical portion of bronchial epithelium (particularly in goblet cells) and in some inflammatory cells (Chu et al. 2000). In rat lung, NK-1R is present in the apical portion of the airway epithelium, from the trachea to the respiratory bronchioles, but not in alveoli, submucosal glands, or smooth muscle of the airways or most blood vessels, except for the smooth muscle of some intermediate-sized pulmonary veins (Ichikawa et al. 1995). In contrast to rats, guinea pig lung has NK-1R immuno-reactivity in smooth muscle from trachea to small bronchioles and from pulmonary vessels (Carstairs and Barnes 1986). These studies underscore the lack of complete correlation between the anatomic distribution of receptors and SP-like nerve fibers in the lung of various species, indicating the importance of revealing the distribution of NK-1R, particularly in attempting to understand the neuropeptide's mechanisms of action and functions.
Corcoran and Haigh (1992) have suggested that NK-1R predominates in sheep airways compared to other NK receptors. Therefore, the first aim of our study was to assess NK-1R distribution in the lung of healthy weaned lambs and to compare its immunoreactivity with various stages of pneumonia caused by the ovine pathogen M. haemolytica (Ramirez-Romero et al. 2001) during the 45-day period. Our second objective was to test the distribution of immunoreactivity of three rabbit polyclonal antibodies generated against a synthetic NK-1R peptide identical to a 15-amino-acid section of the cytosolic C-terminal portion of NK-1R (residues 393-407) in ovine lung.
Materials and Methods
Experimental Animals
Nineteen sheep of both sexes and mixed breed, 3 months old (weaned), were obtained from Iowa State University Laboratory Animal Resources. The lambs were randomly assigned to two groups, each group being maintained in a separate climate-controlled isolation room until sacrifice. After a 24-hr adaptation period, one group (nine animals) received saline and the other (10 animals) received Mannheimia (Pasteurella) haemolytica. The lambs were subdivided into six groups according to the time of euthanasia at day 1, 15, or 45 after inoculation (Table 1). This protocol was approved by the Iowa State University Animal Care Review Committee.
Inoculation
M. haemolytica serotype A1 ovine strain 82-25 was obtained from a natural case of pneumonic pasteurellosis in sheep. The animals received 5 ml of either bacterial inoculum containing 1.9 × 108 cfu/ml (Brogden et al. 1995) or pyrogen-free saline (PFS). Intrabronchial inoculation was performed using fiberoptic bronchoscopy as previously described (Brogden et al. 1984,1995). Briefly, in non-intubated, sedated lambs, the bronchoscope (model VFS-2; Schott Fiber-optics, Southbridge, MA) was inserted via the nasotracheal route. The inoculum was deposited into the main bronchus of the right cranial lobe.
Experimental design
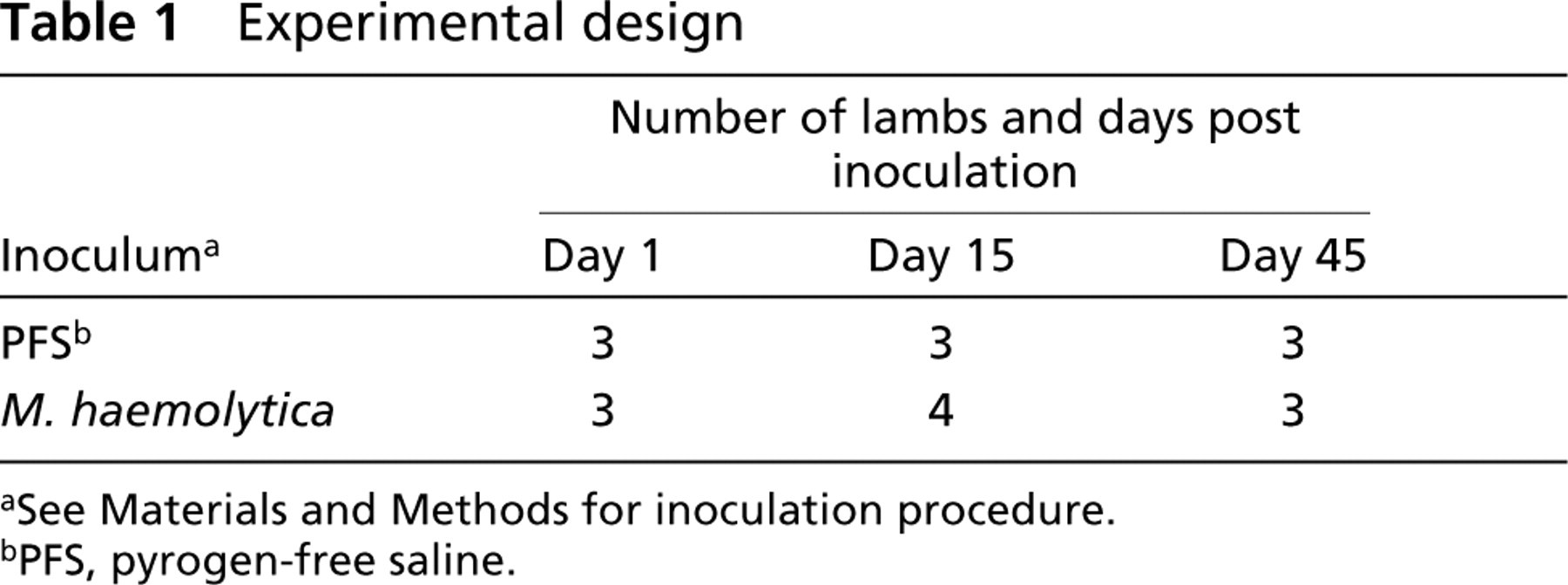
aSee Materials and Methods for inoculation procedure.
bPFS, pyrogen-free saline.
Tissue Collection and Processing
The animals were sacrificed at day 1, 15, or 45 after inoculation with an IV (intrajugular) overdose of sodium pentobarbital. At necropsy, samples were taken from the pulmonary deposition site (right cranial lobe). One sample was taken proximal to the hilus and included the cranial bronchus. The other one was from the distal part and included the apex of the cranial lobe (Ramirez-Romero et al. 2001). Samples were fixed by immersion in 10% neutral buffered formalin [48 hr at room temperature (RT)] and replicate sections were stained with hematoxylin-eosin to assess the microscopic lesions of the lung samples, or stained by immunohistochemistry (IHC) for localization of the NK-1 receptor in the same areas of lung.
Immunohistochemistry
Sections were stained with antibodies to the NK-1 receptor using the streptavidin-biotin complex peroxidase method, and the metal-enhanced diaminobenzidine (DAB) method was used to reveal the peroxidase activity as previously described (Shu et al. 1988). Briefly, the slides were heated in the oven (58C) for 30 min, deparaffinized in xylene, immersed through a succession of graded alcohols, and then fully hydrated. Next they were immersed in 1% H2O2 in ultrapure water for 40 min to block endogenous peroxidase activity. After several rinses in BioGenex PBS at pH 7.2-7.4 containing 0.1% Tween-20 (Wash Buffer; BioGenex, San Ramon, CA), nonspecific binding was blocked by incubation in 10% normal goat serum (NGS; Sigma, St Louis, MO) and 3% bovine serum albumin (essentially γ-globulin-free BSA; Sigma A-7030) in BioGenex PBS for 20 min. The sections were then placed in a humidified container and incubated with primary antibodies for 18 hr at 4C. The primary antibodies used in this study were polyclonal rabbit anti-NK-1R peptide, in the form of hyperimmune serum, diluted 1:4000 (antibody 1 from Oncogene rabbit #653), 1:3000 (antibody 2 from Oncogene rabbit #652), and 1:3000 (antibody 3 from Oncogene rabbit #654) in antibody diluent (Common Antibody Diluent; BioGenex). Then the slides were placed on the OptiMax Plus automated cell-staining machine (BioGenex) to complete the remaining procedures. The machine washed the slides with BioGenex PBS, applied 2% H2O2 in BioGenex PBS for 20 min, and then washed with BioGenex PBS again. The sections were then preincubated briefly (30 sec) with a biotinylated goat anti-rabbit secondary antibody and then, with no rinse in between, the same reagent was applied for 50 min (ready-to-use human serum-adsorbed biotinylated goat anti-rabbit already at 2 μg biotinylated IgG/ml; Kirkegaard & Perry Laboratories, Gaithersburg, MD). After several rinses in BioGenex PBS, the sections were incubated for 50 min at RT with a streptavidin-HRP complex (Super-Sensitive peroxidase-conjugated streptavidin; BioGenex). Finally, the slides were washed with BioGenex PBS and exposed for 12 min to a metal (cobalt chloride and nickel chloride)-enhanced DAB substrate (ImmunePure Metal Enhanced DAB Substrate Kit; Pierce Chemical, Rockford, IL) to visualize immunoreactivity (IR). The slides were then washed thoroughly with ultra-pure water to stop the DAB reaction, counterstained for 2 min with quarter-strength Shandon's acidified Harris haematoxylin, subjected to Scott's tapwater (10 g MgSO4 and 2 g NaHCO3 per liter of water) for 1 min, dehydrated through graded alcohols, cleared in xylene, and coverslipped with Permount (Sigma). Dark brown precipitate was accepted as a positive reaction. Control sections lacked staining and included (a) no primary antibody, (b) normal rabbit serum, (c) normal rabbit IgG (Upstate Biotechnology; Lake Placid, NY), and (d) NK-1R antibody preincubated with synthesized NK-1R peptide.
Primary Antibodies
Antisera tested in this study were purchased from Oncogene (Oncogene Research Products, a division of CN Bioscience; San Diego, CA). They were raised in three rabbits (#652, #653, and #654 at Oncogene) in response to the peptide KT-MTESSSFYSNMLA, which corresponds to a 15-amino-acid section (residues 393-407) of the cytosolic carboxy-terminal region of the NK-1 receptor. The peptide was crosslinked via glutaraldehyde to bovine thyroglobulin. These hyperimmune rabbit sera were tested for specificity to NK-1R by preincubating them for 45 min with different concentrations of a synthesized 15-mer peptide (KTMTESSSFYSNMLA), which was identical to the peptide used by Oncogene (now Calbiochem) to immunize the rabbits. This peptide was synthesized by automated solid-phase peptide synthesis and purified by reversed-phase high-pressure liquid chromatography (Joel Nott; Protein Facility, ISU, Ames, IA). A minimum of 33.93 μg/ml (20 μM) concentration of this peptide completely inhibited IHC staining by all three rabbit anti-NK-1R IgGs.
Mean values for the area of neurokinin 1 receptor (NK-1R) protein expression in the lung of sheep inoculated with PFS or M. haemolytica at 1, 15, and 45 days post inoculation (p.i.)
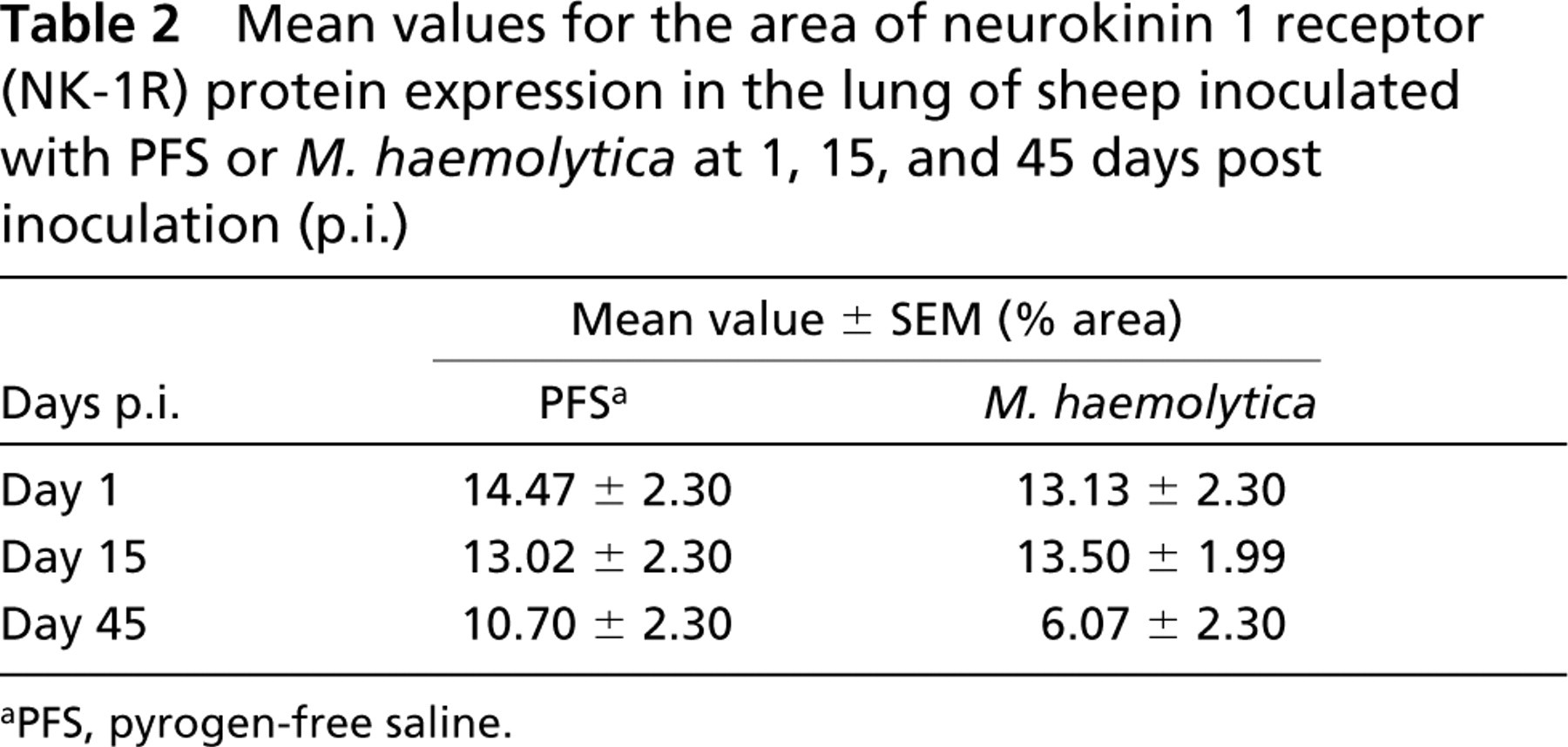
aPFS, pyrogen-free saline.
Mean values for distribution of neurokinin 1 receptor (NK-1R) protein in sheep lung during the period of 45 days; values for all three primary antibodies were pooled
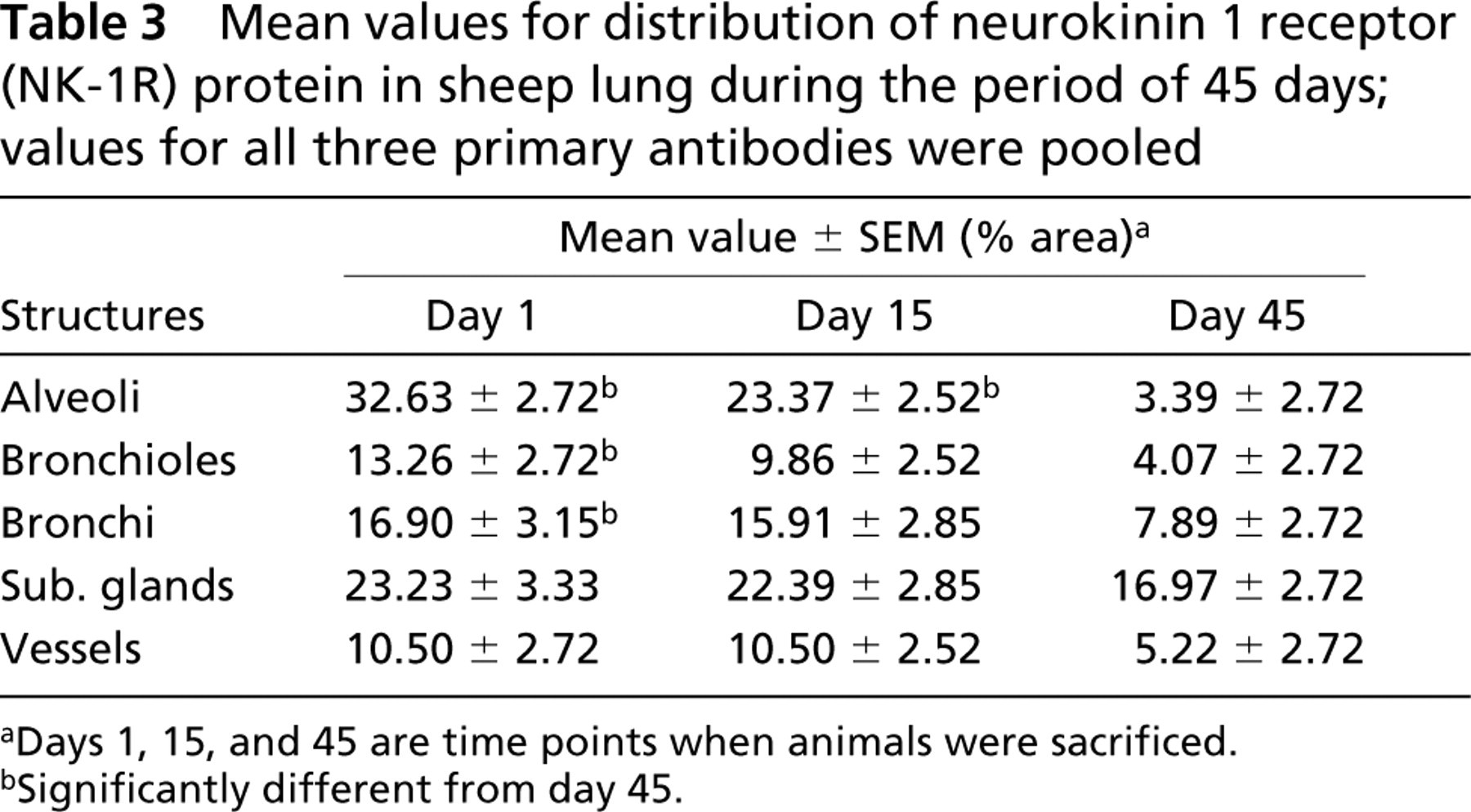
aDays 1, 15, and 45 are time points when animals were sacrificed.
bSignificantly different from day 45.
Image Analysis for NK-1R Immunoreactivity
The structures examined in the lung samples included alveoli, bronchi, bronchioles, submucosal glands, and vessels. Specific compartments observed within bronchi and bronchioles included lamina epithelialis mucosae, lamina propria mucosae, and lamina muscularis mucosae, while vessels included tunica intima and media as one compartment and adventitia as the other. Alveoli and submucosal glands were assessed as single compartments. Three fields of each structure, chosen randomly from every slide, were examined with a Zeiss Axioskop fitted with Neoflur objectives (×20 objectives), a 100-W light source, and a Sony DXC-3000A camera. The images were then captured with IPLAB Software (Scanalytics; Fairfax, VA) on a Macintosh G3 (Apple; Cupertino, CA). Images were edited on a Micron workstation with Photoshop (Adobe Photoshop 6.0; Mountain View, CA), thresholded for analysis, and quantified on the G3 with the IPLAB software.
Statistical Analysis
Results represent percentages of stained areas over total areas of randomly picked lung structures/compartments. Data are expressed as means ± SEM of three randomly chosen fields from each structure/compartment for each of the assessed animals. Differences in means between three tested antibodies, day, treatment, and structure/compartment effects were analyzed using two-factor analysis of variance (ANOVA) test. All analyses were performed using SAS software. Values were considered to be significant at p<0.05.
Mean values for total area of neurokinin 1 receptor (NK-1R) protein expression in sheep lung during the period of 45 days; three primary antibodies were compared
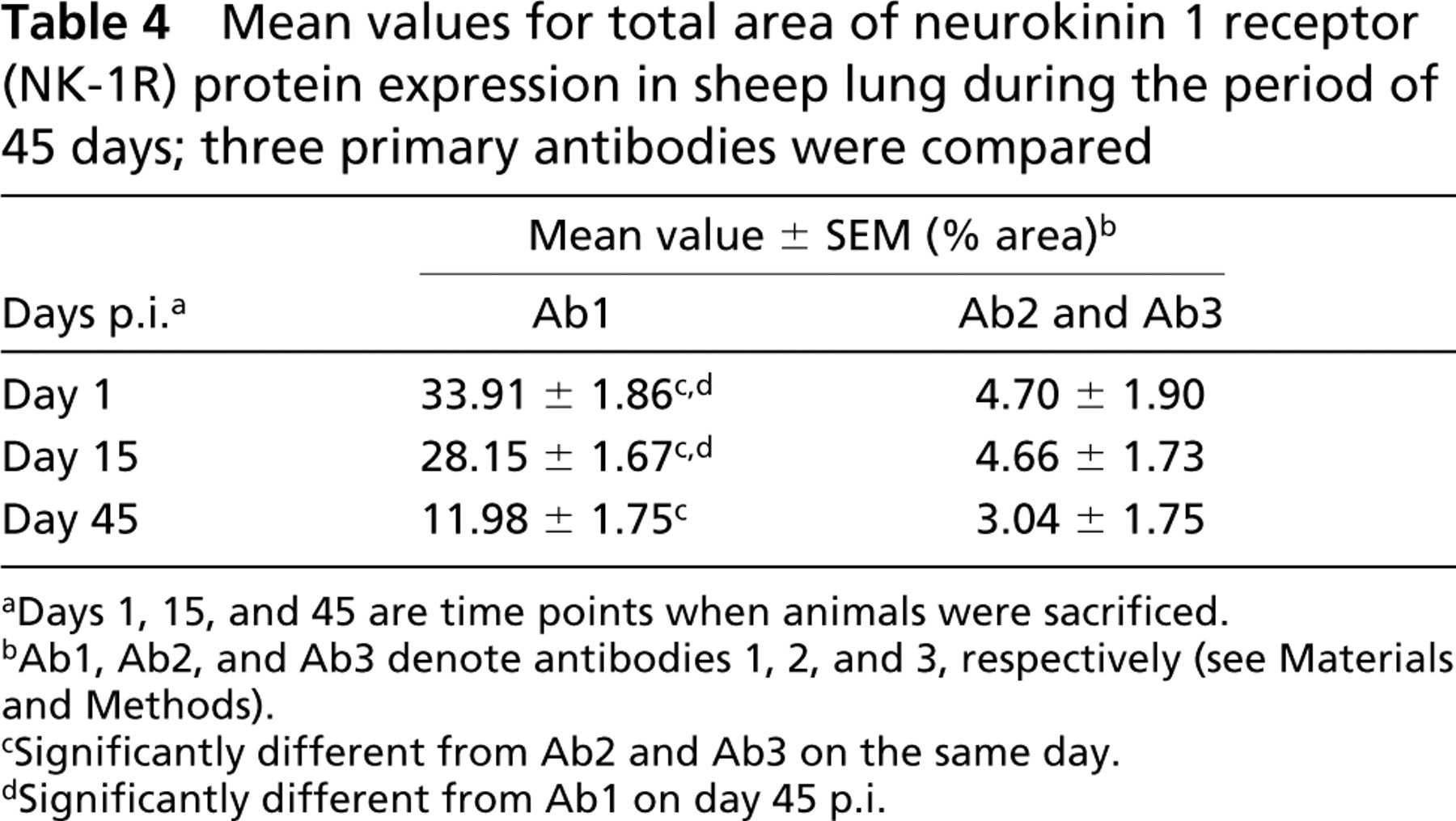
aDays 1, 15, and 45 are time points when animals were sacrificed.
bAb1, Ab2, and Ab3 denote antibodies 1, 2, and 3, respectively (see Materials and Methods).
cSignificantly different from Ab2 and Ab3 on the same day.
dSignificantly different from Ab1 on day 45 p.i.
Results
Microscopic Lesions
Animals inoculated with M. haemolytica developed severe lesions at the site of inoculum deposition and have been described previously (Ramirez-Romero et al. 2001). Briefly, in animals sacrificed at 1 day after inoculation with bacteria, lungs had extensive areas of seroproteinaceous fluid, neutrophil infiltration, hemorrhage, and necrosis. Lungs of the animals at 15 days after inoculation were characterized by fibrosis of alveolar and interlobular septa and pleura, marked proliferation of type II alveolar cells, many alveolar macrophages within the alveoli, and occasional foci of pyogranulomatous inflammation. Finally, at 45 days after inoculation, lungs had epithelial cell proliferation in alveoli and bronchioles as well as marked pleural and interlobular fibrosis. In addition, there was increased tortuosity of the small airway lumens that were partially obliterated by fibrous connective tissue and occasional areas of alveolar atelectasis or emphysema. All control animals, inoculated with saline, lacked lesions.
Immunoreactivity of Primary Antibodies
IHC for NK-1R was repeated on the same set of lung tissues using three different polyclonal antibodies, each of which was produced in a different rabbit but to the same region of NK-1R. Control sections lacked staining and included no primary antibody, normal rabbit serum from Sigma, normal rabbit IgG, and NK-1R antibody preincubated with a synthetic 15-aa peptide of NK-1R residues 393-407.
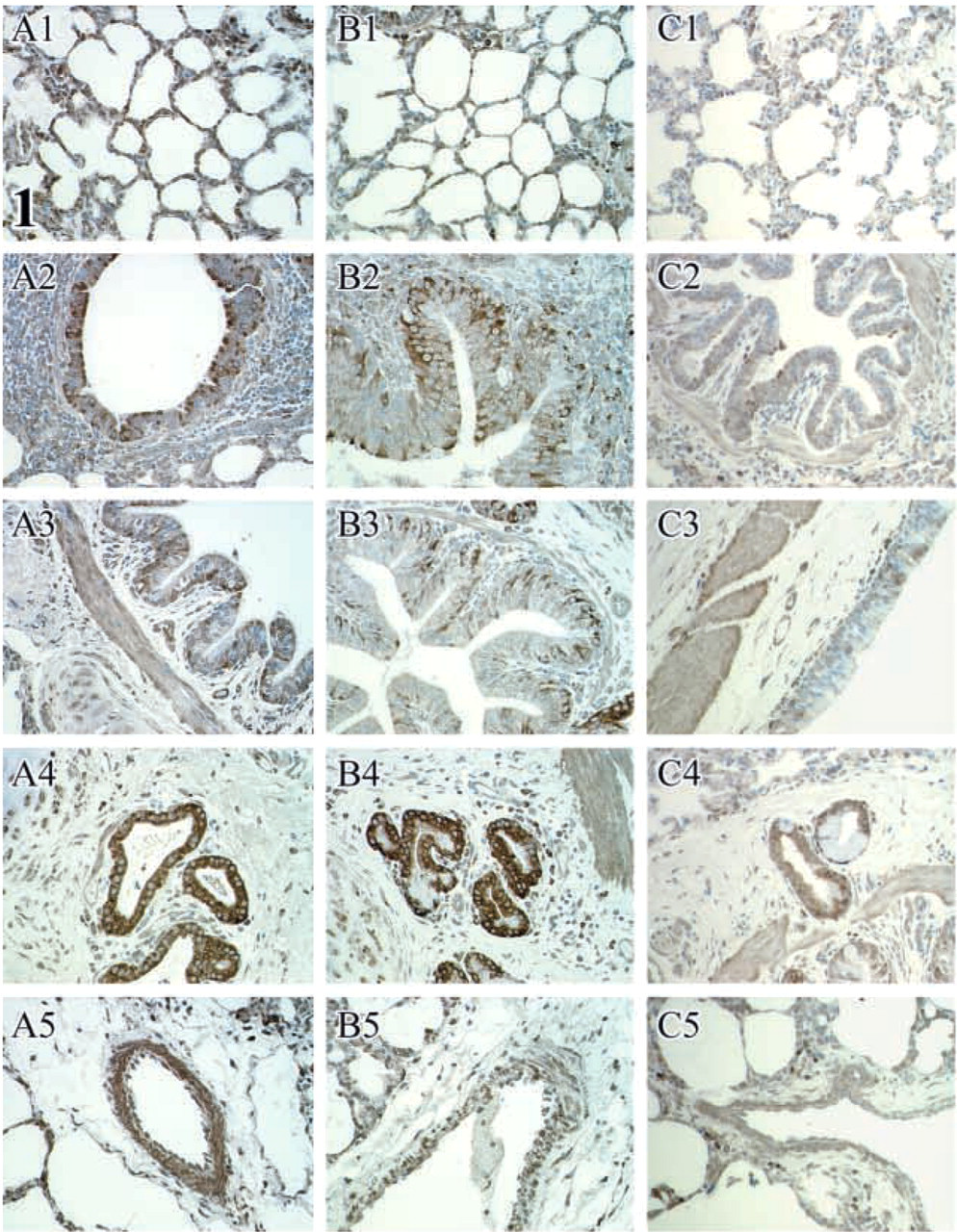
NK-1R immunoreactivity in lung tissues stained with Ab1 on day 1 (
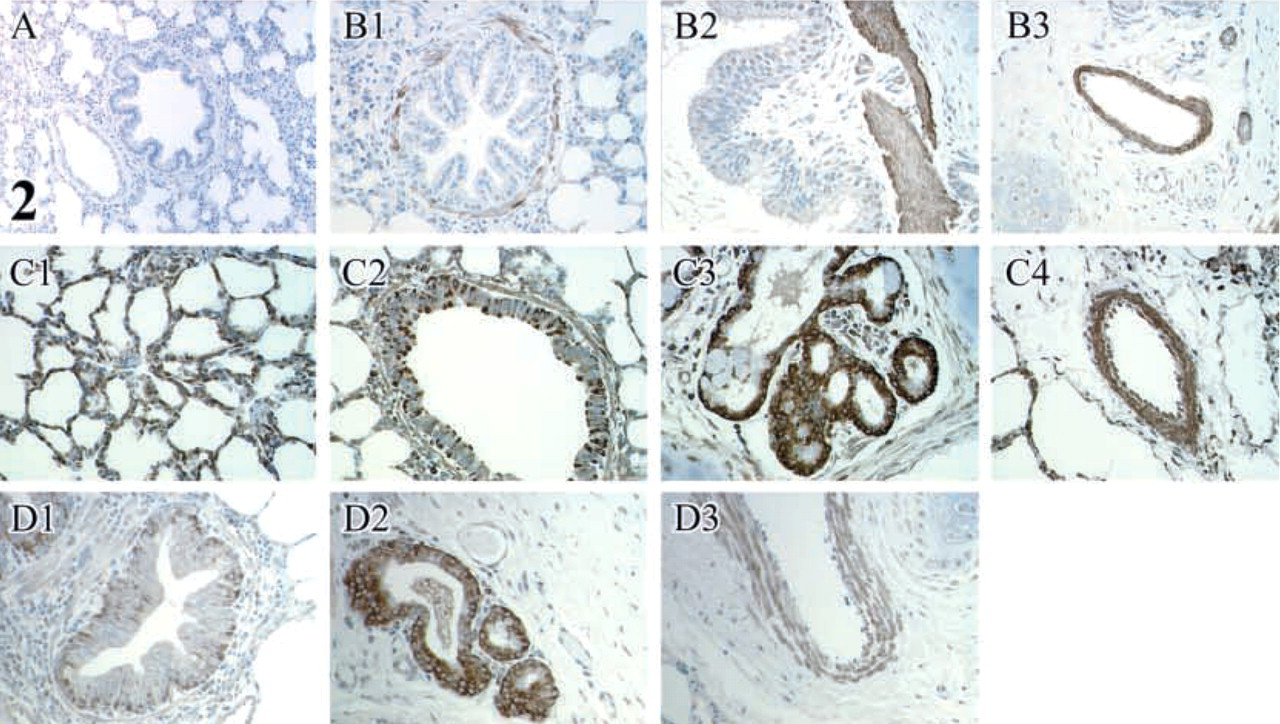
(
Quantitative Image Analysis of NK-1R
The NK-1R immunoreactivity was widely distributed in ovine lung. However, we did not see any difference in the amount of NK-1R protein expression when control (PFS) and infected (M. haemolytica) animals were compared at day 1, 15, and 45 p.i. (Table 2). Because of this finding, all the values from control and infected animals were pulled together and further analyzed. The area of NK-1R immunoreactivity were greatest at 1 day after inoculation and progressively decreased at 15 and 45 days after inoculation (Table 3). However, this progressive decrease in immunoreactivity was significant only for Ab1 (p<0.05) (Table 4; Figure 1).
When immunoreactivity for three primary antibodies was compared, the area of immunoreactivity for Ab1 was significantly greater (p<0.05) than for Ab2 and Ab3 in all the assessed structures (alveoli, bronchioles, bronchi, submucosal glands, and vessels) (Table 5; Figure 2). The area of immunoreactivity for Ab1 was significantly greater in alveoli and submucosal glands of bronchi (p<0.05) than in bronchioles, bronchi, and blood vessels. In contrast, the areas of immunoreactivity for Ab2 and Ab3 were significantly greater in bronchi, submucosal glands of bronchi, and in blood vessels (p<0.05) than in alveoli and bronchioles. Within bronchi and bronchioles, the area of immunoreactivity was greatest in the lamina muscularis and lowest in the lamina propria for all three antibodies (p<0.05) (Table 6). The area of immunoreactivity was significantly greater in lamina muscularis as well as the bronchial and bronchiolar epithelium for Ab1 (p<0.05) compared to Ab2 and Ab3 (Table 6). Within the vessels, the area of immunoreactivity was significantly greater in the tunica media than in the tunica adventitia for all antibodies (p<0.05) (Table 7).
Mean values for the area of neurokinin 1 receptor (NK-1R) protein expression in various regions of lung
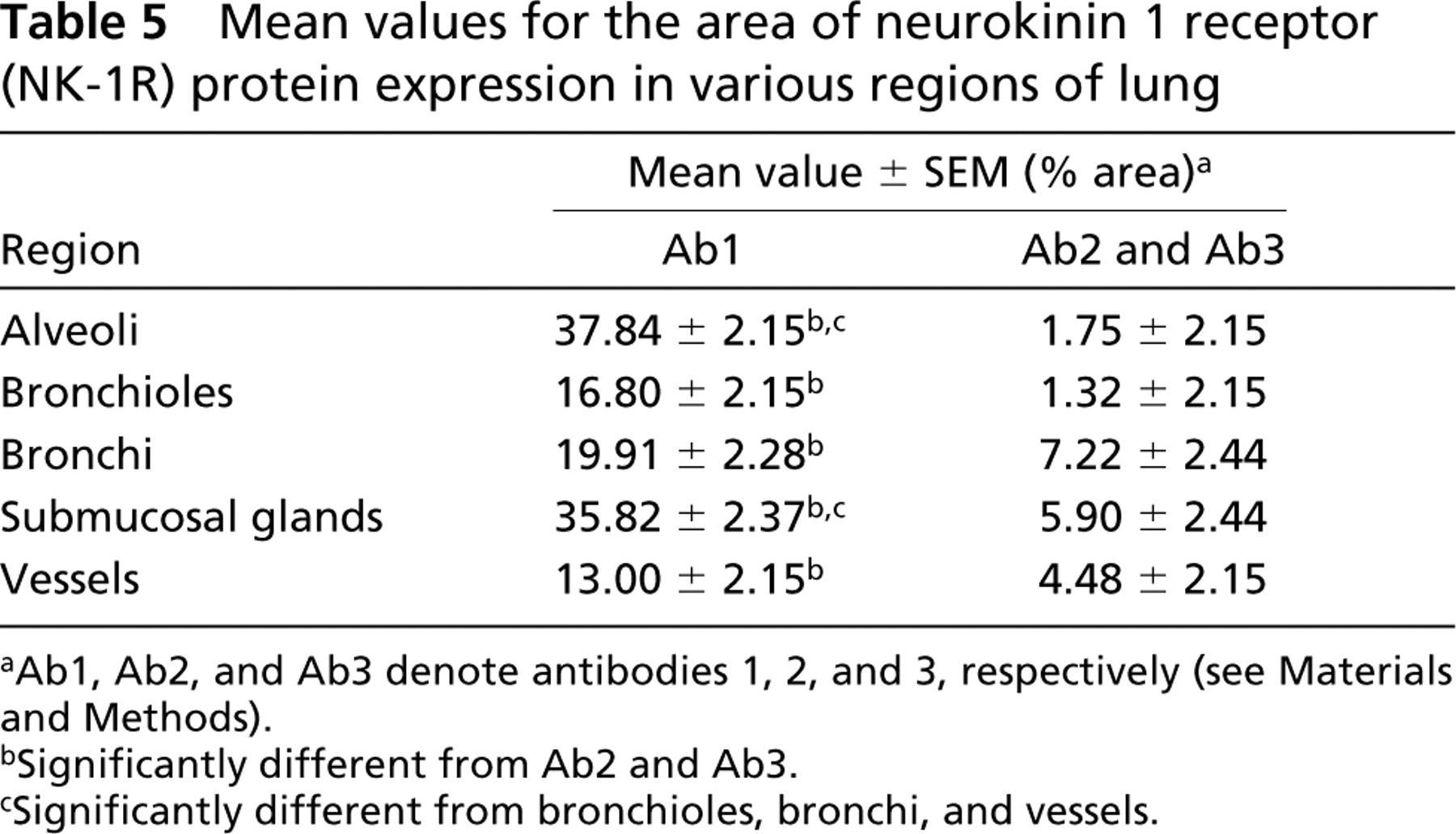
aAb1, Ab2, and Ab3 denote antibodies 1, 2, and 3, respectively (see Materials and Methods).
bSignificantly different from Ab2 and Ab3.
cSignificantly different from bronchioles, bronchi, and vessels.
Mean values for the area of neurokinin-1 receptor (NK-1R) protein expression in the bronchial and bronchiolar compartments
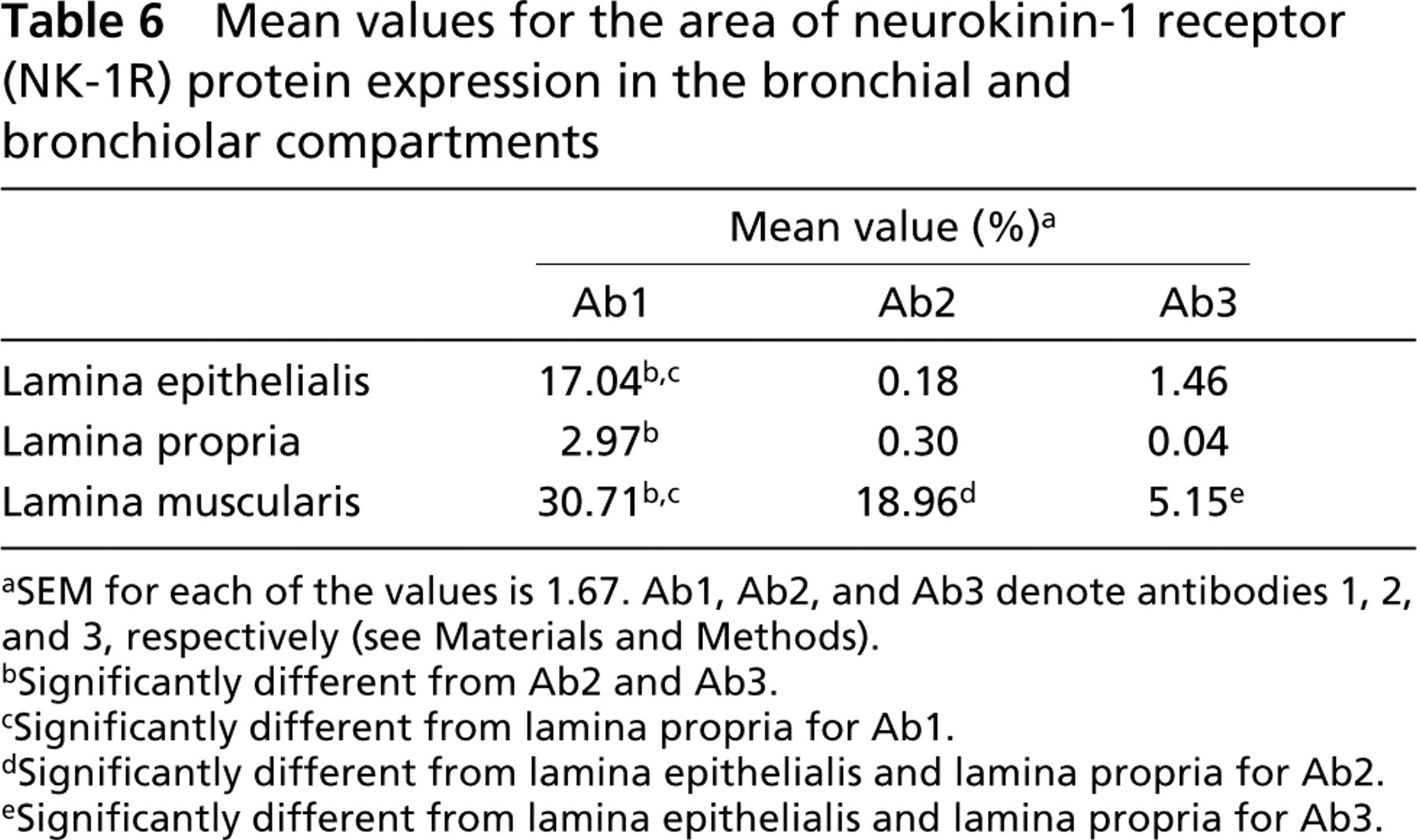
aSEM for each of the values is 1.67. Ab1, Ab2, and Ab3 denote antibodies 1, 2, and 3, respectively (see Materials and Methods).
bSignificantly different from Ab2 and Ab3.
cSignificantly different from lamina propria for Ab1.
dSignificantly different from lamina epithelialis and lamina propria for Ab2.
eSignificantly different from lamina epithelialis and lamina propria for Ab3.
Discussion
The findings regarding the amount of NK-1R mRNA and protein expression in the lung vary greatly. One study shows that, in asthmatics and patients with COPD, NK-1R mRNA levels are unchanged compared with non-smoking controls but increase twofold in smokers (Bai et al. 1995). Conversely, Chu et al. (2000) demonstrated that asthmatic subjects have increased expression of SP and NK-1R protein in their airway epithelium but not in the submucosa, compared with normal control subjects. In our study, no differences in NK-1R protein densities were note on comparing control to infected lambs. The reason for that may be that the extent of SP production does not change during the M. haemolytica infection. Even though the density of SP-Li nerve fibers tends to decrease during the initiation and at 45 days of M. haemolytica infection, SP production appears increased in macrophages and neuronal ganglion cells during infection (Ramirez-Romero et al. 2001). Therefore, up-regulation of NK-1R appears to be dependent on SP, which is in agreement with studies performed on rat intestine (Pothoulakis et al. 1998). However, we cannot exclude the possibility that the overall level of NK-1R remains unchanged, i.e., any increase in SP binding could be due to an increased presence of NK-1R during pulmonary immune responses resulting from the influx of receptor-bearing leukocytes which, as of yet, have not been assessed by IHC.
Mean values for the area of neurokinin-1 receptor (NK-1R) protein expression in the vascular compartments
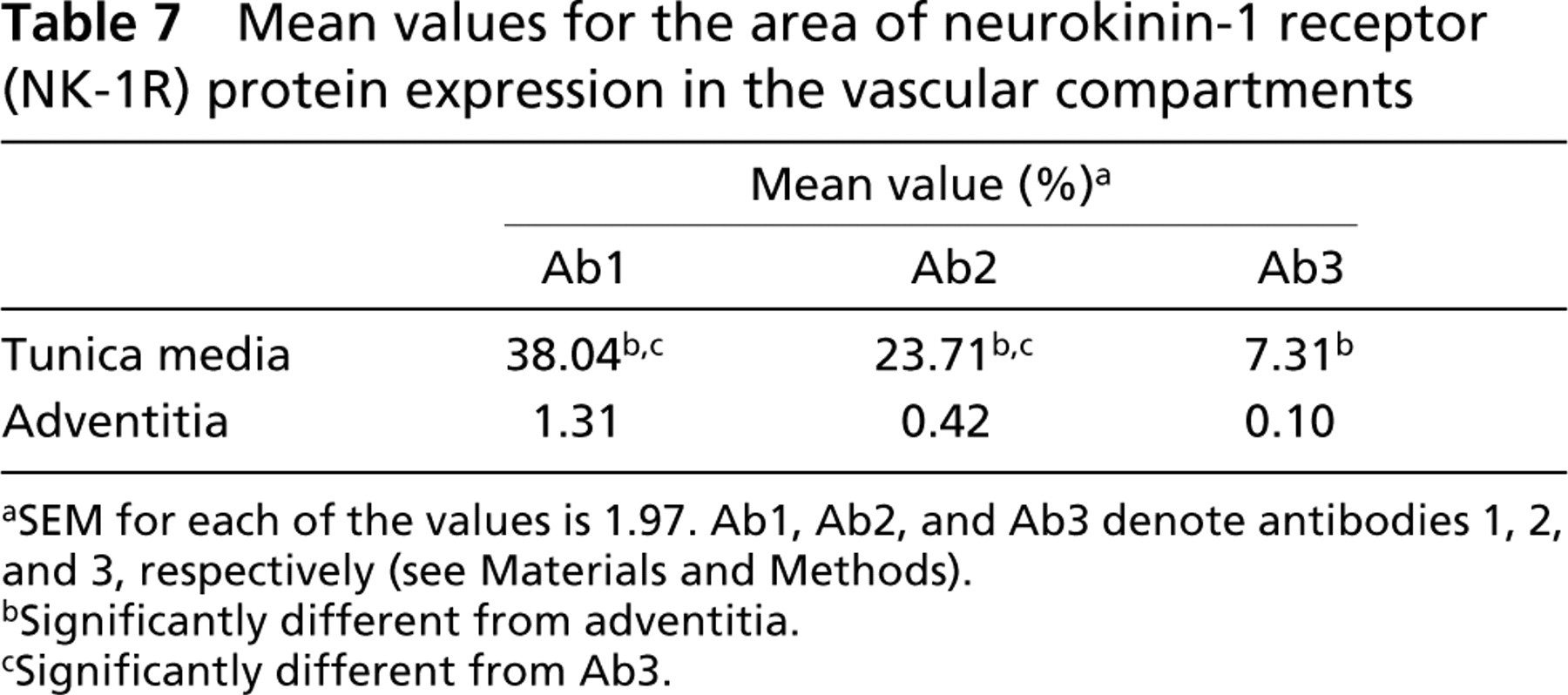
aSEM for each of the values is 1.97. Ab1, Ab2, and Ab3 denote antibodies 1, 2, and 3, respectively (see Materials and Methods).
bSignificantly different from adventitia.
cSignificantly different from Ab3.
Our data suggest that NK-1R is widely distributed in healthy ovine lung. These results were expected because it was demonstrated that, when injected IV into sheep, SP is a more potent bronchoconstrictor than NKA (Corcoran and Haigh 1992; Rice et al. 2001). It is also likely that the density of pulmonary NK-1R decreases with age (at least for Ab1). Progressive loss of SP-Li nerve fibers during postnatal maturation of the respiratory tract was found in the foal lung (Sonea et al. 1994b), human bronchi (Hislop et al. 1990), and the nasal septum of lambs (Ramirez-Romero et al. 2001). Furthermore, McLeod et al. (1998) demonstrated that the anatomic distribution of the SP-containing fibers and NK-1R in the rat spinal cord correlate well. When combined, these results favor our hypothesis that the density of both the SP-Li nerve fibers and NK-1R decreases with aging in the sheep lung. It has also been shown that NK-1R -/- mice lack generalized immune complex-mediated lung injury compared to wild-type NK-1R +/+ animals (Bozic et al. 1996). Higher numbers of NK-1R in young animals could increase their susceptibility to inflammatory responses.
Localization and characterization of tachykinin receptors have been extensively studied using various morphological and pharmacological approaches (Joos et al. 2000). In the present study, applying a standard IHC procedure, we have evaluated three “different” polyclonal antibodies generated to the same antigen (NK-1R cytosolic region, residues 393-407), whose density and expression have been shown to be related to the NK-1R protein expression (Vigna et al. 1994). After testing these antibodies for their specificity and affinity, we thought that Ab1 was the most specific in recognizing the antigen of interest, for two reasons. First, Ab2 and Ab3 recognized only some of the targets stained by Ab1 with a lower intensity than stained by Ab1. Second, there was no background staining or staining of the structures labeled by Ab1, Ab2, or Ab3 when normal rabbit serum or NRS IgG was used instead of the polyclonal antibodies for the IHC procedure as negative controls. These findings are not unusual because it is well known that antisera produced in response to immunization are a mixture of a heterogeneous population of antibody molecules (Janeway et al. 1999). This heterogeneity can be due to production of antibodies to different epitopes on the immunizing agent (bovine thyroglobulin) as well as to different spots on the single antigenic determinant, such as hapten (peptide in our study).
This work demonstrates the following. (a) The density of NK-1R is unchanged during progression of bacterial (M. haemolytica) bronchopneumonia. (b) NK-1R is widely distributed in ovine lung and decreases with age. (c) Antibodies to the same NK-1R cytosolic region can vary in specificity and affinity. By revealing the precise location and distribution of NK-1R in the lung, selective NK-1R antagonists can be applied more effectively as potential inhibitors of mucus hypersecretion (MEN 11467; Khan et al. 2001) and plasma extravasation (CP 99,994; Ball et al. 1993), FK 888 (Murai et al. 1993). Non-selective antagonists such as FK 224, which are active on both NK-1 and NK-2 receptor sites and can block both plasma extravasation and bronchoconstriction (Regoli et al. 1994), may also be useful for respiratory diseases such as asthma or COPD.
Footnotes
Acknowledgements
Supported by a grant no. 1999-02323 from the USDA (Ames, IA).