
Abstract
CSF-1 stimulates monocyte and osteoclast populations. However, the molecular mechanisms involved in regulating CSF-1 gene expression are unclear. To identify regulatory regions that control normal CSF-1 gene expression, a −774/+183-bp fragment of the murine CSF-1 promoter was analyzed in vitro and in vivo. Transcriptional activity was high in cultured osteoblasts that express CSF-1 mRNA compared to ARH-77 B cells that lack CSF-1 gene expression. Transient transfection of osteoblasts with promoter deletion constructs showed that the −774-bp fragment conferred the highest transcriptional activity and contained activator and repressor sequences. To assess the ability of the CSF-1 promoter to confer normal tissue expression of CSF-1, transgenic mice containing the −774/+183-bp region driving the E. coli β-galactosidase (lacZ) reporter gene were generated. β-Gal analysis of whole tissue extracts showed transgene expression in all tissues tested except liver and kidney. At the cellular level, the pattern of β-gal expression in the spleen, thymus, bone, lung, and testes of adult transgenic mice mimicked normal endogenous CSF-1 mRNA expression in non-transgenic littermates detected by in situ hybridization. This region also directed appropriate transgene expression to sites in other tissues known to synthesize CSF-1, with the exception of the liver and kidney. These findings indicate that the −774-bp fragment contains cis-acting elements sufficient to direct CSF-1 gene expression in many tissues. CSF-1 promoter/lacZ mice may be useful for studying the transcriptional mechanisms involved in regulating CSF-1 gene expression in tissues throughout development.
C
Studies in normal and CSF-1-deficient op/op mice have shown that CSF-1 regulates the development of tissue macrophages and osteoclasts that are involved in perinatal tissue remodeling and organogenesis (Wiktor-Jedrzejczak et al. 1990; Cecchini et al. 1994, 1997). Stimulation of resident macrophages in multiple tissues contributes to the diverse biological effects of CSF-1 on male and female fertility, trophoblast implantation, mammary gland development during pregnancy, dermal thickness, and neural function (Araki et al. 1996; Cohen et al. 1996; Pollard and Henninghausen 1996; Pollard and Stanley 1996; Pollard 1997). CSF-1 mRNA is expressed in multiple tissues in a tissue-specific manner in rodents and humans (Rajavashisth et al. 1987; Roth and Stanley 1995). In adult mice, abundant levels are detected in the heart, lung, brain, gestational uterus and, to a lesser extent, in the spleen, liver, gut, and kidney. Although the precise molecular mechanisms that control CSF-1 expression in these tissues are unknown, it is likely that shared and cell-specific transcriptional factors are involved.
In previous studies, −774 bp with respect to the transcription start site (+ 1) of the 5’ flanking region of the murine CSF-1 gene was sequenced and putative binding sites for transcription factors were identified (Harrington et al. 1991,1997). This region has only been examined in vitro using cultured osteoblasts, monocytes, and fibroblasts. Transient transfection of these cells with a series of promoter constructs showed that the −774-bp fragment conferred transcriptional activity and contained positive and negative regulatory sequences (Konicek et al. 1998; Rubin et al. 2000). In the present study, the transcriptional activity of the −774-bp fragment was assessed in vitro and in vivo. Promoter deletion constructs were tested in CSF-1-producing fetal rat calvarial osteoblast cells (FRCs) and in ARH-77 B-cells that lack CSF-1 mRNA expression. Putative activator and repressor regions were identified and tested for their functional activity in FRCs. To determine if the −774-bp region contains regulatory elements required for normal endogenous gene expression, transgenic mice harboring this fragment linked to the E. coli beta-galactosidase (lacZ) reporter gene were established. Expression of the transgene in tissues was compared to normal endogenous CSF-1 gene expression in non-transgenic controls using in situ hybridization (ISH) and to previously reported sites of CSF-1 synthesis.
Materials and Methods
Cell Culture and Northern Blot Hybridization
Primary cultures of fetal rat calvarial osteoblasts were prepared from parietal bones obtained from rat pups at 22 days’ gestation as we have previously described (Harris et al. 1994). These cells and a B-cell myeloma line, ARH-77 (ATCC; Rockville, MD), were grown in 10% FCS-containing α-MEM and RPMI 1640 medium, respectively. Total RNA was isolated from confluent cells using the RNAzol B method (Cinna Biotex; Houston, TX). Northern blots were prepared, hybridized with the full-length murine CSF-1 cDNA (Chiron; Emeryville, CA), and washed as we have previously described (Abboud et al. 1998; Ladner et al. 1998). The same filters were stripped and re-probed with 36B4 cDNA probe that detects a ubiquitously expressed ribosomal phosphoprotein (provided by Dr. Chambon; INSERUM, Strasbourg, France) (Basset et al. 1990).
CSF-1 Constructs and Transient Transfection
Deletion constructs of the murine CSF-1 5’ flanking region from −774 to −43 bp, with all containing the same 3’ end point at +183 bp, were generated by removing promoter fragments from previously constructed promoters fused to the CAT gene (obtained from M. Harrington; Indiana University, Bloomington, IN) (Harrington et al. 1991). The −774-bp fragment is with respect to the transcription start site (+ 1). Each fragment was excised from pCAT-BASIC and ligated upstream of the luciferase reporter gene in the pGL2-Basic vector. To test potential activator and repressor regions, sequences from −774 to −627 bp and from −774 to −509 bp were PCR amplified and each product containing SmaI linkers was inserted into pCR2.1 and sequenced. Separate promoter regions containing either the activator in the forward or reverse orientation, or the activator fused to the repressor, were then cloned upstream of the basal promoter in the SmaI site of the −43pGL2 construct. CSF-1 promoter constructs or the promoterless vector (l μg DNA/ well) were transiently transfected into FRC or ARH-77 cells using lipofectamine plus (Gibco Laboratories; Grand Island, NY). As an internal control for transfection efficiency, cells were co-transfected with pRL-CMV (Promega; Madison, WI) containing the Renilla luc gene linked to the CMV promoter. After 24 hr, cells were harvested and assayed for dual luciferase (firefly and renilla) activity using an assay kit (Promega). Two to three separate experiments were performed, each in triplicate or quadruplicate wells.
Generation and Analysis of Transgenic Mice
The −774 /+183-bp fragment of the CSF-1 promoter was subcloned into pBluescript at the XbaI/HindIII site. After restricting with XbaI, the fragment was blunt-ended by Klenow treatment, excised with KpnI, and then inserted into the SmaI/KpnI-digested pUC19/AUGβ-gal plasmid (obtained from Dr. Stephen Harris; UTHSC, San Antonio, TX). Transgenic mice were generated by microinjection of fertilized mouse oocytes according to standard methods (Gordon and Ruddle 1981) at the San Antonio Cancer Institute Transgenic facility. Fertilized oocytes were isolated from super-ovulated CB6F1/J (C57Bl/6 × Balb/c) mice, microinjected, and reimplanted into pseudopregnant females of the same hybrid mouse strain. Offspring were screened for the presence of the transgene by hybridizing Southern blots of tail DNA with a radiolabeled CSF-1/lacZ probe. Two founders expressed the transgene and, by Southern blotting, contained between 10 and 40 copies of the transgene per haploid genome. Animals were maintained and used according to the principles outlined in the Guide for the Care and Use of Laboratory Animals, prepared by the Institute of Laboratory Animal Resources, National Research Council.
β-Galactosidase Analysis
Tissues (100 mg each) from 7-week-old transgenic and non-transgenic mice were homogenized in 200 μl of lysis solution and β-galactosidase (β-gal) activity in 20 μl of extract was measured using the Galacto-Light Plus chemiluminescent reporter assay (Tropix; Bedford, MA). The β-gal activity of each extract was corrected for variations in protein concentration using the BCA microassay (Pierce; Rockford, IL). Tissues from six transgenic and non-transgenic mice were analyzed and values represent the level of transgene expressed minus the endogenous background levels detected in non-transgenic controls. Cellular localization of transgene expression in whole tissues of 7-week-old mice was detected by standard β-gal histochemical staining. After processing in a xylene substitute, Histo-Clear (National Diagnostics; Atlanta, GA), tissues were embedded in paraffin and 20-μm-thick sections were counterstained with 1% neutral fast red or left unstained. Non-transgenic tissues were examined in parallel as controls for endogenous β-gal activity.
ISH of Non-transgenic Tissues
A 140-bp PstI-DraI fragment of the CSF-1 cDNA inserted into a pGEM3Zf vector was used to generate antisense and sense RNA probes (Regenstreif and Rossant 1989). In vitro transcription was performed using the Riboprobe System II kit (Promega) according to the manufacturer's instructions. ISH was carried out as we have previously described (Park et al. 1998). Serial sections of tissues hybridized under identical conditions with the sense probe served as negative controls.
Results
Transcriptional Activity of the CSF-1 Promoter In Vitro
Northern blotting analysis in Figure 1A demonstrates that FRC but not ARH-77 cells constitutively express CSF-1 mRNA. FRCs express the expected 4.4- and, to a lesser extent, 2.2- and 1.4-kb transcripts for CSF-1. To examine the transcriptional activity of the −774-bp region and identify potential cis-acting elements involved in controlling CSF-1 gene expression, the cells were transiently transfected with a series of CSF-1 promoter luciferase constructs shown in Figure 1B. CSF-1 transcriptional activity paralleled CSF-1 mRNA expression, with high luciferase activity in FRCs compared to ARH-77 cells. In FRCs, deletion of DNA sequences between −774 and −627 decreased luciferase activity two- to three-fold, indicating the presence of transcriptional activators in this region. Deletion of sequences between −627 and −509 bp increased transcriptional activity, indicating the presence of negative regulatory elements. Fusion plasmids with 152 bp or less progressively diminished luciferase activity, and minimal basal promoter activity was observed with 43-bp CSF-1 promoter construct.
To confirm potential activator and repressor regions of the promoter, FRCs were transfected with constructs containing distinct promoters: CSF-1 basal promoter (p-43luc), activator region cloned in either the forward [p(−774/−627)-43luc] or reverse [p(−627/−774)-43luc] orientation upstream of the basal promoter, or the activator and repressor regions cloned upstream of the basal promoter [p(−774/ −509)-43luc]. Figure 1C shows that the activator in the forward but not the reverse orientation enhanced basal transcriptional activity approximately twofold. The effect of the activator was abrogated by the addition of the repressor region, with promoter activity of p(−774/−509)-43luc close to basal (p-43luc). This confirms the functional activity of the two regions and indicates that the repressor region has a comparatively stronger effect than the positive elements. The inability to detect the repressor in the context of the entire promoter is likely due to the interaction of proteins with DNA sequences between −509 and −;43 bp that masked the repressor effect.
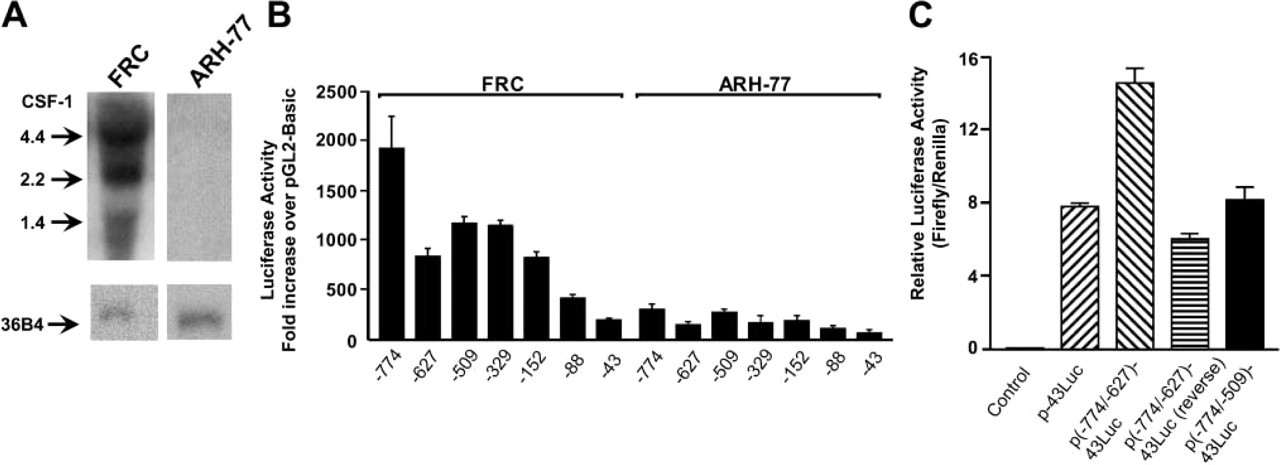
CSF-1 mRNA expression and transcriptional activity of the CSF-1 promoter in FRCs and ARH-77 cells. (
Transcriptional Activity of the −774-bp CSF-1 Promoter in Transgenic Mice
To determine if the −774/+183 bp region is also transcriptionally active in osteoblasts and other CSF-1 synthesizing cells within tissues in vivo, transgenic mice containing this same region linked to the lacZ reporter gene were generated. Two transgenic founders were identified, with both showing a similar pattern of transgene expression. These mice were phenotypically normal and one line was studied in detail. Whole tissue extracts were assayed for β-gal activity. Figure 2 shows transgene expression in the tissues analyzed, with the exception of the liver and kidney. High β-gal activity was detected in tissues that normally express abundant CSF-1 mRNA levels, such as the heart and brain.
Transgene expression at the cellular level was analyzed in hematopoietic tissues, bone, and other organs and correlated with endogenous sites of CSF-1 synthesis. The marginal zone and red pulp of the spleen (Figure 3A) showed strong β-gal staining of cells that were consistent with endothelial and T-cells. Little or no staining was present in lymphoid follicles or germinal centers of the white pulp where B-lymphocytes are concentrated. In the thymus shown in Figure 3D, scattered positive cells that likely represent epithelioreticular cells were identified in the cortical region and, to a lesser extent, in the medulla. Transgene expression in these tissues mimicked the pattern of endogenous CSF-1 mRNA expression in non-transgenic tissues analyzed by ISH (Figures 3B and 3E). In the bone marrow (Figure 3F), β-gal expression localized to cells along bone trabeculae that likely represent osteoblasts and correlated with previous ISH studies that have identified CSF-1 transcripts in osteoblasts. Figure 3G shows the lung with positive cells scattered in the peribronchial/perivascular region and along the alveolar septae. In the testes (Figure 3I), strong staining was confined to the interstitial compartment between seminiferous tubules that contains Leydig and, to a lesser extent, macrophage cells. By ISH, endogenous CSF-1 transcripts localized to similar sites in the lung and testes (Figures 3H and 3J). In contrast to these tissues, transgene expression was not observed in the liver or kidney (Figures 3K and 3M). The weak β-gal staining observed in the proximal tubules of transgenic kidneys represents endogenous β-gal activity because similar staining was also present in non-transgenic kidneys. ISH of the liver and kidney, however, identified CSF-1 transcripts in cells along the hepatic sinusoids and within renal glomeruli (Figures 3L and 3N).
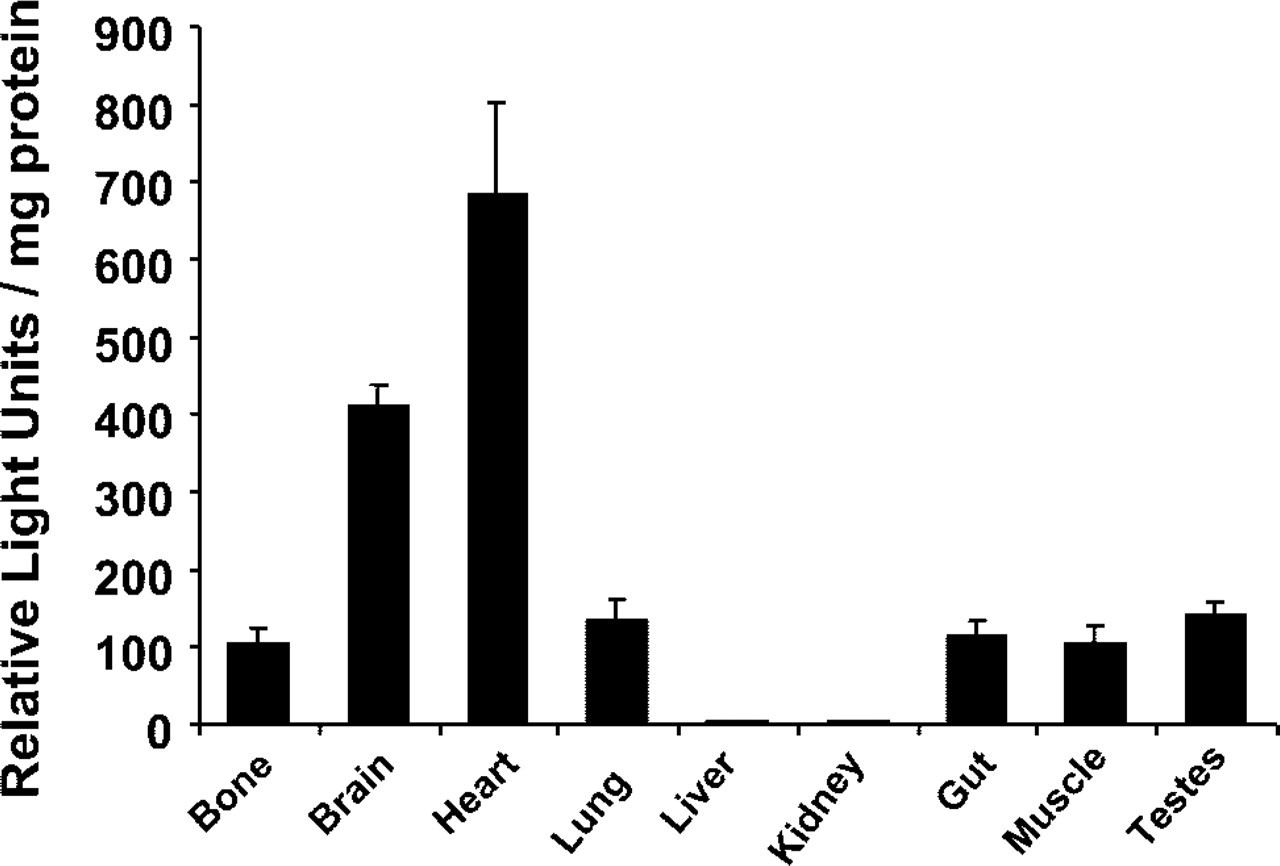
Transgene expression in CSF-1/lacZ transgenic mice. β-Galactosidase activity was quantitated in tissue extracts from 7-week-old transgenic mice and non-transgenic controls as described in Materials and Methods. Values represent β-gal levels in transgenic tissues minus endogenous levels detected in non-transgenic tissues, corrected for protein concentration of each sample (n = 6 mice/group). Data are expressed as the mean ± SE.
The −774-bp fragment also directed appropriate transgene expression to cells in other tissues known to synthesize CSF-1. Sections of the brain (Figure 4A) showed strong staining of neuronal cells in the upper granular layer of the cortex. The endocardial surface of the heart (Figure 4B) showed strong staining of the endothelial cells lining the ventricles, atria, and valves. Endothelial cells lining vascular structures in the lung, brain, skin, uterus, and ovary were also positive (Figures 3G, 4A, 4C, 4G, and 4H). In the salivary gland, there was widespread staining in serous acinar cells (Figure 4D), while sections through the gut revealed staining in epithelial cells along the base of gastric crypts (Figure 4E) and in Paneth cells located in the small intestine (Figure 4F). Although staining was also detected in cells at the neck of sebaceous glands (Figure 4C), uterine epithelial cells, and ovarian granulosa cells (Figure 4H), this reflected endogenous β-gal staining that was also observed in non-transgenic tissues.
Discussion
The present study demonstrates that the −774/+183-bp fragment of the CSF-1 promoter contains cis-acting elements that, alone, are sufficient to direct promoter activity in multiple cell types that normally express CSF-1. In vitro and in vivo transcriptional activity of this fragment correlated in two cell types, with reporter gene expression observed in osteoblasts but not B-cells. Moreover, transgene expression in mice harboring the −774-bp promoter correlated with endogenous gene expression by ISH and with previously reported sites of local CSF-1 production within many tissues.
Analysis of promoter deletion constructs in cultured osteoblasts identified activator and repressor sequences within the −774-bp fragment that may be critical for regulating CSF-1 gene expression. Since the −774-bp fragment conferred the highest transcriptional activity among the deletion constructs tested, its ability to direct CSF-1 gene expression in osteoblasts and other cells types known to synthesize CSF-1 was tested in vivo using transgenic mice. β-Gal assay showed transgene expression in many tissues that normally express CSF-1 mRNA, except for liver and spleen.
β-Gal expression in transgenic mice localized to the same cell populations in the spleen, thymus, bone, lung, and testes that were shown to synthesize CSF-1 by ISH. These findings are novel and are the first to localize the sites of CSF-1 gene expression in normal murine adult tissues. In these and other tissues, CSF-1-producing cells appeared to lie close to target cells expressing CSF-1 receptors, including macrophages and/or osteoclasts. In the spleen, CSF-1 transcripts and β-gal expression were present in cells in the marginal zone and red pulp, sites that are rich in macrophages. There was little or no staining of the white pulp of the spleen, which is composed primarily of B-cells and lacks macrophages. The thymus showed a few positive cells in the cortex and medulla, which normally contains scattered macrophages (Cecchini et al. 1994; Takahashi et al. 1994). As predicted by our in vitro studies and those of others (Rubin et al. 2000) as well as previous ISH of bone organ cultures (Hofstetter et al. 1995), transgene expression in the bone was localized to osteoblast cells, known to regulate osteoclastogenesis via the release of cytokines and through cell-to-cell contact. Although previous in vitro studies also predicted transgene expression in monocytes and fibroblasts (Konicek et al. 1998), this was not observed, indicating that in vitro and in vivo results may not always correlate. ISH signals in the lung correlated with β-gal staining in small vascular structures and scattered alveolar cells which may be involved in regulating the local macrophage population. In the testes, we observed strong endogenous CSF-1 and β-gal staining in interstitial cells that likely represent Leydig cells. These cells interact closely with adjacent CSF-1-dependent macrophages that, in turn, stimulate testosterone synthesis by Leydig cells (Cohen et al. 1996; Pollard 1997).
In several other sites, the −774-bp region also recapitulated the expected pattern of normal endogenous gene expression (Ryan et al. 2001). CSF-1 is highly expressed in the brain throughout development and promotes neuron survival, process outgrowth, and electrical conduction (Michaelson et al. 1996; Pollard 1997). Localization of transgene expression to neurons in the cortex suggests that CSF-1 may mediate these effects in an autocrine fashion. Moreover, release of CSF-1 by neurons after brain injury may stimulate surrounding microglial cells to remove damaged cells (Takeuchi et al. 2001). The presence of transgene expression in endothelial cells in the heart, brain, skin, and other tissues would be expected. Endothelial cells are the main source of circulating CSF-1 and provide a mechanism for chemotaxis and transmigration of monocytes from the circulation and macrophage activation during injury (Roth and Stanley 1992). In the uterus and ovary, endothelial cells lining vascular structures were strongly positive in the myometrium and interstitium, respectively. This correlates with studies that have localized macrophages primarily in the myometrium of non-pregnant uteri (Pollard et al. 1998). During pregnancy, the macrophage population shifts to sites beneath the luminal epithelium, which expresses high levels of CSF-1 mRNA by ISH (Pollard et al. 1987). In the gut, we identified β-gal staining in mucosal cells, suggesting that these cells may regulate macrophages known to reside in the underlying muscularis. High transgene expression in serous acinar cells of the salivary gland, which lie adjacent to macrophages, correlates with the high endogenous levels of CSF-1 reported in this tissue (Bartocci et al. 1986). Although the liver and kidney are also a source of CSF-1, expression of the transgene was not detected in these tissues. Further analysis of the CSF-1 gene should identify additional regulatory elements required for expression in the liver and kidney.
Recently, a larger 3.13-kb fragment of the mouse CSF-1 promoter and first intron (3.28 kb) linked to the lacZ gene was reported to drive normal tissue-specific CSF-1 gene expression in transgenic mice, although localization of endogenous CSF-1 transcripts by ISH was not performed (Ryan et al. 2001). In our transgenic mice that contained a shorter promoter lacking the first intron, the pattern of β-gal expression in the spleen, thymus, bone, myometrium of the uterus, testes, brain, and salivary gland was similar to that observed in mice containing the 3.13-kb promoter/intron. Mice with the longer promoter fragment also showed transgene expression in the proximal convoluted tubules of the kidney. These findings differed from our results and those of Matsuda et al. (1996), which indicated that CSF-1 mRNA expression would be expected in glomeruli. The reason for this discrepancy is unclear and remains to be determined. The cellular pattern of β-gal expression in the liver of mice containing the 3.13-kb promoter/intron was not addressed. Using ISH, endogenous CSF-1 mRNA was detected in cells along the sinusoids that likely represent liver fat-storing cells (Ito cells). We have demonstrated that Ito cells release CSF-1 that is critical for the survival of neighboring Kupffer cells (Pinzani et al. 1992).
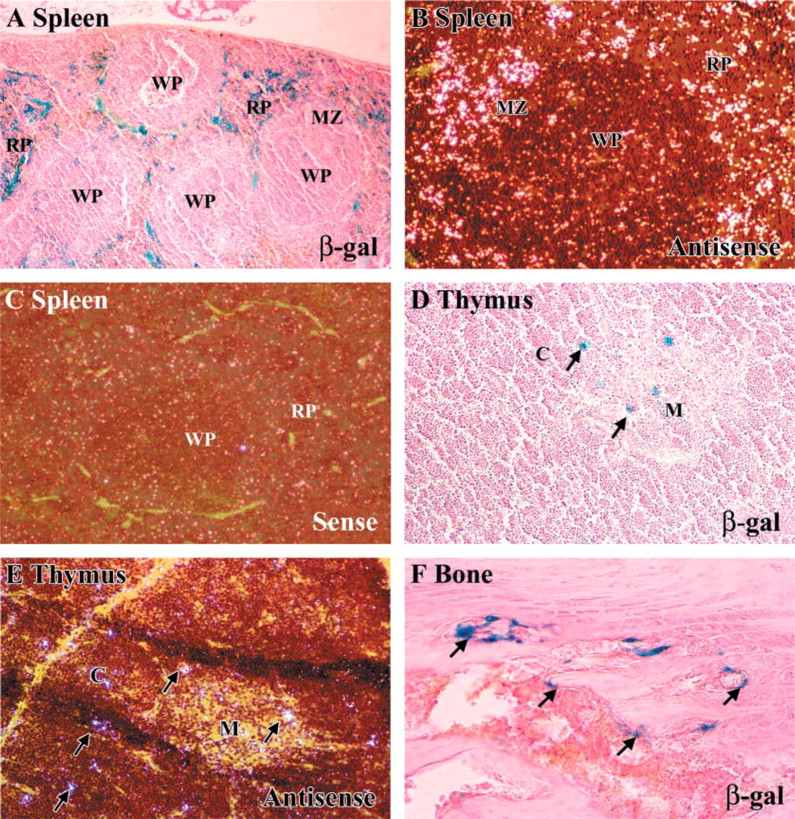
Comparative localization of β-gal expression in transgenic hematopoietic tissues, bone, and other organs using X-gal staining (
Our data indicate that the −774-bp fragment contains cis-acting elements that confer CSF-1 gene expression in multiple adult murine tissues. Studies utilizing CSF-1 promoter/lacZ mice will increase our understanding of the local biological effects of CSF-1 on target cells and may provide a model system for elucidating the molecular mechanisms involved in regulating CSF-1 gene transcription during embryonic development.
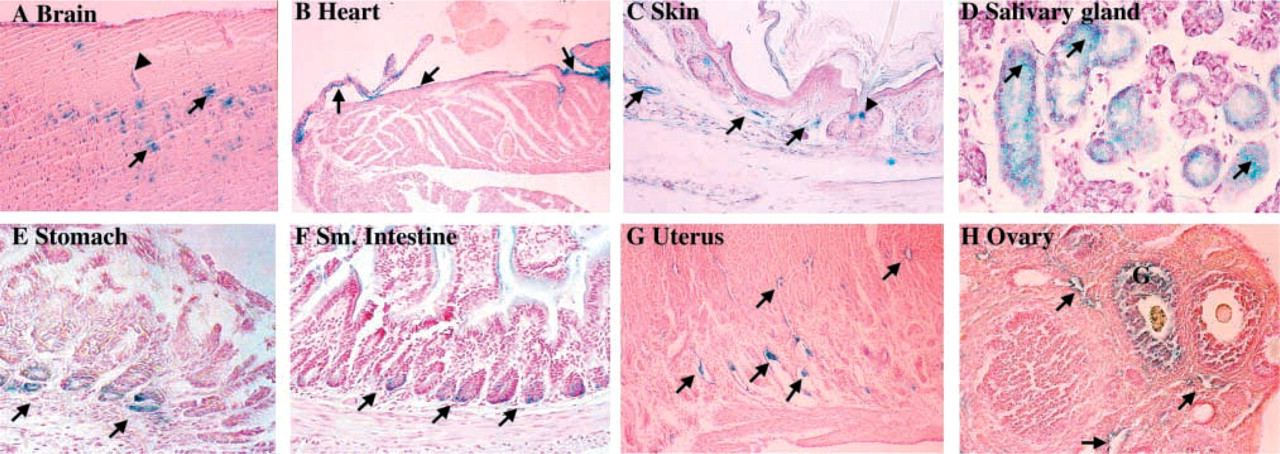
β-Gal expression in other transgenic tissues. Expression is identified in (
Footnotes
Acknowledgements
Supported in part by funding from the NIH (AR-42306, SLA), Veteran's Administration Merit Award (SLA), the Research Enhancement Award Program (REAP) from the Veterans Administration (S.L.A.), and Department of Defense (DAMD17-99-9400, NG–C).