
Abstract
Gastrin-releasing peptide (GRP) and its receptor (GRP-R) are not normally expressed by epithelial cells lining the colon but are aberrantly expressed in cancer, where they act as morphogens and regulate tumor cell differentiation. Studies of colon cancer formation in mice genetically incapable of synthesizing GRP-R suggested that this receptor's morphogenic properties were mediated via focal adhesion kinase (FAK). We therefore set out to determine the presence of both total and phosphorylated forms of FAK in human colon cancer specimens as a function of tumor cell differentiation and GRP/GRP-R co-expression. Ten colon cancers containing 25 regions of distinct differentiation were randomly selected from our GI Cancer Tumor Bank. All specimens were immunohistochemically probed using antibodies recognizing GRP, GRP-R, total FAK, and FAK specifically phos-phorylated at tyrosine (Y) 397, 407, 576, 577, 861, and 925. Antibody-specific chromogen was determined by quantitative immunohistochemistry (IHC) for each region of defined differentiation. Here we confirm that GRP/GRP-R co-expression is a function of differentiation, with highest levels observed in well-differentiated tumor cells. We also show that the amount of total FAK and of FAK phosphorylated at Y397 and Y407 tightly correlates with differentiation and with the amount of GRP/GRP-R co-expression. These findings are consistent with GRP/GRP-R acting as a morphogen by activating FAK, and suggest that this occurs via phosphorylation of this enzyme at two specific tyrosine residues.
F
A number of studies have shown increased expression of FAK in colon cancers compared to that observed in adjacent non-malignant mucosal epithelial cells (Owens et al. 1995; Han et al. 1997; Cance et al. 2000). Yet these studies have been limited to evaluating total amounts of FAK present. As an enzyme, however, FAK is not functional until it is activated, a process that requires tyrosine (Y) autophosphorylation at one or more sites including Y397, Y407, Y576, Y577, Y861, and Y925 (Eide et al. 1995; Rodriguez-Fernandez 1999; Sarkar 1999). Therefore, in evaluating the presence of FAK in cancer it is important not only that enzyme quantity be considered but that activity in the form of protein phosphorylation also be determined.
Tumor cell differentiation is another important parameter that has yet to be considered in studies of FAK expression in colon cancer. In the context of solid tumors, differentiation describes the degree to which the tumor cell(s) resemble the non-malignant tissues whence they originated. Clinically, the most important implication of tumor differentiation relates to the fact that it correlates with the development of metastases for all tumors studied (Birchmeier et al. 1996; Villavicencio et al. 1997; Kowalski and Medina 1998; Nicolson and Moustafa 1998; Ng 1998; Park et al. 2000; Rampaul et al. 2001), including colorectal cancers (Jass et al. 1986). Yet colon cancer is unusual and differs from most other solid tumors—including even rectal cancers (Jass et al. 1986)—insofar as it tends to be heterogeneously differentiated (Steinberg et al. 1986; Shepherd et al. 1989; Carroll et al. 1999). Hence the presence of multiple regions of distinct differentiation, as exist in colon cancer, makes this tumor type an excellent model for studying the regulation of differentiation at the cellular level.
We have recently proposed that gastrin-releasing peptide (GRP) and its receptor (GRP-R) are morphogens, critically involved in regulating colon cancer differentiation. Although mature epithelial cells lining the colon do not normally express GRP/GRP-R, they are upregulated in post-neoplastic transformation where they act to retain tumor cells in a better-differentiated state (reviewed in Jensen et al. 2001). Our study of chemically-induced colon cancers in wild-type mice and mice genetically incapable of synthesizing GRP-R (i.e., GRPR−/- mice) suggested that FAK might be critically important for regulating differentiation. Specifically, the amount of GRP/GRP-R expressed in tumors occurring in wild-type mice tightly correlated with improved differentiation as well as the amount of total focal adhesion kinase (FAK), whereas this enzyme was nearly undetectable in tumors developing in GRPR−/- mice (Carroll et al. 2000). However, nothing is known about the phosphorylation status of this enzyme as a function of either differentiation or GRP/GRP-R expression in human colon cancer.
We therefore undertook this study to evaluate FAK expression, as well as FAK activity as measured by the presence of various phosphorylated forms of this enzyme, in human colon cancers as a function of the differentiation of individual tumor cells. Using our novel algorithm for quantitative immunohistochemistry (Matkowskyj et al. 2000,2003), we confirm that GRP/GRP-R co-expression is a function of tumor cell differentiation. In addition, we show for the first time that total FAK, as well as FAK phosphorylated at tyrosine 397 (pY397) and tyrosine 407 (pY407), is also present as a function of tumor cell differentiation and GRP/GRP-R expression in human colon cancer. In contrast, the other phosphorylated forms of FAK (pY576, pY577, pY861, pY925) were variably present in colon cancer and did not correlate with either tumor cell differentiation or the amount of GRP/GRP-R expression. These findings strongly suggest that GRP/GRP-R regulates colon cancer cell differentiation by causing FAK to be phosphorylated at specific tyrosines located at amino acid positions 397 and 407.
Materials and Methods
Materials
Anhydrous ethanol (100%, 95%) and xylene were purchased from Pharmco Products (Brookfield, CT). Wash Buffer, Target Retrieval Solution, Protein Block Serum, Antibody Diluent, LSAB2 System-HRP, Liquid DAB Substrate-Chromogen System, EnVision+ HRP (DAB)—Rabbit System, and Automated Hematoxylin were all from DAKO (Carpinteria, CA). Auto/Iodine, Redusol, and all other reagents were obtained from Fisher Scientific (Pittsburgh, PA).
As previously described (Carroll et al. 1999), GRP expression was detected using a polyclonal rabbit anti-human peptide antibody directed to amino acids 14–27 from DAKO; while GRP-R expression was detected using a polyclonal rabbit anti-peptide antibody directed to the distal third intracellular loop of the mouse and human GRP-R (CVEGNIHVKKQIESRKR). Total FAK expression was detected using a monoclonal mouse anti-human whole peptide antibody from Upstate Technologies (Lake Placid, NY), while phosphospecific FAK staining was detected using phosphorylation site-specific polyclonal rabbit antibodies from Biosource International (Camarillo, CA).
Tumor Specimens and Histological Grading
The GI Cancer Tumor Bank contains surgical pathology specimens from all patients undergoing surgical resection for colon cancer between 1980 and 1999. Ten tumors were randomly selected equally divided across all four Dukes' stages. The University of Illinois Institutional Review Board approved this study. Differentiation was assessed non-subjectively across a 5-stage scale as previously defined (Carroll et al. 2000; Glover et al. in press). Briefly, digital photographs were obtained of all regions and differentiation was assessed according to the ratio of the tumor cell nuclear area to overall cell area (N:C), an accepted paradigm for assessing tumor cell differentiation (Hamilton et al. 1989; Fernandez-Lopez et al. 1999). As previously defined, well-differentiated tumor cells were defined as those showing the presence of wellformed glands containing malignant columnar cells with small regular nuclei, and had an N:C of <0.410 (Glover et al. in press). In contrast, poorly differentiated tumors were defined as having a complete absence of gland formation with a N:C ratio >0.951. Moderately differentiated tumors had well formed glands but contained cells that were less columnar or frankly cuboidal, along with reduced cell polarity and more dysplastic nuclei, possessing a N:C ratio between 0.551 and 0.750. The intermediate grades of moderately well (N:C = 0.411–0.550) and moderately poor (N:C = 0.751–0.950) were defined in a similar fashion. Using this system, we next determined the differentiation status of every single tumor cell contained within each colon cancer. Distributed among the 10 tumor samples were six areas of well-, four areas of moderately well-, six areas of moderate, four areas of moderately poor, and five areas of poorly differentiated tumor cells. Areas of differentiation within a given tumor sample ranged from one to four distinct regions within any one sample. The smallest histologic area was poorly differentiated and measured 0.3 mm × 0.4 mm, while the largest area was well differentiated and measured 2.6 mm × 10.5 mm.
Image Capture
All photomicrographs were obtained using a SPOT RT Digital Scanning Camera from Diagnostic Instruments (Sterling Heights, MI) attached to a Nikon E600 microscope system. Images were acquired at X1000 magnification and were saved in TIFF format for further evaluation.
Quantitative Immunohistochemistry
IHC was performed on 10 tumor specimens randomly selected from the UIC GI Cancer Tumor Bank and which had been originally resected between 1985 and 1997. FAK antibody conditions were established using sections of human prostate cancer, with tumors randomly selected from the UIC Cancer Center Tumor Bank. This study was approved by the UIC Institutional Review Board (IRB). In all instances, IHC was performed using antibodies resuspended in DAKO Antibody Diluent.
A modified indirect immunoperoxidase technique was performed on 4-μm-thick paraffin-embedded sections that were hydrated in graded alcohols and rinsed in a running water bath. Slides were incubated for 5 min in a 3% hydrogen peroxide solution to quench endogenous peroxidase activity. Mercuric pigments were removed by incubating in Auto/Iodine for 1 min, followed by incubation in Redusol two times for 2 min each. Hydration was completed by rinsing in DAKO 1 × Wash Buffer three times for 3 min. Antigen retrieval was used for tissue sections submerged in 1 × DAKO Target Retrieval Solution for 30 min at 100C in a steamer. Slides were allowed to cool to RT and rinsed in 1 × DAKO Wash Buffer. Sections were blocked for 15 min with Protein Block Serum and then incubated with antibody overnight at 4C. After rinsing with Wash Buffer, slides treated with GRP (1:125), GRP-R (1:750), and total FAK antibodies were incubated with biotinylated IgG for 10 min, rinsed, and then incubated with streptavidin conjugated to horseradish peroxidase (LSAB2 kit from DAKO) for 10 min. These same sections were then rinsed and incubated with Liquid DAB Substrate-Chromogen System for 5 min to identify bound antibody. Tissue sections treated with phospho-specific FAK antibodies were incubated with labeled polymer for 30 min, rinsed, and incubated with Liquid DAB+ for 10 min to identify bound antibody (EnVision+ HRPDAB kit from DAKO). After a final rinse in 1 × Wash Buffer, all slides were counterstained with DAKO Automated Hematoxylin for 2 min, dehydrated in alcohol, and mounted with a coverslip using Permount. Control tissues were processed simultaneously as the treated slides, with the exception that primary antibody was not applied.
Chromogen abundance was quantified by determining the cumulative signal strength of the digital image file of a histologically relevant region of interest (Matkowskyj et al. 2000,2003). The amount of chromogen per pixel was determined by subtracting the mathematical energy (EM) of the control slide (i.e., not exposed to primary antibody) from that in the homologous region of the experimental slide (i.e., exposed to primary antibody). Chromogen quantity (EM) is expressed as energy units per pixel (eu/pix).
Statistical Analysis
All data reported herein are valueless and are reported as energy units per pixel (eu/pix). Statistical evaluations were performed using StatView (Abacus Concepts; Berkeley, CA). In all instances, data are expressed as the mean ± SEM.
Results
As we (Carroll et al. 1999) and others (Steinberg et al. 1986; Shepherd et al. 1989) have previously demonstrated, human colon cancers are heterogeneously differentiated (Figure 1). Overall, 25 separate regions of distinct differentiation were identified in the 10 randomly selected tumors evaluated in this study. Similar to what we have previously described (Carroll et al. 1999), GRP/GRP-R co-expression is a function of tumor cell differentiation such that significantly more protein present is in well-differentiated than in less well-differentiated cancer cells (Figure 1). Specifically, whereas non-malignant epithelial cells across the tumor margin did not express these proteins (Figure 1), well-differentiated tumor cells expressed 229.6 ± 28.0 eu/pix GRP and 253.7 ± 15.5 eu/pix GRP-R, but this declined to 26.1 ± 3.5 eu/pix and 17.4 ± 1.9 eu/pix, respectively, in poorly differentiated tumor cells.
We next established the conditions necessary for performing IHC using our antibodies to total and the various phosphorylated forms of FAK. To do this, we used sections of human prostate cancer because this tumor type ubiquitously expresses FAK (Rovin et al. 2002). We best demonstrated expression of total FAK and all the other phospho-FAKs in human prostate cancer using the following antibody concentrations: 1:50 for total FAK; 1.2 μg/ml for FAK-pY397, pY407, pY576, Y577, and pY861; and 2.0 μg/ml for FAK-pY925 (data shown for Y861 in Figure 2A).
Using these concentrations, we evaluated human colon cancer sections that had been sequentially sectioned adjacent to those used to quantify GRP and GRP-R expression. In this way, FAK expression could be determined as a function both of tumor cell differentiation and as a function of the amount of GRP/GRP-R expressed in each region of defined histology. Similar to what we previously observed in murine colon cancer (Carroll et al. 2000), the amount of GRP, GRPR, and total FAK (t-FAK) present in human colon cancer varied as a function of tumor cell differentiation (Figure 3). Similarly, a link between tumor cell differentiation and the presence of phosphorylated forms of FAK was observed for those phosphorylated at Y397 (Figure 3 and 4) and at Y407 (Figure 3).
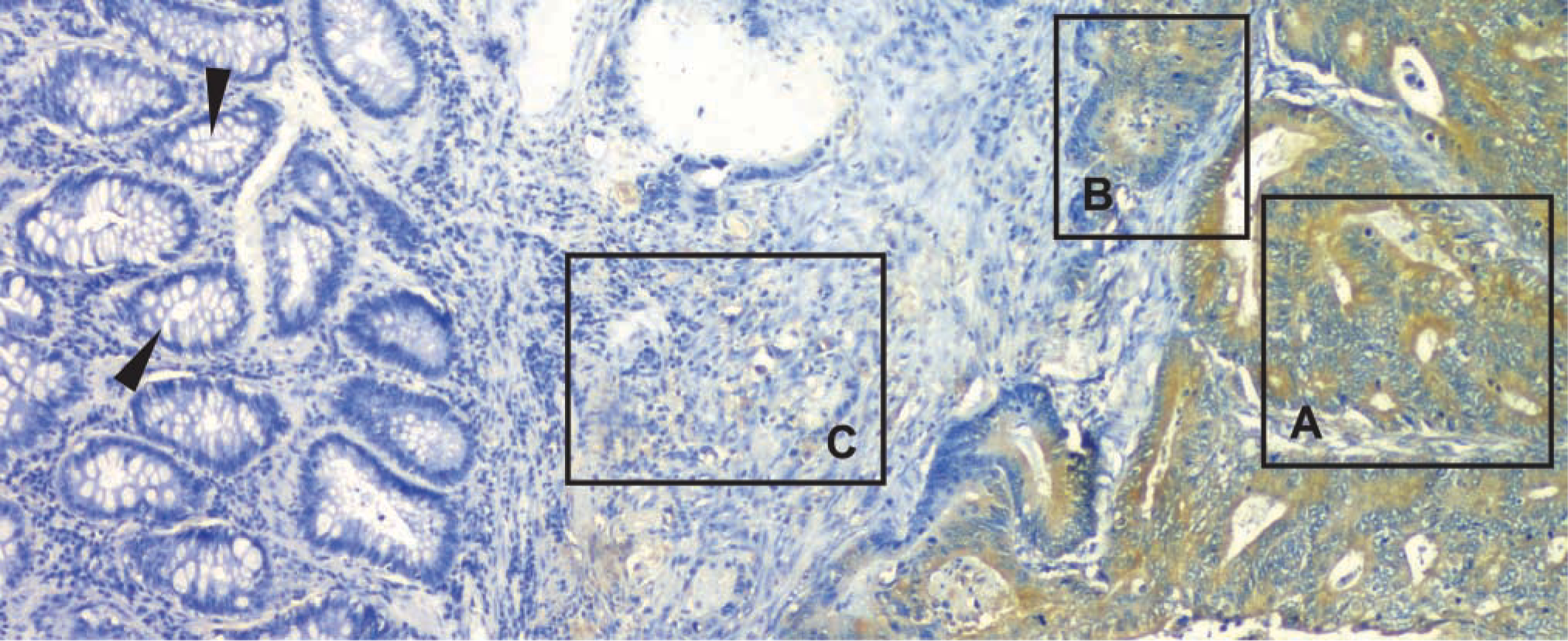
Expression of GRP-R across the margin of a human colon cancer. Expression of both GRP (not shown) and its receptor was observed only in malignant cells and was never observed in adjacent non-malignant cells (arrowheads), with chromogen quantity higher in well-differentiated tumor cells than in more poorly differentiated tumor cells [well differentiated (
The amount of total FAK and of pY397 and pY407 was not only associated with tumor cell differentiation but was also tightly correlated with the amount of GRP/GRP-R co-expression (Figure 3 and 5). To determine this, we plotted the amount of total GRP and GRP-R chromogen present in each region of defined histological differentiation with the amount of total FAK (Figure 5, left) or the amount of pY397 and pY407 (Figure 5, middle). The tightness of this correlation was striking, with correlation co-efficient's possessing r 2 values in excess of 0.9. In contradistinction, there was no correlation between GRP/GRP-R coexpression and the amount of Y576, Y577, Y861, and Y925 present (Figure 5).
The amount of chromogen generated by antibodies to the phosphorylated forms of FAK other than Y397 and Y407 varied considerably and showed no correlations with the degree of tumor cell differentiation (Table 1; Figure 5, right). Specifically, the amount of pY576-specific chromogen was less for moderately poorly differentiated cells (41.8 ± 17.0 eu/pix) than for those that were more or less well differentiated (82.6 ± 6.6 eu/pix and 80.1 ± 15.8 eu/pix for tumor cells that were moderate and poorly differentiated, respectively). A similar roller-coaster pattern was observed for pY577, with lower chromogen quantities for moderately well-differentiated tumor cells (35.6 ± 2.8 eu/pix) than for those that were well (73.2 ± 31.9 eu/pix) or moderately (87.6 ± 35.4 eu/pix) differentiated. In contrast, the least amount of chromogen for pY861 was for well (27.7 ± 7.9 eu/pix) and poorly differentiated (44.5 ± 5.0 eu/pix) tumor cells, whereas intermediate stages contained up to threefold more chromogen. But for Y925, similar low amounts of chromogen (range 34–50 eu/pix) were observed for all tumor cells, irrespective of their stage of differentiation. Finally, the very large standard errors generally observed for chromogen generated using antibodies to Y576, Y577, Y861, and Y925 (Table 1) further reflects their lack of association with differentiation.
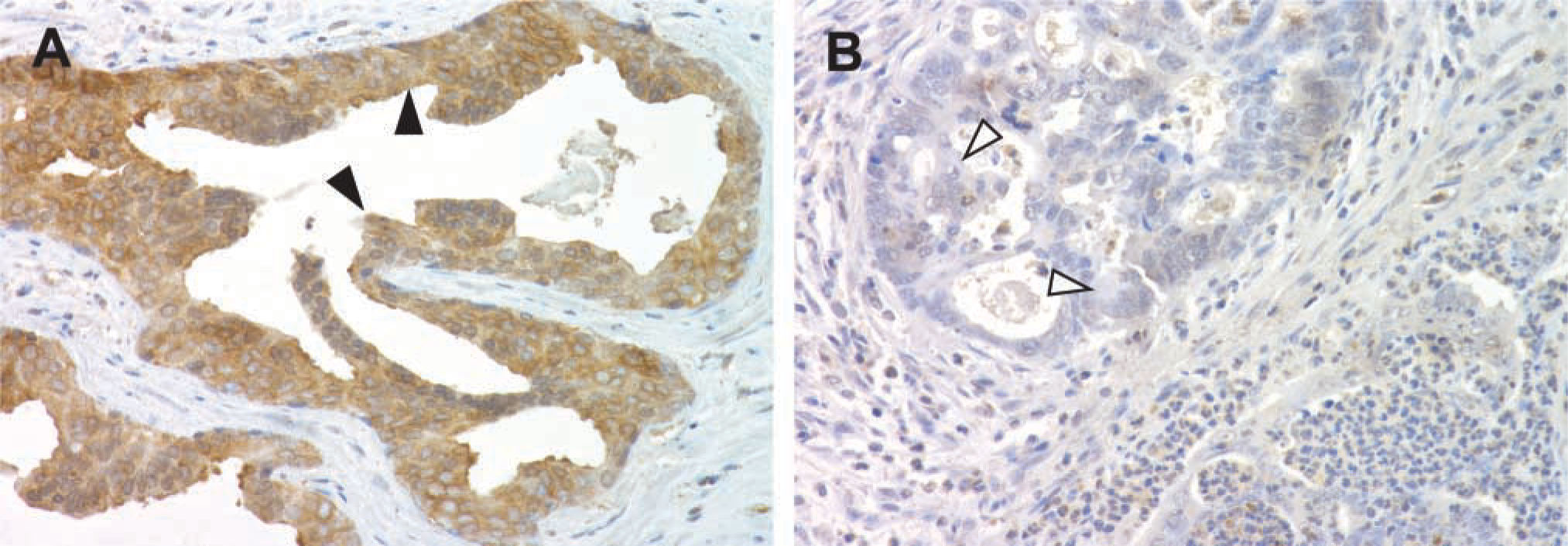
Approach used to determine conditions for FAK immunohistochemistry. (
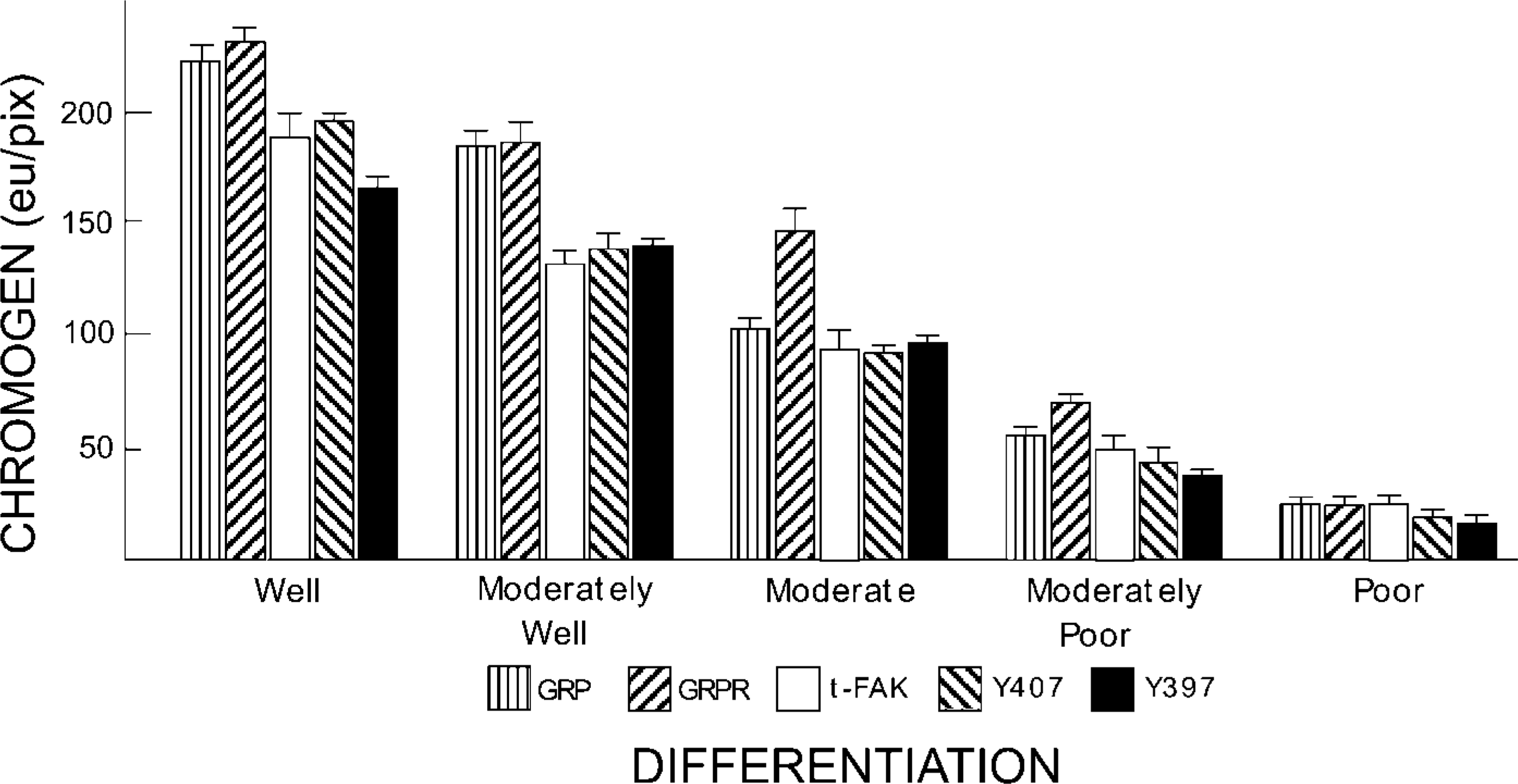
Amount of GRP, GRPR, total and indicated phosphorylated forms of FAK present in human colon cancers as a function of tumor cell differentiation. Sections were treated with the indicated antibody and the amount of chromogen quantified by Q-IHC as a function of tumor cell differentiation described in Materials and Methods. Data are shown as means ± SEM. For each antibody, chromogen quantity is statistically different compared to normal colonocyte expression and between each region of defined differentiation (p<0.05, ANOVA).
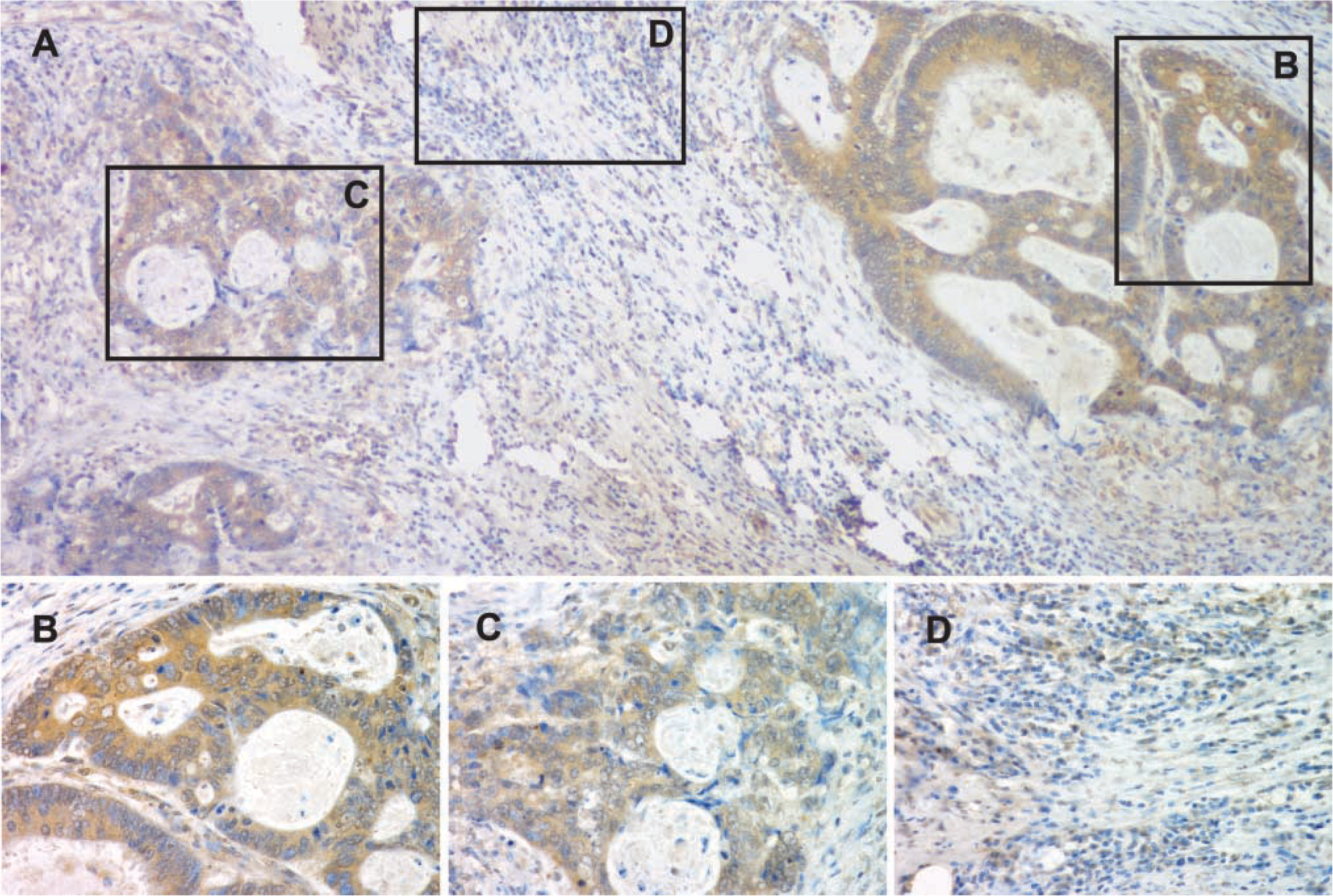
pY397 immunostaining in human colon cancer as a function of differentiation. (
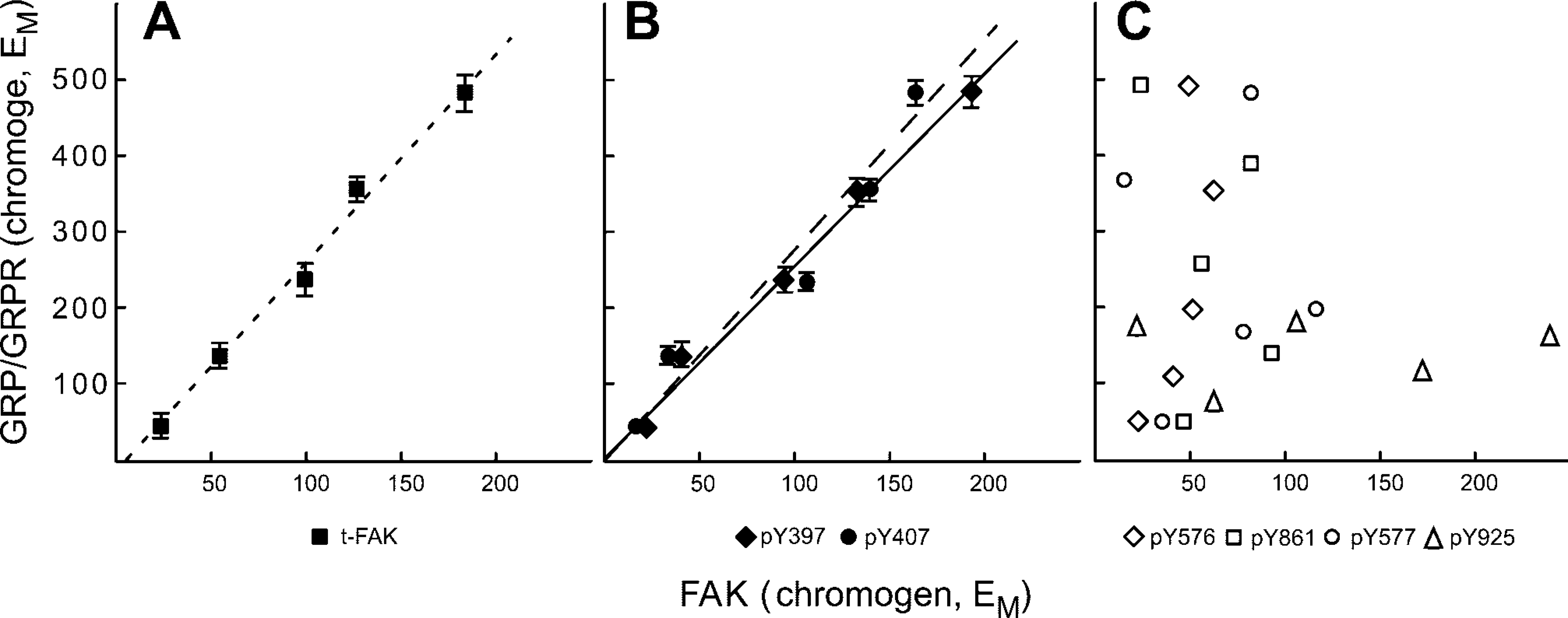
Correlation of total and phospho-FAK expression with the degree of GRP/GRP-R co-expression. (
Discussion
The role of FAK in colon cancer is not clear. Previous IHC studies performed on resected human colon cancers indicated that total FAK is present in greater amounts in invasive compared to non-invasive cancers (Owens et al. 1995). Yet this terminology is difficult to interpret because it indicates only that tumor cells have broached the muscularis propria, and as such includes all cancers irrespective of their Dukes' stage. This may account for the confusing situation where some studies have reported the presence of FAK in metastases (Han et al. 1997), whereas others failed to find evidence of this enzyme in sites away from the primary lesion (Ayaki et al. 2001). Because differentiation has been shown to be an important predictor of patient prognosis for most solid tumors studied (reviewed in Birchmeier et al. 1996), including those arising in the rectum (Jass et al. 1986; Park et al. 2000). No information has ever been presented regarding FAK and the differentiation of any tumor type.
Although the assessment of differentiation of entire colon cancers is not good for predicting patient outcome, this reflects the fact that this type of malignancy is unusual insofar as they are heterogeneously differentiated. However, when the differentiation of specific cell populations within a colon cancer is considered, this parameter does become an important prognostic parameter (Wood et al. 1981; Hase et al. 1993). For example, individual tumors containing well-differentiated cells at the leading edge (Wood et al. 1981), or at the point of budding (Hase et al. 1993), do better even in the face of local lymph node invasion than tumors not associated with local metastases but which are composed of less well-differentiated cells. Thus, differentiation and the factors regulating tumor cell appearance are as important in colon cancer as for other solid tumors, albeit with the caveat that this analysis be performed at the level of the individual tumor cell. Our novel algorithm for quantitative IHC (Matkowskyj et al. 2000,2003) now permits this type of analysis to be readily performed. Using this technique we show for the first time that total FAK, and FAK phosphorylated at Y397 and Y405 but not at other sites, correlates with the degree of individual tumor cell differentiation within any particular colon cancer.
Expression of phosphorylated FAKs in human colon cancers of various degrees of differentiationa
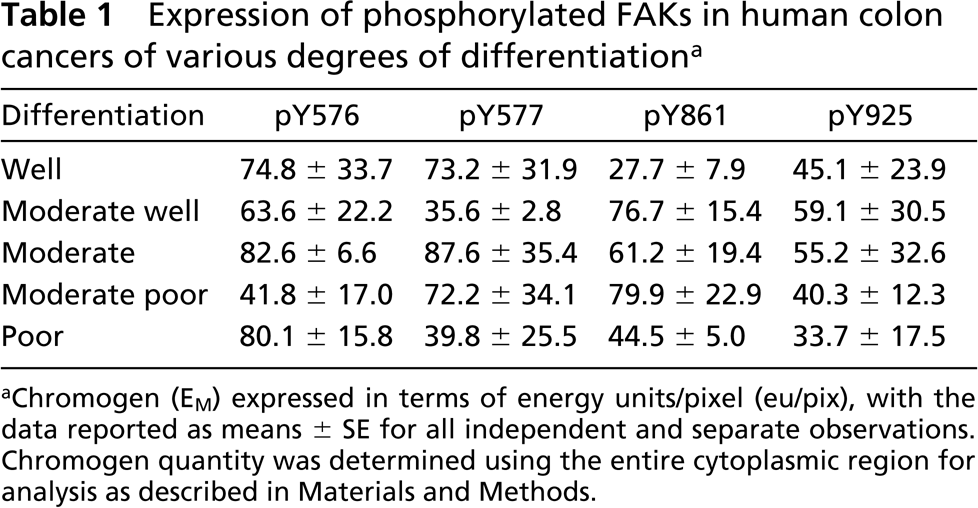
Chromogen (EM) expressed in terms of energy units/pixel (eu/pix), with the data reported as means ± SE for all independent and separate observations. Chromogen quantity was determined using the entire cytoplasmic region for analysis as described in Materials and Methods.
We previously showed that GRP/GRP-R upregulation in colon cancer recapitulates, albeit dysfunctionally, their normal role in regulating intestinal development during organogenesis (Carroll et al. 2002). Hence when aberrantly expressed in colon cancer, GRP/GRP-R act as morphogens, serving to promote the retention or adoption of a better differentiated phenotype (reviewed in Jensen et al. 2001). Indeed, GRPR−/- mice exposed to the carcinogen azoxymethane develop poorly differentiated adenocarcinomas of the colon, whereas tumors expressing GRP/GRP-R in wild type mice are well to moderately well differentiated (Carroll et al. 2000). In that study, and similar to what we show here, total FAK varied in cancers forming in wild-type mice as a function of differentiation, whereas this enzyme was simply not detected in any cancers forming in GRPR−/- mice (Carroll et al. 2000).
But FAK has traditionally been viewed, perhaps because of its ability to promote the motility of cancer cell lines studied in vitro (Sieg et al. 2000; Hauck et al. 2002a,b), as a contributor to tumor invasion and metastasis, and hence to greater malignancy. Yet the contradictory information regarding FAK expression in colon cancer metastases notwithstanding (Han et al. 1997; Ayaki et al. 2001), it could also be argued that motility is critical to allowing any tissue, including cancers, to remodel, and as such could equally be a core property of a morphogen. Intriguingly, three studies of human colon cancer cell lines (Caco-2, HT-29, and Colo 201) all show increased amounts of total FAK expression as they are subjected to conditions promoting better differentiation (Levy et al. 1998; Nakagawa et al. 1998; Rognoni et al. 1998).
Studies of murine fibroblast cell lines have shown that GRP binding to its receptor causes FAK to be phosphorylated (Sinnett-Smith et al. 1993; Zachary et al. 1993; Rozengurt 1998). However, the specific tyrosine residues phosphorylated, and whether this occurs in human colon cancer, has not been previously elucidated. This study certainly does not prove that GRP/GRP-R activate FAK by causing this enzyme to be phosphorylated at Y397 and Y405. But this study does provide the first evidence that tumor cell differentiation in colon cancer, known to be regulated at least in part by GRP/GRP-R (Carroll et al. 2000; Jensen et al. 2001), may occur via phosphorylation of specific tyrosines in FAK. And the link of FAK activation with differentiation suggests that the role of this enzyme in at least colon cancer may be less malignant than previously considered.
Footnotes
Acknowledgements
Supported by NIH grant DK-07788 (to S. Glover) and CA-094346 and a VA Merit Review award (to R.V. Benya).