
Abstract
Antigen retrieval (AR) is frequently required for successful immunohistochemistry (IHC) in archival formalin-fixed, paraffin-embedded tissue sections. Although AR by heating is most generally used, the majority of existing methods are useful only for paraffin-embedded sections. This article describes a simple alternative method for AR that can be used for aldehyde-fixed frozen sections. After fixation in paraformaldehyde, tissue blocks were heated in retrieval solutions and then frozen with dry ice. The optimal temperatures for heating were 90C and above, and the optimal retrieval solutions were distilled water and 10 mM sodium citrate, pH 6.0. Sections were cut with a cryostat and mounted on poly-L-lysine-coated glass slides. After the sections dried, routine IHC was performed. Alternatively, free-floating sections were used. This method not only greatly enhanced the immunoreactivity for a wide range of antigens, especially for nuclear proteins, but also effectively lowered the background staining in some cases. I examined the staining of 14 antibodies using sections of mouse brain and rat testis. The heating process was essential for five antibodies, improved immunoreactivity for seven antibodies, and provided no change for two antibodies.
I
Although AR by heating is widely effective, the majority of the existing methods are designed for paraffinembedded sections. Frozen sections have been generally exempt from these methods because such sections are fragile and are easily destroyed by heating. The author reports here an alternative AR method by heating, which is useful for paraformaldehyde-fixed frozen sections. The optimal conditions, including the retrieval solution, temperature, and time, were examined. This method is quite simple but effective and needs no special equipment except for a cryostat. We have already reported the successful IHC detection of several proteins using this method (Ino and Chiba 1996,2000,2001).
Materials and Methods
Preparation of Tissue Sections
Male and female ddY mice (body weight 30–40 g) and male Sprague-Dawley rats (body weight 250–300 g) were perfused via the heart with 4% paraformaldehyde in 0.1 M sodium phosphate buffer, pH 7.5 (80 ml, 5 ml/min for mice; 400 ml, 25 ml/min for rats). Mouse brains were coronally or parasagittally sliced at 3–5-mm thickness. Rat testes were scissored in the tunica albuginea. The tissues were further fixed in the same solution at 4C overnight. The testes were cut in half. The tissues were then transferred to retrieval solutions and incubated at 4C overnight. The tissue blocks were placed in a small metal basket and immersed in 300 ml of preheated retrieval solutions with gentle stirring using a hot plate for 3 min or 10 min. The heating temperatures tested were 50C, 60C, 70C, 80C, 90C, or boiling (97–99C). This heating step was omitted for the control tissue blocks. Then the blocks were immediately placed in cold 30% sucrose in PBS and incubated at 4C overnight. The heattreated and untreated (control) blocks were immersed in an embedding medium (Tissue-Tek OCT Compound; Sakura Fine Technical, Tokyo, Japan), quickly frozen with crushed dry ice, sectioned with a cryostat at 10-μm thickness, and mounted on glass slides coated with freshly prepared 0.02% poly-L-lysine. The sections were dried well for more than 1 day in a vacuum desiccator at room temperature (RT). The frozen tissue blocks could be stored at −80C for several months and the sections could be stored in a desiccated chamber at −20C for several months. To prepare free-floating sections, the frozen tissue blocks were cut with a cryostat at 40-μm thickness and the sections were stored in PBS containing 0.2% sodium azide at 4C.
All animals were treated and cared for in accordance with the Chiba University School of Medicine guidelines pertaining to the treatment of experimental animals.
Immunohistochemistry
Sections were reacted on glass slides (slide-mounted sections) or in wells (free-floating sections). The sections were immersed in 0.3% Triton X-100 in PBS for 3 hr at RT and then in a blocking solution (5% skim milk in PBS) at 4C for 4–6 hr. After brief washing with PBS, the sections were reacted with primary antibodies diluted with the blocking solution at RT overnight. The primary antibodies were as follows: rabbit polyclonals, anti-cdk5 (C-8; Santa Cruz Biotechnology, Santa Cruz, CA; 1:2000), anti-c-fos (Ab-5; Oncogene Research Products, Cambridge, MA; 1:5000), anti-Egr-1 (C-19; Santa Cruz; 1:2000), and anti-cdk2 (M2; Santa Cruz; 1:2000); goat polyclonals, anti-RORα1 (C-16; Santa Cruz; 1:2000), anti-Fos B (102; Santa Cruz; 1:2000); and mouse monoclonals, anti-PCNA (19F4; Boehringer Mannheim, Mannheim, Germany; 1:2000), anti-PCNA (PC10; Pharmingen, San Diego, CA; 1:5000), anti-cdc2 p34 (17; Santa Cruz; 1:2000), anti-tyrosine hydroxylase (MAB318; Chemicon International, Temecula, CA; 1:2000), anti-neuronal nuclei (NeuN; Chemicon; 1:2000), anti-GFAP (G5A; Boehringer Mannheim; 1:200), and anti-vimentin (V9; Boehringer Mannheim; 1:200). After washes with PBS (8 min, four times), the sections were reacted with a secondary antibody, biotinylated anti-rabbit, goat or mouse IgG(H+L) antibody (Vector Laboratories, Burlingame, CA; 1:200) at RT for 3 hr. After washes with PBS, the sections were reacted with the reagents of the Vectastain ABC Kit (Vector) at RT for 1 hr. After subsequent washes, the sections were stained with 0.3 mg/ml 3,3'-diaminobenzidine, 0.6 mg/ml ammonium nickel (II) sulfate hexahydrate, and 0.0009% hydrogen peroxide in 50 mM Tris-HCl, pH 7.5.
Brief Protocol for Antigen Retrieval
For convenience, the brief protocol for AR is listed here. Optimal conditions for immunostaining should be tested in each case.
Prepare tissues fixed with 4% paraformaldehyde in 0.1 M sodium phosphate buffer, pH 7.5. The tissue blocks should be cut to a proper size (e.g., slices 3–5 mm thick).
Immerse the tissue blocks in a retrieval solution (distilled water or 10 mM sodium citrate buffer, pH 6.0) at 4C overnight.
Place the tissue blocks in a small, heat-resistant basket and immerse in boiling retrieval solution (200–500 ml) with gentle stirring using a hot plate for 3–5 min. For heating, a conventional burner can also be used.
Immediately place the tissue blocks in cold 30% sucrose in PBS and incubate at 4C until the blocks sink.
Immerse the tissue blocks in an embedding medium and freeze quickly with crushed dry ice. The frozen tissue blocks can now be stored at −80C.
Cut frozen sections with a cryostat and mount them on glass slides coated with freshly prepared 0.02% poly-
Dry the sections well for more than 1 day in a vacuum desiccator at RT. If is no vacuum desiccator is available, dry for several days at RT to prevent detachment of the sections during processing. The dried sections can be stored in a desiccated chamber at −20C for several months.
Results
To demonstrate the effect of AR by heating, first the author compared immunostaining on slide-mounted sections, those that were boiled for 3 min in distilled water and others that were not (untreated). Then the conditions for AR were examined, including the retrieval solution, temperature, and heating time. Finally, the effect of AR on immunostaining by the freefloating method is shown. In the following experiments, sections were stained using the avidin-biotin system with 3,3'-diaminobenzidine, but the same retrieval effect was obtained with fluorophore-conjugated secondary antibodies (data not shown). In the absence of primary antibodies, no staining was observed in heat-treated tissues (data not shown).
CDK5 and RORα in the Mouse Brain
The remarkable effect of the AR method on the immunostaining of two neuronal proteins, CDK5 and RORα, in the mouse cerebellum is shown. CDK5 is one of the cyclin-dependent kinases enriched in the nervous system. CDK5 is located in both the nuclei and cytoplasm of neurons (Ino and Chiba 1996). However, only cytoplasmic staining was observed in untreated sections (Figure 1A). Heating unmasked nuclear staining and even enhanced cytoplasmic staining (Figure 1B). In Figure 1B, Purkinje cells show clear cytoplasmic and nuclear staining of CDK5. Molecular layer cells also show nuclear staining. Unmasking of nuclear CDK5 was observed in other areas as well, such as the hippocampus, cerebral cortex, and brainstem (data not shown). RORα is an orphan nuclear receptor strongly expressed in the cerebellum and thalamus (Matsui et al. 1995), and the protein is also located in these areas (unpublished observations). Although staining of RORα was occasionally observed in untreated sections, the signal was weak and unstable (Figure 1C). With heating, clear nuclear staining of RORα in Purkinje cells and molecular layer cells was demonstrated (Figure 1D).
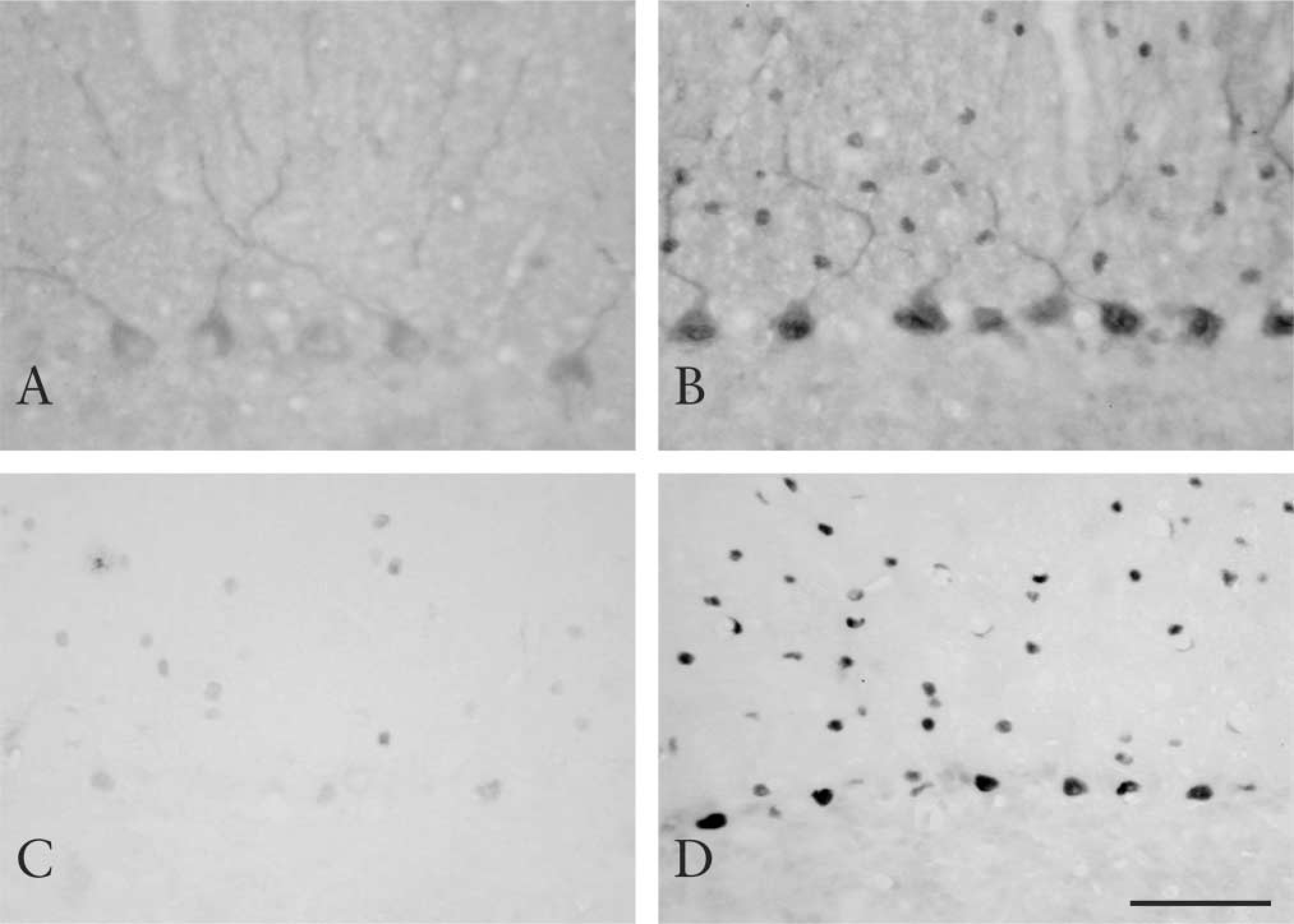
Immunostaining of CDK5 (
NeuN, GFAP, and Tyrosine Hydroxylase (TH) in the Mouse Brain
NeuN and GFAP are frequently used as cell markers for neurons and astrocytes, respectively. NeuN is a neuron-specific transcription factor located in both the nuclei and cytoplasm, and GFAP is an intermediate filament protein located in the cytoplasm. TH, a key enzyme mediating catecholamine metabolism, is located in the cytoplasm of catecholaminergic neurons. Although NeuN, GFAP, and TH were observed in untreated sections (Figures 2A, 2C, and 2E), heating enhanced their immunoreactivity (Figures 2B, 2D, and F). The AR effect on GFAP and TH immunoreactivity was conspicuous. More clearly stained GFAP-positive astrocytes and TH-positive dopaminergic neurons were seen in heat-treated sections than in untreated sections. In addition, heating lowered the background staining in these three cases.
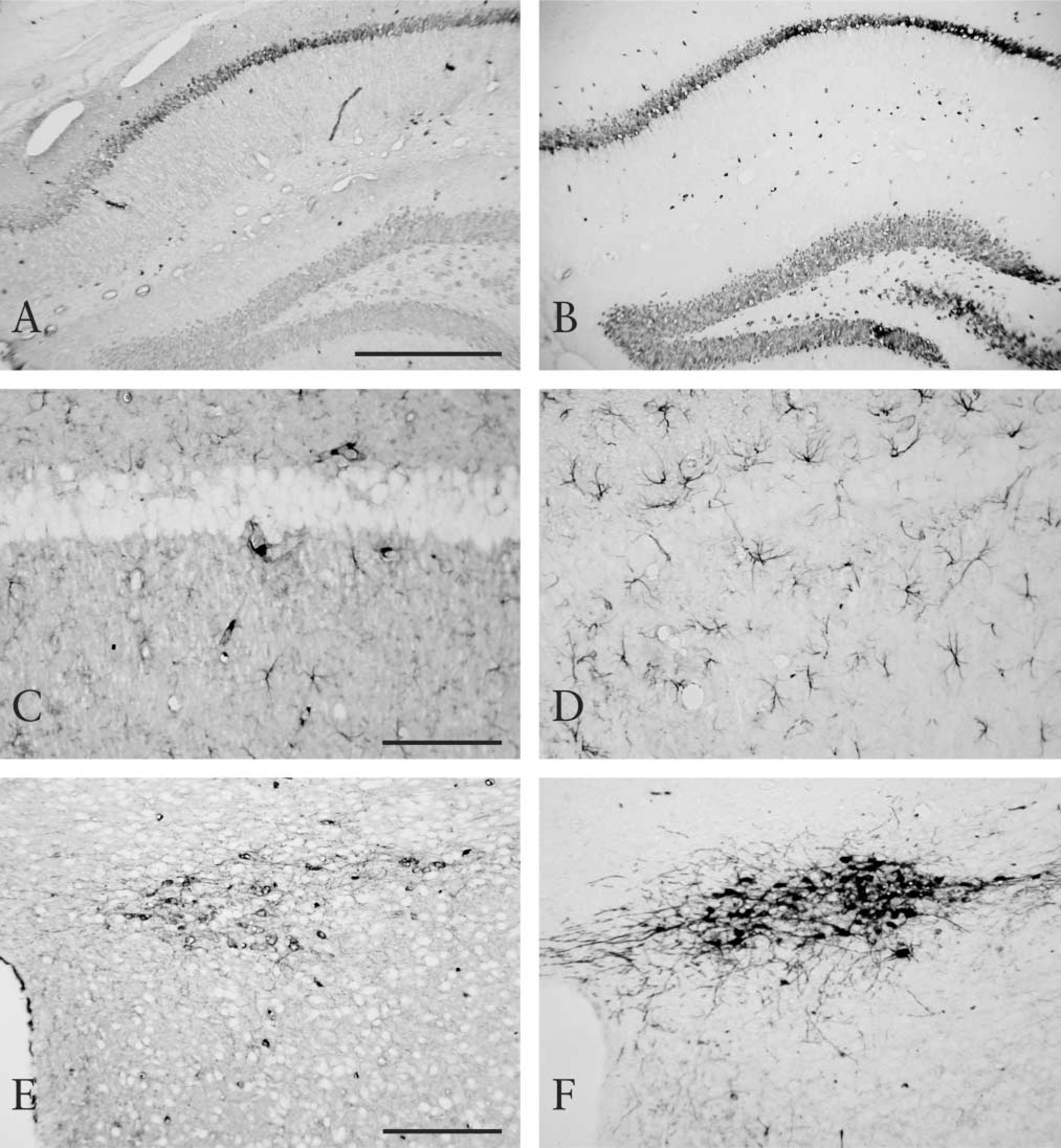
Immunostaining of NeuN (
c-Fos, FosB, and Egr1 in the Mouse Brain
c-Fos, FosB, and Egr1 are transcription factors called immediate-early genes because of their immediate induction after several stimuli. c-Fos is widely used as a marker of neuronal activity. Although these proteins are greatly induced by stimuli, certain levels of these proteins exist in the normal mouse brain. All were detected in untreated sections. Although c-Fos and FosB immunoreactivities were slightly enhanced by heat treatment, the effect was not dramatic (data not shown). Staining of Egr1 was almost uninfluenced by heating (data not shown). Staining of these proteins was seen in the nuclei.
PCNA, CDC2, CDK2, CDK4, and Vimentin in the Rat Testis
The effect of AR on immunostaining of these proteins in the rat testis was tested. PCNA is a protein necessary for DNA synthesis. It has been reported that, for detection of PCNA, AR by heating is effective in paraffin-embedded sections (Greenwell et al. 1991; Malmstrom et al. 1992). Here, two monoclonal antibodies to PCNA, PC10 and 19F4, were used. In untreated sections PCNA was not detected at all, with either PC10 or 19F4 (Figure 3A and 3C). However, clear and intense nuclear staining of PCNA was demonstrated mainly in spermatogonia with heating (Figure 3B and 3D). Although PC10 is usually preferred for detection of PCNA, this antibody also reacts with the nuclei of postmitotic nonproliferating cells, such as neurons (Ino and Chiba 2000). In contrast, 19F4 reacts only with the nuclei of proliferating cells. In the present study, PC10-positive cells were more widely distributed than 19F4-positive cells in germ cells. CDC2, CDK2, and CDK4 are cyclin-dependent kinases responsible for cell cycle progression in eukaryotic cells. Although CDC2 and CDK2 immunostaining was observed in untreated sections, the intensity was weak (Figure 3E and 3G). Heat treatment enhanced their immunoreactivity (Figure 3F and 3H), especially the nuclear staining of CDC2 (Figure 3F, arrows). In contrast, CDK4 immunoreactivity was almost unchanged (Figure 3I). CDC2 and CDK2 immunoreactivities were located mainly in the nuclei of spermatogonia and spermatocytes and were also observed in the cytoplasm of these cells. CDK4 immunoreactivity was mainly observed in the nuclei of Leydig cells and spermatogonia. Vimentin constitutes cytoskeletal intermediate filaments, which are located in Sertoli cells of the rat testis. A typical anti-vimentin antibody, V9, showed immunoreactivity in the cytoplasm of Sertoli cells in both untreated and heattreated sections (Figure 3J and 3K). Heating slightly enhanced V9 immunoreactivity.
Examination of Retrieval Conditions
Antigen retrieval by boiling tissue blocks in distilled water for 3 min was effective for a number of antibodies. The author further examined the heating conditions, including the retrieval solution, temperature, and heating time.
First, the composition and pH of the retrieval solutions were assessed under the fixed conditions of boiling for 3 min (Table 1). Citrate and bicarbonate buffers were chosen because these buffers were used occasionally in AR by heating with paraffin-embedded sections. For most of the antibodies, 10 mM sodium citrate, pH 6.0, was as effective as distilled water as a retrieval solution. Although in the cases of CDK5 and CDC2, 100 mM sodium citrate, pH 6.0, and 10 mM sodium bicarbonate, pH 9.0, were less effective, the immunoreactivity of the other antibodies showed equivalent levels of enhancement in these buffers. In contrast, 10 mM sodium citrate, pH 3.0, was not suitable as a retrieval solution, except in the cases of GFAP and TH. Sections heated in this buffer showed uneven staining (the peripheral area of the sections was not stained although the central area was strongly stained).
Next, the heating temperature and time were assessed using 10 mM sodium citrate, pH 6.0 (Table 2). Intense nuclear staining of CDK5 was observed at temperatures of 70C and above. For PCNA immunostaining with PC10, heating at 90C and above for 3 min or at 80C for 10 min was necessary. For 19F4, heating at 90C for 10 min or boiling for 3 min was needed. For demonstration of nuclear staining of CDC2, tissues should be heated at 90C for 10 min or boiled for 3 min.
In addition, the background-lowering effect of heating, which was observed in the cases of NeuN, GFAP, and TH, was seen at temperatures of 90C and above.
Effects of AR on Free-Floating Sections
Immunostaining by the free-floating method is often used for brain sections. The effect of AR by heating on free-floating sections was examined for CDK5 and RORα (Table 3). Nuclear staining of CDK5 was not observed in untreated slide-mounted sections, as mentioned above. However, weak nuclear staining was occasionally found in untreated free-floating sections. With heat treatment either in distilled water or 10 mM sodium citrate, pH 6.0, strong enhancement of immunoreactivity was observed. In the same way, the weak nuclear immunoreactivity of RORα in free-floating sections was greatly enhanced by heat treatment.
Blocking of Endogenous Pseudoperoxidase Activity by Heating
Antigen retrieval by heating is effective for IHC with several antibodies, as described. In addition, there is one secondary beneficial effect. Endogenous pseudoperoxidase activity mainly derived from erythrocytes was completely blocked by boiling. Therefore, special treatment for blocking endogenous pseudoperoxidase was unnecessary.
Effect of retrieval solutions on antigen retrieval a
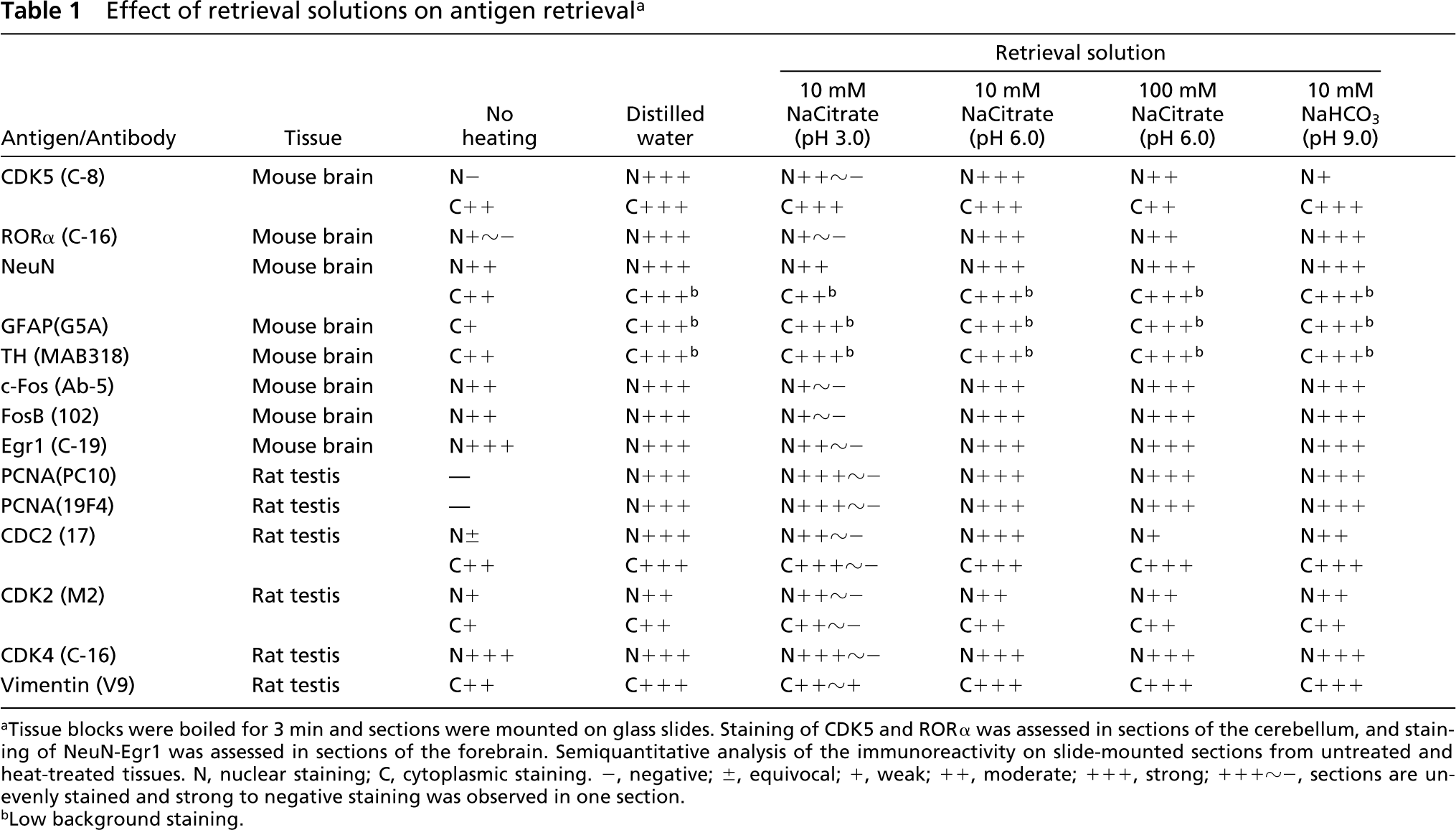
Tissue blocks were boiled for 3 min and sections were mounted on glass slides. Staining of CDK5 and RORα was assessed in sections of the cerebellum, and staining of NeuN-Egr1 was assessed in sections of the forebrain. Semiquantitative analysis of the immunoreactivity on slide-mounted sections from untreated and heat-treated tissues. N, nuclear staining; C, cytoplasmic staining. −, negative; ±, equivocal; +, weak; ++, moderate; +++, strong; +++~-, sections are unevenly stained and strong to negative staining was observed in one section.
Low background staining.
Morphological Changes of Tissues Caused by Heating
With heating, the surface of the tissue blocks underwent artifactual morphological changes, such as vacuole formation and irregular arrangement of cells (data not shown). However, the interior portions retained their normal morphology, similar to unheated tissues (compare brain tissues in Figure 1 and 2 and seminiferous tubules in Figure 3). In general, the damage caused by heating in 10 mM sodium citrate, pH 6.0 was less serious than that in distilled water. Tissues heated in a low ionic strength solution swell, possibly as a result of the imbalance in osmotic pressure, and this may be one of the factors responsible for the tissue damage.
Discussion
Preparation and rehydration of paraffin-embedded sections are sometimes laborious and time-consuming if there is no equipment such as an automated paraffin processor. The method described here enables AR by heating for cryostat sections. Advantages of the method are as follows: (a) fixed frozen sections that are slide-mounted and dried can be used; (b) no special equipment is necessary; and (c) the effect of AR is remarkable for a wide range of antibodies.
As described above, this method showed extreme effects on CDK5, RORα, PCNA, and CDC2 immunostaining, and was effective for NeuN, GFAP, TH, c-Fos, FosB, CDK2, and vimentin immunostaining. There was no remarkable effect on CDK4 and Egr1 immunostaining. Antigen retrieval methods on free-floating sections, in which tissues were heated before slicing as done here, have been reported (Evers and Uylings 1994b; Shiurba et al. 1998). However, because the applicable extent of the free-floating method is limited, the current method, which is applicable to either free-floating or dried slide-mounted sections, surpasses the previous methods. In addition, it was unexpected that heating would lower background staining in the cases of NeuN, GFAP, and TH. This may be due to the denaturation of endogenous IgG by heating, because this background-lowering effect was seen when the mouse monoclonal antibodies were used on mouse brain sections. Although heating showed no negative effect on the antibodies tested here, heating may hinder the immunoreactivity of other antibodies. When an antibody recognizes the tertiary structure of an epitope, heating of tissues may abolish this interaction. In contrast, when an antibody can recognize a denatured epitope on a Western blot, heating may not hinder immunoreactivity.
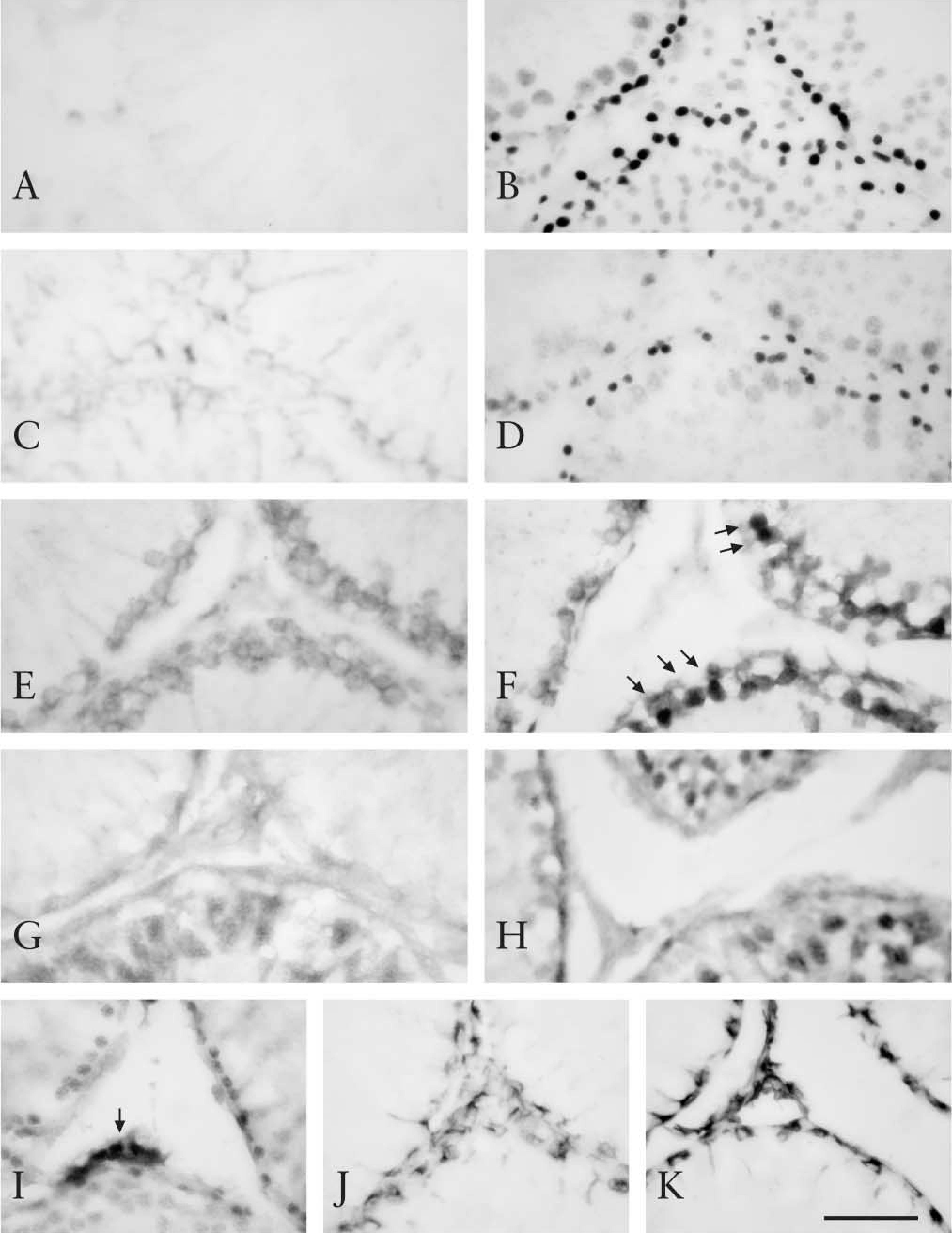
Immunostaining of PCNA (PC10;
Effect of heating temperature and time on antigen retrieval in 10 mM sodium citrate, pH 6.0 a
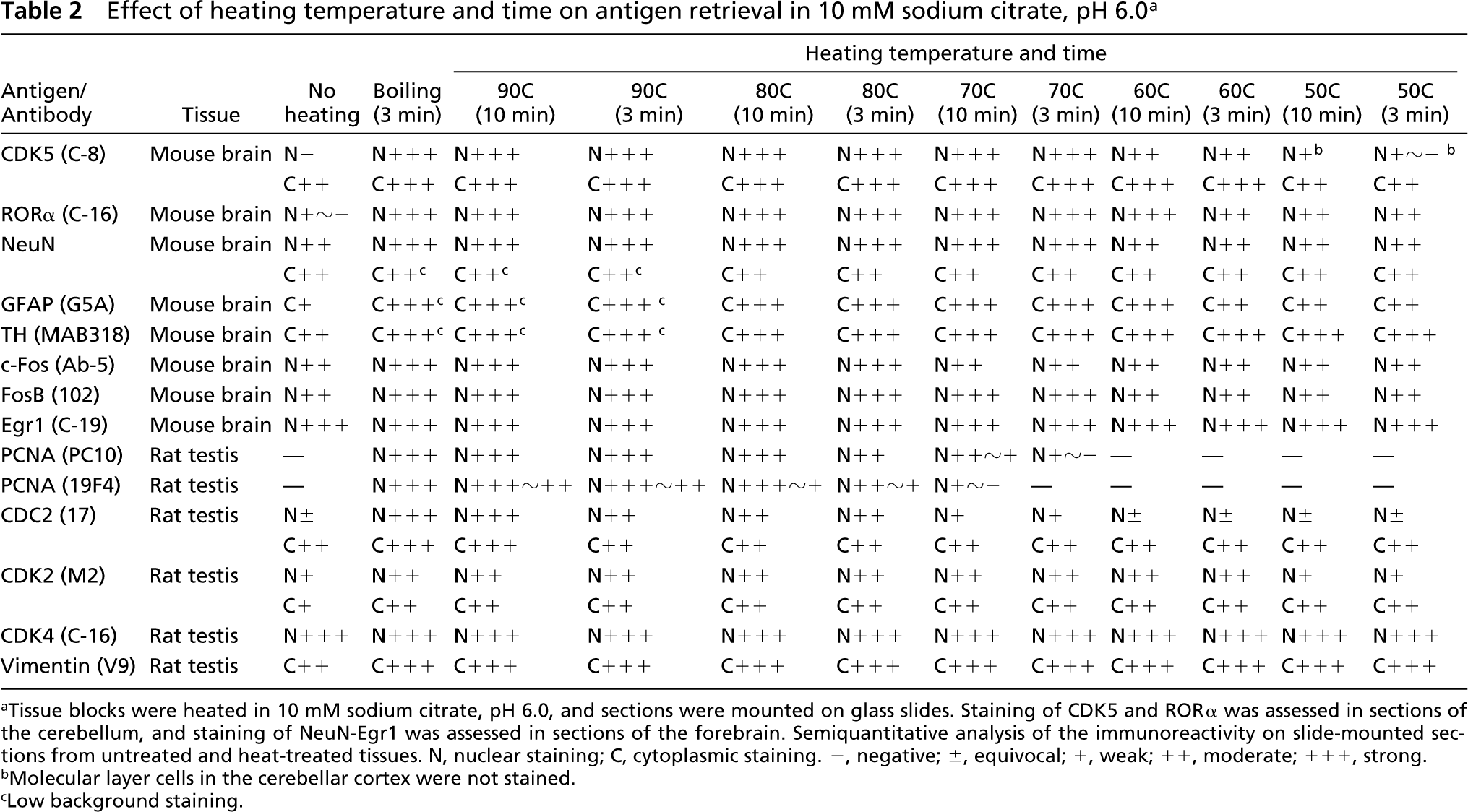
Tissue blocks were heated in 10 mM sodium citrate, pH 6.0, and sections were mounted on glass slides. Staining of CDK5 and RORα was assessed in sections of the cerebellum, and staining of NeuN-Egr1 was assessed in sections of the forebrain. Semiquantitative analysis of the immunoreactivity on slide-mounted sections from untreated and heat-treated tissues. N, nuclear staining; C, cytoplasmic staining. −, negative; ±, equivocal; +, weak; ++, moderate; +++, strong.
Molecular layer cells in the cerebellar cortex were not stained.
Low background staining.
For AR by heating in paraffin-embedded sections, factors influencing retrieval efficiency, including pH, molarity, calcium ion concentration, and chemical composition of the retrieval solution, have been examined. It has been reported that the pH is more critical than the chemical composition of retrieval solutions (Evers and Uylings 1994a; Shi et al. 1995), although there is no universally optimal pH. In the current study, the best retrieval solutions were distilled water and 10 mM sodium citrate, pH 6.0. The acidic retrieval solution, 10 mM sodium citrate, pH 3.0, was not suitable for this method. The optimal temperature was boiling or at least at 90C and above. If boiling was used, 3 min was enough for effective retrieval. However, one should examine retrieval conditions when one applies this method to other antibodies.
Antigen retrieval on free-floating sections a

Tissue blocks were boiled for 3 min and free-floating cerebellar sections were immunostained. Semiquantitative analysis of the immunoreactivity. N, nuclear staining; C, cytoplasmic staining. −, negative; +, weak; ++, moderate; +++, strong.
Heat-treated slide-mounted sections can be used for in situ hybridization and TUNEL, the technique most frequently used to detect apoptotic cells in tissue sections. Enhancement of the TUNEL reaction by microwave irradiation on paraffin-embedded sections has been reported (Sträter et al. 1995). By the method reported here, TUNEL can be performed even without protease pretreatment (Ino and Chiba 2001). Therefore, it is unnecessary to prepare another set of sections for each technique, and it is also possible to perform double staining in combination with different techniques.
In spite of the many advantages, there are some disadvantages to this method: (a) heat treatment damages tissues and (b) the heating effect is not always evenly distributed in tissues. Damage caused by heating is serious in soft tissues. However, many paraformaldehyde-fixed tissues, including the brain, spinal cord, lung, liver, thymus, spleen, kidney, alimentary tract, testis, and ovary of mice and rats, are viable after heating. Mouse and rat fetuses, at least after embryonal day 14, can be used. The heating temperature should be lowered or the heating time should be shortened for soft tissues. However, at present this method cannot be used for cultured cells. In addition, the unevenness of the heating effect in tissues is inevitable. Although the surface of the tissue is seriously damaged, its interior structure is retained. Damaged surface sections should be abandoned and undamaged sections should be collected. The AR effect on large tissues may be insufficient in the most interior part. It would be necessary to cut tissues to an appropriate size. Although there was no apparent problem in the present study using paraformaldehyde-fixed mouse brains and rat testes, the relationship of the AR effect to the size and sort of tissues and fixation conditions should be examined using larger tissues.
In conclusion, the antigen retrieval method reported here is simple and requires no special equipment. This method is useful for researchers who prefer frozen sections.