
Abstract
Enteroendocrine cells are a complex population of intestinal epithelial cells whose hormones play critical roles in regulating gastrointestinal and whole-animal physiology. There are many subpopulations of enteroendocrine cells based on the major hormone(s) produced by individual cells. Intracellular calcium plays a critical role in regulating hormone release. Inositol 1,4,5-trisphophate (IP3) receptors regulate calcium mobilization from endoplasmic reticulum-derived calcium stores in many endocrine and excitatory cells and are expressed in the intestine. However, the specific subtypes of enteroendocrine cells that express these receptors have not been reported. Immunohistochemical (IHC) studies revealed that enteroendocrine cells did not express detectable levels of type 2 IP3 receptors, whereas nearly all enteroendocrine cells that produced chromogranin A and/or serotonin expressed type 1 and type 3 IP3 receptors. Conversely, enteroendocrine cells that produced glucose-dependent insulinotropic polypeptide, glucagon-like peptide-1, cholecystokinin, or somatostatin did not express detectable levels of any IP3 receptors. Subsets of enteroendocrine cells that produced substance P or secretin expressed type 1 (33% or 18%, respectively) and type 3 (10% or 62%, respectively) IP3 receptors. Thus, different subtypes of enteroendocrine cells, as well as individual cells that express a particular hormone, exhibit remarkable heterogeneity in the molecular machineries that regulate hormone release in vivo. These results suggest that therapeutic agents can be developed that could potentially inhibit or promote secretion of hormones from specific subtypes of enteroendocrine cells.
E
GIP is produced almost exclusively by a specific subpopulation of EE cells in the proximal small intestine, called intestinal K-cells. This hormone is released in response to nutrients present in the lumen of the gut but not to those circulating in the blood (Fehmann et al. 1995; Schmier 1995; Drucker 1998). Surprisingly, hormone release from GIP-producing cell lines is independent of ATP-regulated potassium channels (Ramshur et al. 2002). Consistent with this result, IHC studies revealed that GIP-producing EE cells in the mouse small intestine do not express detectable levels of inward rectifying potassium channel 6.1 (Kir 6.1) or inward rectifying potassium channel 6.2 (Kir 6.2) (Wang et al. 2003). Therefore, ATP-regulated potassium channels are not major regulators of cell depolarization and subsequent calcium mobilization in GIP-producing EE cells in vivo. A detailed survey of the mouse small intestinal epithelium revealed that EE cells that produce GLP-1, CCK, or SST also do not express detectable levels of Kir 6.1 or Kir 6.2 (Wang et al. 2003). Conversely, most of the EE cells that produce chromogranin A (CGA), serotonin, and/or substance P (sub P) also express Kir 6.2, suggesting that these particular subpopulations of EE cells utilize ATP-regulated potassium channels to control hormone release. About 40% of EE cells that produced secretin also expressed Kir 6.2. Therefore, different subtypes of EE cells exhibit remarkable heterogeneity with respect to expression of subunits required to generate functional ATP-regulated potassium channels.
Endocrine cells can also release hormones independent of ATP-regulated potassium channels by mobilizing calcium from intracellular stores (Rutter 2001). Release of ER-derived calcium stores can be regulated by ryanodine receptors (RyR) (Islam 2002) and/or inositol 1,4,5-trisphosphate receptors (IP3Rs) (Blondel et al. 1995; Taylor et al. 1999; Hagar and Ehrlich 2000; Thrower et al. 2001). IP3Rs are expressed in secretory granules of neuroendocrine cells and are believed to facilitate secretion by controlling calcium release from the granules (Blondel et al. 1995). There are at least three different IP3Rs. Type 1, 2, and 3 receptors (IP3R-1, IP3R-2, and IP3R-3, respectively) are encoded by different genes and exhibit a high degree of overall sequence homology. IP3Rs assemble into homo- or heterotetrameric complexes (Nucifora et al. 1996). Because each isoform exhibits unique regulatory properties, complexes of different IP3Rs display different calcium-gating properties. IP3R-1 and −3 are expressed in isolated intestinal epithelial cells (Matovcik et al. 1996; Nakanishi et al. 1996), suggesting that they are present in EE cells. However, the specific subtypes of EE cells that express IP3Rs have not been identified. Therefore, double-label immunofluorescence studies were performed to determine the pattern of IP3R expression in EE cells in the mouse intestinal epithelium. This analysis revealed unexpected levels of complexity and heterogeneity in IP3R expression by different subtypes of EE cells.
Materials and Methods
Preparation of Intestinal Swiss Rolls
Animal protocols were approved by the Animal Studies Committee of Washington University. Entire small intestines from 10-week-old C57BL/6 mice (given free access to food and water) were removed en bloc immediately after the animals were sacrificed and then flushed with PBS, followed by freshly prepared 4% paraformaldehyde in PBS. Each intestine was then cut longitudinally along its entire length. After overnight fixation, the intestine was stored in 70% alcohol (at least overnight) and then rolled up from the duodenum to distal ileum. The resulting “Swiss roll” (Griffiths et al. 1988; Hermiston and Gordon 1995) was cut in half with a razor blade along the duodenal to ileal axis and then impregnated with agar (2% agar, 1.83% formaldehyde, 0.2% NaH2PO4, 0.325% Na2HPO4, pH 7). The Swiss rolls were dehydrated in graded alcohols, cleared in xylenes, infused with paraffin, and then embedded in paraffin blocks. Embedded tissues were then sectioned and labeled as described below. Using this technique, the entire duodenal to ileal axis of a single mouse small intestine can be examined on a single section.
Antibodies
Information concerning the primary and secondary antibodies is presented in Table 1. To generate our own antibodies against GIP, human GIP was purchased from California Peptide Research (Napa, CA), conjugated to KLH using EDAC, and then used to immunize guinea pigs. The specificity of the antisera was confirmed by double-label immunofluorescence. Briefly, a single paraffin-embedded section of mouse small intestine was labeled with commercial rabbit anti-GIP plus the guinea pig anti-GIP antiserum (Table 1). Bound primary antibodies were detected using FITC-conjugated donkey anti-rabbit IgG plus Cy3-conjugated donkey anti-guinea pig IgG secondary antibodies. Three hundred labeled cells were counted under the fluorescent microscope and all positive cells were labeled with both antibodies.
Antibody Labeling
Tissue sections were deparaffinized in xylene and rehydrated in graded alcohols followed by water. Each of the anti-IP3R antibodies required antigen retrieval techniques (Pileri et al. 1997) to unmask immunogenic epitopes. This was performed using aqueous solutions of either 1 mM ethylenediaminetetraacetic acid, pH 8 [(EDTA) to detect IP3R 1, 2, and 3] or 10 mM sodium citrate, pH 6 [(citrate) to detect pan IP3R epitopes]. Briefly, slides were added to pre-warmed EDTA or citrate solution, placed in a pressure cooker, and then incubated for 3 min at 20 psi of pressure. Once the pressure returned to normal, slides were transferred to fresh buffer that had been incubated simultaneously in the pressure cooker. After cooling to room temperature (RT), sections were washed with PBS and then blocked using BACKGROUNDSNIPER (Biocare Medical; Walnut Creek, CA). Sections were then incubated overnight at 4C with mixtures of the appropriate primary antibodies to IP3Rs and EE cell products diluted to the indicated concentration in Da Vinci Green Primary Antibody Diluent (Biocare Medical) (Table 1). After three washes with PBS, sections were incubated for 45 min at RT with the indicated secondary antibodies diluted in Da Vinci Green Primary Antibody Diluent (e.g., Cy3-conjugated donkey anti-rabbit IgG plus FITC-conjugated donkey anti-guinea pig IgG). Sections were washed, incubated with bis-benzimide to label nuclei, and then mounted in PBS/glycerol as described (Wice and Gordon 1998). In all cases, EE cells were not labeled when the primary antibodies were omitted.
Antibodies used to detect EE cell hormones and IP3Rs a
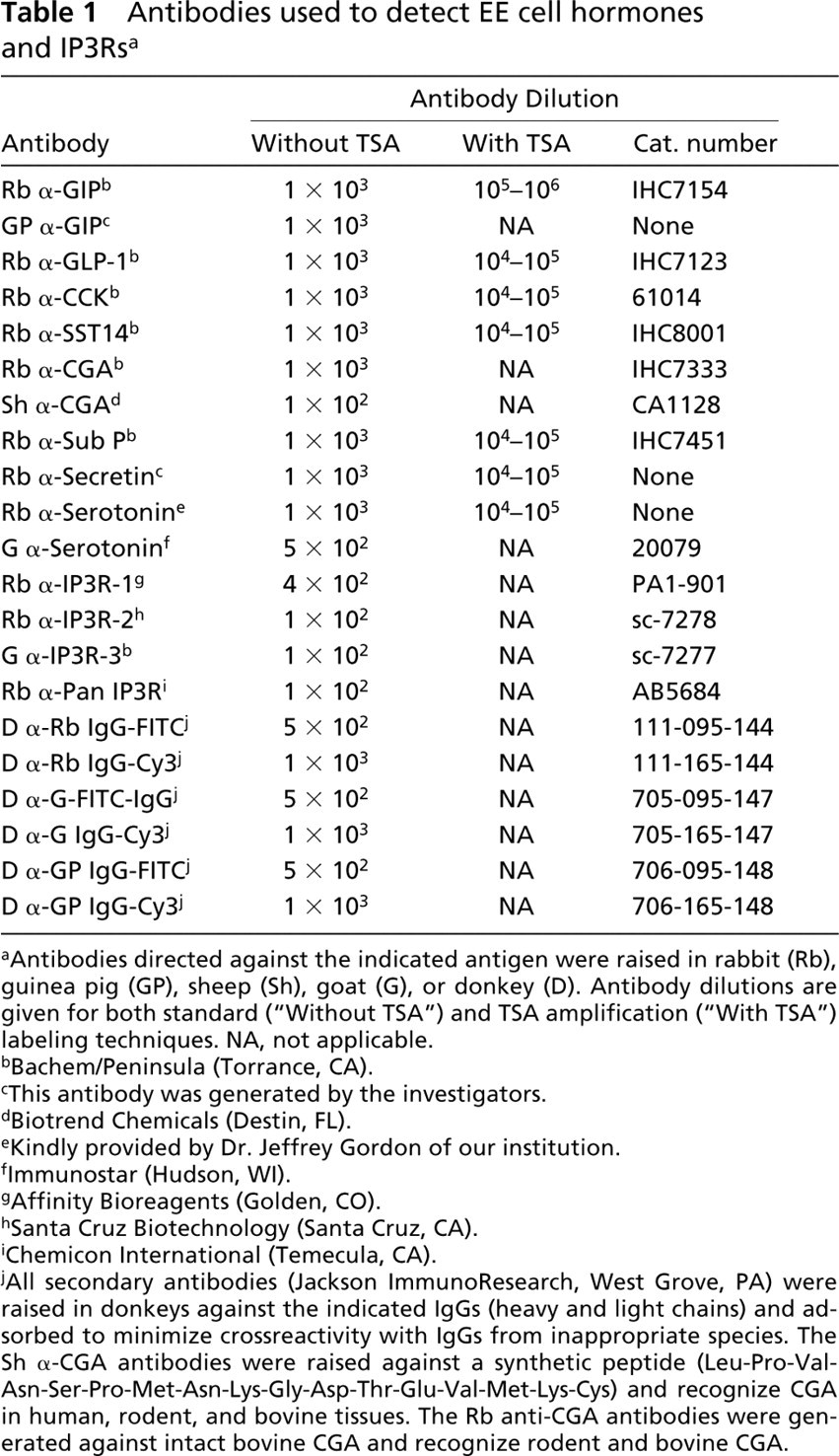
aAntibodies directed against the indicated antigen were raised in rabbit (Rb), guinea pig (GP), sheep (Sh), goat (G), or donkey (D). Antibody dilutions are given for both standard (“Without TSA”) and TSA amplification (“With TSA”) labeling techniques. NA, not applicable.
bBachem/Peninsula (Torrance, CA).
cThis antibody was generated by the investigators.
dBiotrend Chemicals (Destin, FL).
eKindly provided by Dr. Jeffrey Gordon of our institution.
fImmunostar (Hudson, WI).
gAffinity Bioreagents (Golden, CO).
hSanta Cruz Biotechnology (Santa Cruz, CA).
iChemicon International (Temecula, CA).
jAll secondary antibodies (Jackson ImmunoResearch, West Grove, PA) were raised in donkeys against the indicated IgGs (heavy and light chains) and adsorbed to minimize crossreactivity with IgGs from inappropriate species. The Sh α-CGA antibodies were raised against a synthetic peptide (Leu-Pro-Val-Asn-Ser-Pro-Met-Asn-Lys-Gly-Asp-Thr-Glu-Val-Met-Lys-Cys) and recognize CGA in human, rodent, and bovine tissues. The Rb anti-CGA antibodies were generated against intact bovine CGA and recognize rodent and bovine CGA.
When both primary antibodies were raised in the same species, a two-step procedure was conducted for double-label studies (Shindler and Roth 1996). First, tyramide signal amplification (TSA) was performed using the TSA-Plus Fluorescein System (Perkin-Elmer Life Sciences; Boston, MA) to detect the EE cell hormone. Briefly, tissues sections were re-hydrated, treated with hydrogen peroxide, subjected to antigen retrieval required for subsequent detection of the appropriate IP3R, and then incubated only with the rabbit antibody against the indicated EE cell hormone at a dilution greatly increased over the optimal dilution for the standard method (Table 1). After buffer washes, the tissue section was incubated with HRP-conjugated donkey anti-rabbit IgG antibodies (30 min at RT), washed with buffer, and then incubated with FITC-conjugated tyramide (10 min at RT). This protocol labeled the EE cell hormone with FITC (green fluorescence). Next, the tissue section was incubated with the second primary antibody, which was then detected with Cy3-conjugated donkey anti-rabbit IgG secondary antibodies (red fluorescence). Because the TSA amplifies the signal from the highly diluted first primary antibody to such a great extent, the first primary antibody is not detected by the Cy3-conjugated donkey anti-rabbit IgG antibodies used to detect the second primary antibody. This was confirmed for each primary antibody by omitting the second primary antibody. For example, the rabbit anti-GIP antibody, when used at a 1:300,000 dilution, was detected with FITC-labeled tyramide but not with Cy3-conjugated donkey anti-rabbit IgG antibodies. Because of variability in the degree of antigen retrieval and tyramide amplification on each occasion, several concentrations of the first primary antibody were used each time labeling was performed.
Quantification of Double-labeled EE Cells
EE cells expressing a particular hormone were identified with fluorescent microscope. The filters were then switched to determine whether the same EE cells also expressed the second fluorescently labeled antigen. At least 100 EE cells were identified per section and the numbers of those cells that also expressed the indicated IP3R were determined. Sections prepared from at least five different mice were counted to determine the number of GIP-producing vs CGA-producing EE cells that expressed each IP3R isoform. For statistical analyses, data for each tissue section were treated as a single sample (i.e., n = 5 for each pair of antigens). For additional double-label studies, a single section was quantified. However, qualitatively similar results were noted on at least two additional occasions.
Results
IP3R-1 Is Not Detectable in GIP-producing EE Cells
To determine if IP3Rs are expressed in GIP-producing EE cells, a single paraffin-embedded section of mouse small intestine was labeled with two antibodies, one directed against IP3R-1 and the other against GIP. Because both primary antibodies were raised in rabbits, this analysis was performed using a two-step technique (see Materials and Methods) in which the first primary antibody was used at a concentration at which it could not be detected by the secondary antibody used to detect the second primary antibody (Shindler and Roth 1996). Consistent with published results, high levels of IP3R-1 were detected in scattered cells throughout the intestinal epithelium. However, GIP was not detected in any of the IP3R-1-positive cells (Figure 1). To further confirm this observation, an anti-GIP antibody was generated in guinea pigs. Five paraffin-embedded sections of mouse small intestine, each derived from a different animal, were labeled with the guinea pig anti-GIP plus rabbit anti-IP3R-1 antibodies. More than 100 GIP-positive cells were counted on each section, and only a single cell was found that expressed both GIP and IP3R-1 (Table 2; Figure 1). This result agrees with that obtained when labeling was performed with rabbit anti-GIP followed by rabbit anti-CGA antibodies, and therefore, also demonstrates the validity of the two-step TSA technique.
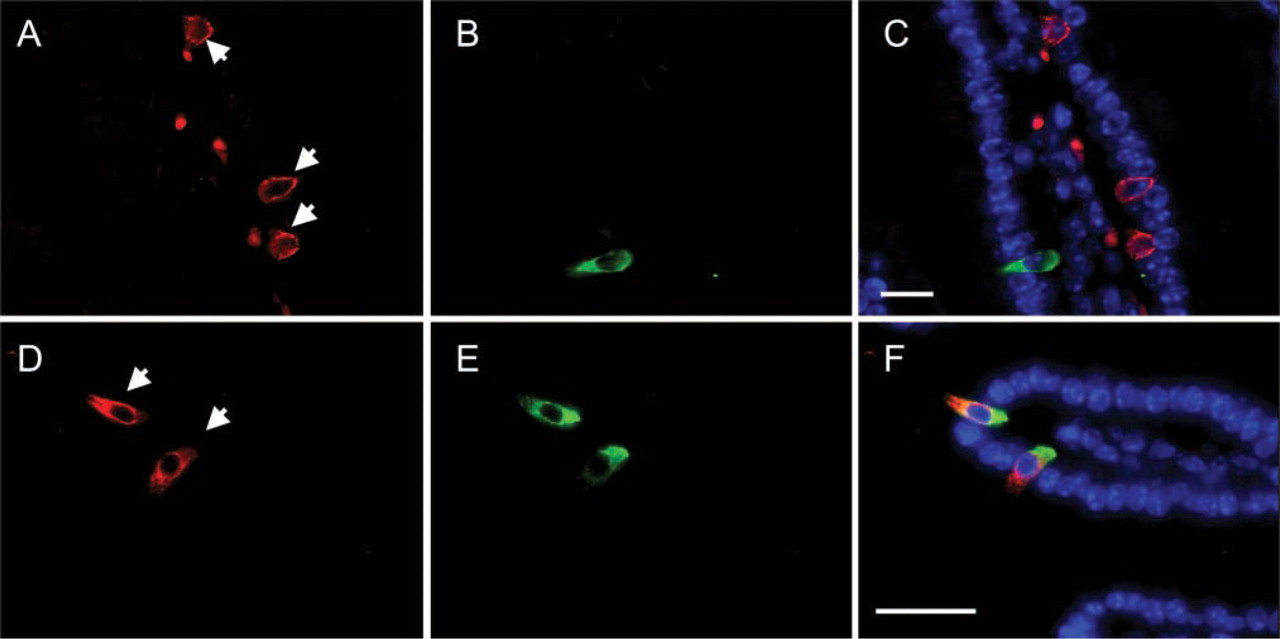
IP3R-1 Is Expressed in EE Cells that Express CGA
Results from the previous section raise the question as to which intestinal epithelial cells express IP3R-1. EE cells that produce CGA, but not GIP, express Kir 6.2 (Wang et al. 2003) suggesting that these two subpopulations of EE cells may employ fundamentally distinct secretory machineries to regulate hormone release. Therefore, sections of mouse small intestine were labeled with sheep anti-CGA plus rabbit anti-IP3R-1 antibodies. As shown in Figure 2, EE cells that express CGA also contain immunoreactive IP3R-1. Note that both IP3R-1 and CGA immunoreactivities are localized to the basal portion of these EE cells. A morphometric analysis of mouse small intestine revealed that 100% of the EE cells that produced detectable levels of CGA also expressed detectable levels of IP3R-1 (Table 2).
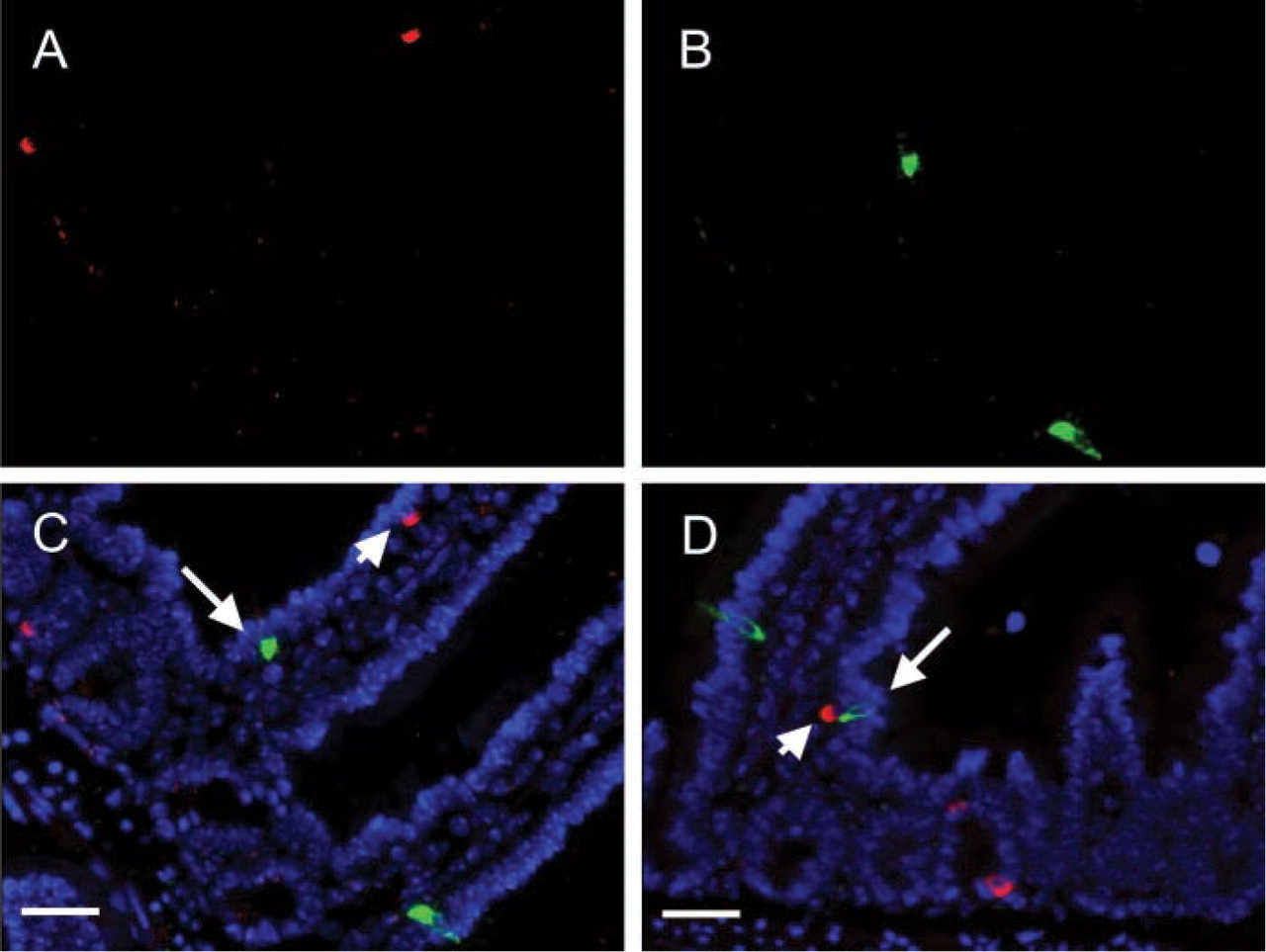
GIP-producing EE cells do not express IP3R-1. (
IP3Rs are detected in EE cells that produce CGA but not GIP a
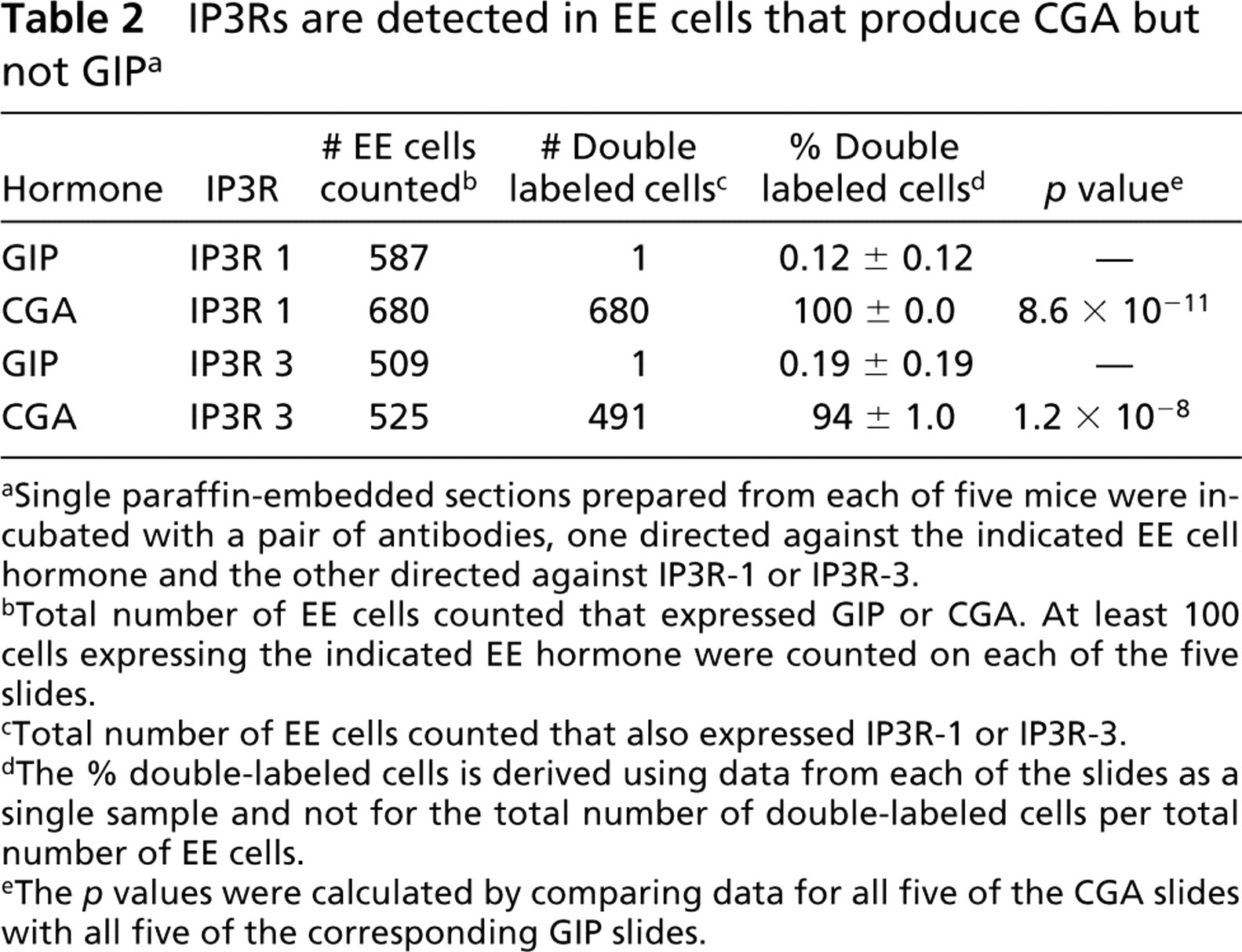
aSingle paraffin-embedded sections prepared from each of five mice were incubated with a pair of antibodies, one directed against the indicated EE cell hormone and the other directed against IP3R-1 or IP3R-3.
bTotal number of EE cells counted that expressed GIP or CGA. At least 100 cells expressing the indicated EE hormone were counted on each of the five slides.
cTotal number of EE cells counted that also expressed IP3R-1 or IP3R-3.
dThe % double-labeled cells is derived using data from each of the slides as a single sample and not for the total number of double-labeled cells per total number of EE cells.
eThe p values were calculated by comparing data for all five of the CGA slides with all five of the corresponding GIP slides.
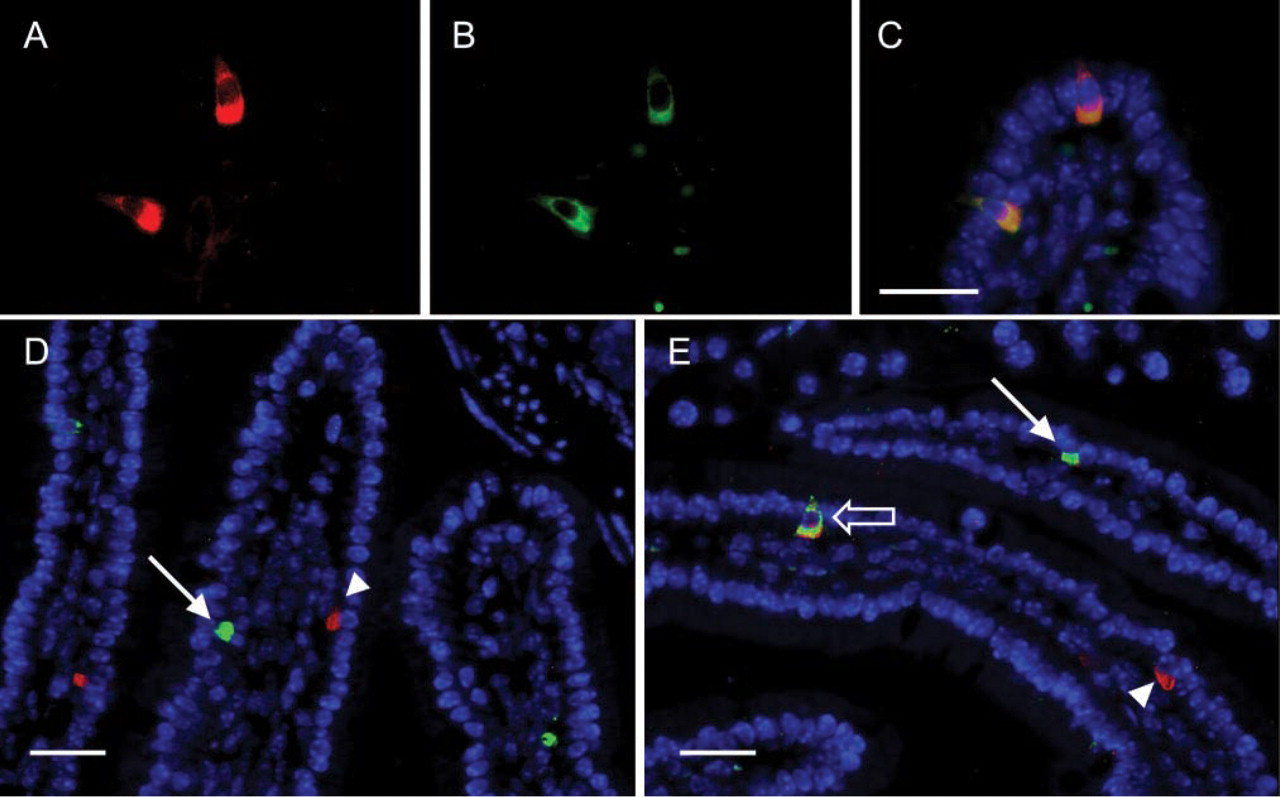
IP3R-1 is expressed in CGA-producing EE cells. (
GIP-producing EE Cells Do Not Express Detectable Levels of IP3Rs In Vivo
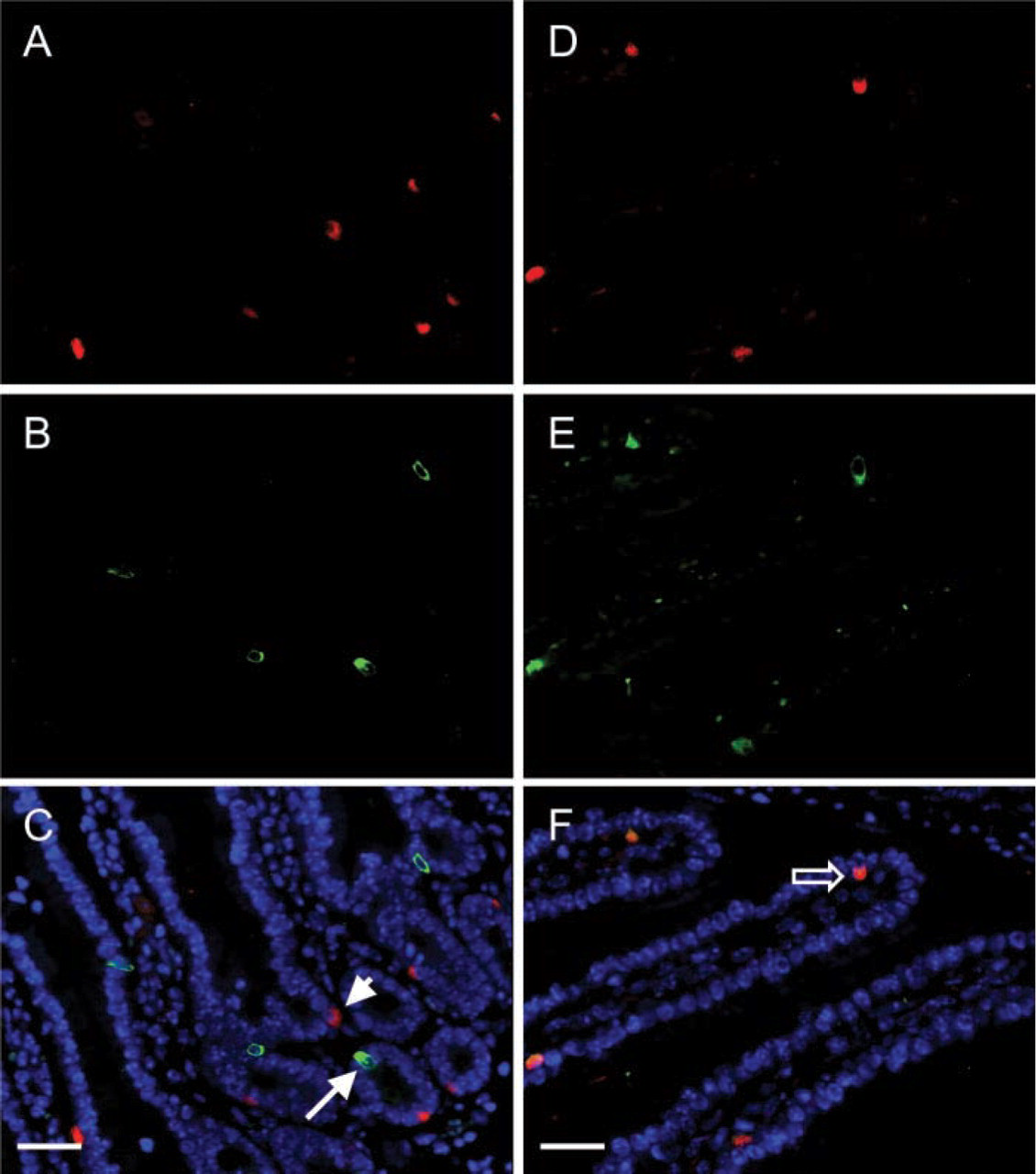
Because IP3R-1 was not detectable in GIP-producing EE cells, double-label antibody labeling was used to determine whether additional IP3Rs are expressed in this specific subtype of EE cells. IP3R-2 was not detected in any subpopulations of EE cells (not shown). Next, sections of small intestine were examined for expression of IP3R-3. Ninety-four percent of the EE cells that expressed CGA also expressed IP3R-3 (Figure 3; Table 2). In contrast to the basal localization of IP3R-1, most of the IP3R-3 immunoreactivity was present in the supranuclear region of EE cells. Therefore, almost all CGA-producing EE cells express both IP3R-1 and IP3R-3. Conversely, GIP-producing EE cells did not express detectable levels of IP3R-3 (Table 2; Figure 3). To determine whether processing of IP3Rs by GIP-producing EE cells could yield proteins that are not recognized by the isoform-specific anti-peptide antibodies, sections of small intestine were labeled with a pan-IP3R antibody that recognizes an epitope that is highly conserved in IP3R-1, −2, and −3 as well as in putative IP3Rs 4 and 5. Consistent with the fact that CGA-producing EE cells express IP3R-1 and −3, this antibody recognized antigens present in EE cells that express CGA (Figure 4). However, none of the EE cells that expressed GIP (Figure 4) contained detectable levels of this pan-IP3R epitope.
CGA Is Not Expressed in GIP-producing EE Cells
Kir 6.2 and IP3Rs are expressed in EE cells that produce CGA but not GIP. Therefore, one would predict that CGA is not expressed in EE cells that produce GIP. To directly test this hypothesis, single sections of mouse small intestine were labeled for CGA plus GIP using different combinations of antibodies. Three hundred GIP-positive cells were examined for co-expression of CGA. As shown in Figure 5, incubating a single tissue section of mouse small intestine with guinea pig anti-GIP plus rabbit anti-CGA antibodies failed to label both antigens in the same cells. Similar results were observed when a single section of mouse intestine was labeled with sheep anti-CGA plus rabbit anti-GIP antibodies. Therefore, CGA and GIP are expressed in distinct subpopulations of EE cells in the mouse small intestine.
IP3R-3 is expressed in EE cells that produce CGA but not GIP. (
Different Subtypes of EE Cells Express Unique Repertoires of IP3Rs
The results presented in Figures 1–5, coupled with published observations from our lab (Ramshur et al. 2002; Wang et al. 2003), indicate that EE cells that produce CGA, but not GIP, express Kir 6.2, IP3R-1, and IP3R-3. Therefore, studies were performed to define the pattern of IP3R expression in additional sub-populations of EE cells. Representative results are presented in Figure 2 and are summarized in Figure 6. Like GIP-producing cells, EE cells that produce GLP-1, CCK, or SST did not express detectable levels of IP3R-1. Interestingly, none of these EE cell types expressed detectable levels of Kir 6.2 (Wang et al. 2003). Ninety-eight percent of the EE cells that produced serotonin also expressed IP3R-1, whereas those that produced sub P or secretin exhibited intermediate phenotypes with 33% or 18% of each of these hormone-producing EE cells, respectively, expressing IP3R-1. Next, additional subpopulations of EE cells were surveyed for co-expression of IP3R-3. These results are summarized in Figure 6. Like GIP-producing EE cells, EE cells that produced GLP-1, CCK, or SST did not express detectable levels of IP3R-3. The pan-IP3R antibodies also failed to detect IP3Rs in these cells (not shown). Only 10% of the EE cells that produced sub P also expressed IP3R-3. Therefore, fewer than 50% of the EE cells that produce this hormone express IP3Rs, and IP3R-1 is the predominant isoform in cells that do express IP3Rs. Conversely, 62% or 87% of secretin- or serotonin-producing EE cells, respectively, produce detectable levels of IP3R-3. Therefore, most serotonin-producing EE cells express both IP3R-1 and IP3R-3, whereas IP3R-3 is the predominant isoform expressed by secretin-producing EE cells that produce IP3Rs.
Pan-IP3R antibodies do not react with antigens expressed in GIP-producing EE cells. (
Discussion
The results presented here and in a previous paper are summarized in Figure 6 and provide strong evidence that the molecular machineries that regulate hormone release from different subpopulations of EE cells are remarkably complex and distinct. EE cells that produce GIP, GLP-1, CCK, or SST do not express detectable levels of Kir 6.2 (Wang et al. 2003) or IP3Rs. Although the IHC studies do not rule out the possibility that these specific subtypes of EE cells express low levels of these proteins, it appears that Kir 6.2 and IP3Rs are much less important for regulating secretion of GIP, GLP-1, CCK, or SST vs CGA. CGA and IP3Rs are believed to be coupled in secretory granules of neuroendocrine cells (Yoo 2000). Therefore, it is interesting that EE cells that express GIP do not express detectable levels of Kirs, CGA or IP3Rs, whereas most EE cells that produce CGA and/or serotonin express high levels of Kir 6.2, IP3R-1, and IP3R-3. Other EE cells exhibit intermediate phenotypes. Most of the cells that produce sub P express Kir 6.2 but not IP3Rs whereas 43%, 18%, and 62% of the cells that produce secretin express Kir 6.2, IP3R-1, and IP3R-3, respectively.
The unique pattern of IP3R expression in different subtypes of EE cells in the mouse small intestine has important biological implications. First, IP3R-1 and IP3R-3 appear to localize to distinct subcellular compartments (basal vs supranuclear, respectively), suggesting that these two different IP3Rs may regulate calcium flux from distinct intracellular pools. Second, the open probabilities for each of the IP3Rs vary as a function of intracellular IP3 and calcium concentrations (Thrower et al. 2001). Therefore, differential expression of these channels would allow different subtypes of EE cells to secrete hormones in response to different concentrations of intracellular signaling molecules. Remarkably, individual cells that produce sub P or secretin express different combinations of Kir 6.2, IP3-1, and IP3R-3. Because these observations were noted in single tissue sections prepared from fed animals, this heterogeneity can not be explained simply by differences in the nutritional state of different mice. Therefore, a single ligand could elicit multiple secretory responses for sub P and secretin.
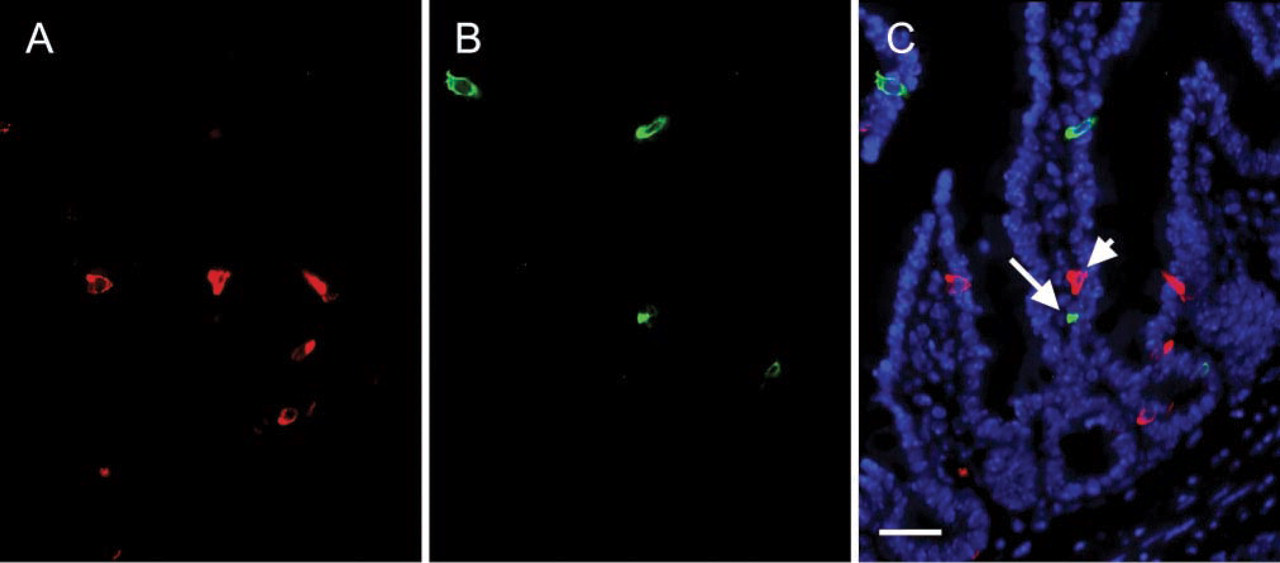
CGA is not expressed in GIP-producing EE cells. A single section of mouse small intestine was labeled with rabbit anti-CGA plus guinea pig anti-GIP antibodies as described in Figure 1.
GIP-producing EE cells reside predominantly in the proximal small intestine and are located in the crypts as well as on the villi. Intestinal L-cells, which produce GLP-1 plus additional hormones, are located principally in the distal intestine. Neither subtype of EE cells expresses detectable levels of Kir 6.2, IP3R-1, or IP3R-3. These results indicate that the patterns of expression of these regulatory molecules are not simply due to the geographic location of different subpopulations of EE cells. Rather, expression of Kir 6.2, IP3R-1, and IP3R-3 is differentially regulated along with different gut hormones.
Our results suggest that EE cells that produce GIP, GLP-1, CCK, or SST do not express CGA. It has been previously reported by many groups that SST-expressing EE cells produce little if any CGA (Varndell et al. 1985; Buffa et al. 1989; Cetin and Grube 1991; Portela-Gomes et al. 1997). However, there are conflicting reports in the literature concerning CGA expression in other subtypes of EE cells (Varndell et al. 1985; Buffa et al. 1989; Cetin et al. 1989; Cetin and Grube 1991; Portela-Gomes et al. 1997). It has been reported that CGA is not expressed in GIP-producing EE cells (Buffa et al. 1989) or is expressed in only a subset of these cells (Varndell et al. 1985; Cetin and Grube 1991; Portela-Gomes et al. 1997). Fixation artifacts could account for some of the conflicting results in the literature. However, Cetin and co-workers (1989) surveyed expression of CGA plus different EE cell hormones in the gastrointestinal tract of human, pig, cat, guinea pig, and cattle. They observed that CGA was produced in specific subtypes of EE cells in one species but not in others (e.g., CCK). Furthermore, individual subtypes of EE cells in a single species could express either none or very high levels of CGA. Because all of the tissues used for their study were similarly prepared and analyzed using the same set of antibodies, it appears unlikely that tissue fixation could account for the conflicting results. ProCGA can be processed to at least 10 different bioactive peptides that (a) inhibit hormone release from many cell types, including islet β-cells, parathyroid chief cells, and parietal cells, (b) regulate intermediary metabolism in peripheral tissues, (c) inhibit vasoconstriction, (d) trigger apoptosis, and (e) exhibit antimicrobial activity (Taupenot et al. 2003). Therefore, an alternative explanation for the conflicting results is that CGA is processed to different peptides by various EE cells and that individual antibodies to CGA can recognize the intact CGA molecule plus only specific subsets of CGA-derived peptides. As shown in Figure 5, we did not detect CGA and GIP in any of the same EE cells. This result was obtained using two different antibodies to GIP and two different antibodies to CGA. One of the antibodies to CGA was raised against the intact protein, whereas the other was generated against a synthetic peptide (see Table 1). When tissue sections were labeled with the sheep anti-CGA plus the rabbit anti-CGA antibodies, EE cells were labeled either with both antibodies or with neither antibody. Therefore, we feel that we are detecting intact CGA in EE cells that express detectable levels of this molecule. Thus, our results could be interpreted to suggest that EE cells that produce GIP, GLP-1, CCK, or SST do not produce detectable levels of intact CGA but may contain CGA-derived peptides.
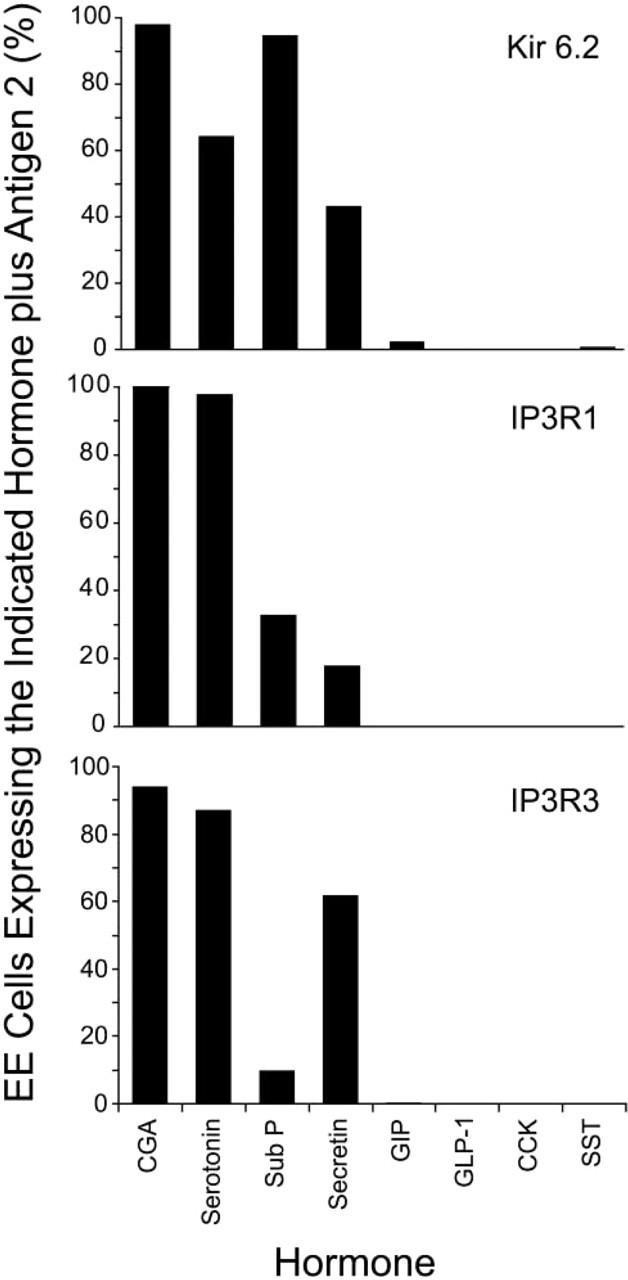
Different subtypes of EE cells express specific repertoires of proteins that regulate hormone release. Values represent the number of EE cells producing a specific hormone that also express Kir 6.2 (top), IP3R-1 (middle), or IP3R-3 (bottom). Data for GIP-producing and CGA-producing EE cells are taken from Table 2. Values for Kir expression are taken from Wang et al. (2003). All other values were calculated after analysis of at least 100 cells expressing the indicated EE cell hormone. Note that EE cells that produce CGA, sub P, serotonin, or secretin express distinct combinations of Kir 6.2 and IP3Rs. Conversely, EE cells that produce GIP, GLP-1, CCK, or SST do not express detectable levels of Kir 6.2 or IP3Rs.
Intact chromogranins are low-affinity, high-capacity calcium-binding proteins that aggregate at acidic pH. This aggregation facilitates the sequestration of hormones and other proteins that enter the regulated secretory pathway (Taupenot et al. 2003). It has been proposed that CGA is an “on/off” switch that controls the biogenesis of dense-core granules in endocrine and neuroendocrine cells (Kim et al. 2001). Because certain subtypes of EE cells do not express detectable levels of intact CGA, other chromogranins or chromogranin-like molecules must play a role in secretory granule formation in these cells. Chromogranin B was not detected in GIP-producing EE cells in humans (Portela-Gomes et al. 1997) or guinea pigs (Cetin and Grube 1991), whereas chromogranin C was detected in GIP-producing EE cells in guinea pigs (Cetin and Grube 1991), but not in humans (Portela-Gomes et al. 1997). However, it is important to keep in mind that, as has been observed with CGA, studies using antibodies to different regions of chromogranin B have yielded conflicting results (Norlen et al. 2001; Portela-Gomes and Stridsberg 2002a,b). Therefore, we can not exclude the possibility that GIP-producing EE cells express chromogranin B.
Our results raise an important question: what molecules control cell depolarization and calcium mobilization from EE cells that do not express detectable levels of Kir 6.2 or IP3Rs? RyRs, as well as IP3Rs, can regulate release of ER-derived calcium stores (Islam 2002). Nicotinic acid adenine dinucleotide phosphate is a newly identified regulator of calcium mobilization from lysosomal stores (Lee and Aarhus 1995; Lee 2003; Patel et al. 2001). This calcium store has been shown to be important for calcium mobilization by islet β-cells (Johnson and Misler 2002; Masgrau et al. 2003; Mitchell et al. 2003), exocrine pancreas (Cancela et al. 1999; Burdakov and Galione 2000), T-lymphocytes (Berg et al. 2000), and sea urchin eggs (Churchill et al. 2002). Studies are under way to determine whether RyRs or lysosomal calcium stores are involved in calcium mobilization from intracellular stores in GIP-producing EE cells.
EE cells that produce GIP, GLP-1, CCK, or SST do not express detectable levels of IP3Rs, Kir 6.2, or Kir 6.1. Therefore, do these subtypes of EE cells express the same repertoire of regulatory proteins or will a detailed molecular analysis reveal additional heterogeneity and complexity in their secretory machineries? Recent observations have demonstrated that GIP plays a critical role in promoting high fat diet-induced obesity and insulin resistance (Miyawaki et al. 2002). PYY, produced by GLP-1-producing L-cells, has been shown to inhibit food intake (Batterham et al. 2002). Therefore, compounds that inhibit GIP secretion and/or promote PYY release represent potential therapeutic drugs that could reduce obesity and insulin resistance. Because hormone release from different subtypes of EE cells appears to be regulated by distinct machineries, an understanding of the unique nature of each type of EE cell should provide important information to lay the groundwork for the development of novel drugs to treat human diseases by targeting the secretory machineries of specific subtypes of EE cells.
Footnotes
Acknowledgements
Supported in part by a Career Development Award from the American Diabetes Association (BMW), a Monsanto-Pharmacia/Washington University Biomedical Research Program Grant, and a Diabetes Research and Training Center Pilot and Feasibility Grant (NIH 5 P60 DK20579). Some services were obtained through the Digestive Diseases Research Core Center of Washington University (P30 DK52574).
We also wish to thank Dr David Kipnis for helpful discussions.