
Abstract
Chicken embryos are an excellent model system for studies related to vascular morphogenesis. Development in ovo allows manipulations otherwise difficult in mammals, and the use of chicken-quail chimeras offers an additional advantage to this experimental system. Furthermore, the chicken chorioallantoic membrane has been extensively used for in vivo assays of angiogenesis. Surprisingly, few markers are available for a comprehensive visualization of the vasculature. Here we report the use of lectins for identification of embryonic chicken blood vessels. Nine lectins were evaluated using intravascular perfusion and directly on sections. Our results indicate that Lens culinaris agglutinin, concanavalin A, and wheat germ agglutinin can be used effectively for visualization of vessels of early chicken embryos (E2.5-E4). At later developmental stages, Lens culinaris agglutinin is a better choice because it displays equal affinity for the endothelia of arteries, veins, and capillaries. The findings presented here expand our understanding of lectin specificity in the endothelium of avian species and provide information as to the use of these reagents to obtain comprehensive labeling of the embryonic and chorioallantoic membrane vasculature.
C
Lectins are specific carbohydrate-binding proteins of nonimmune origin that have proven utility for visualization of blood vessels. In addition, lectins have been used to determine sites of leakage/increased permeability (Dvorak et al. 1988; Thurston et al. 1996,1999; Debbage et al. 2001), to identify vascular hierarchies (i.e., capillaries vs postcapillary venules) (Thurston et al. 1996; Baluk et al. 1998), and are valuable tools for the isolation of endothelial cells (Grafe et al. 1994; Christenson and Stouffer 1996; Plendl et al. 1996; Hatzopoulos et al. 1998; Iruela-Arispe et al. 1999; Gargett et al. 2000; Marelli-Berg et al. 2000; Kader et al. 2001).
Interactions of lectins with the endothelium occur through binding to specific glycoprotein moieties on the luminal and/or abluminal surface. Probably because of the great variability of sugar moieties expressed in each species, the interaction of specific lectins with endothelial cells is frequently species-specific (Roussel and Dalion 1988). Therefore, lectins that bind effectively to murine endothelial cells, such as Griffonia simplicifolia isolectin B4, do not recognize human endothelial cells (Peters and Goldstein 1979; Laitinen 1987). On the other hand, Ulex europaeus agglutinin I binds to human but not murine endothelial cells (Holthofer et al. 1982; Yonezawa et al. 1983; Iruela-Arispe et al. 1999). Consequently, comprehensive studies to address species specificity are required. Experimental evaluation of lectins in the chicken vasculature is limited to brain in late developmental stages and is restricted to a small number of lectin types (Nico et al. 1998).
To investigate the efficacy of lectins as markers for endothelial cells in chick embryos and the chorioallantoic membrane (CAM), we have examined the binding properties of nine lectins, including those frequently used for visualization of murine and human vessels. Our study has focused on lectin binding by intravascular delivery and on tissue sections of early embryos (E2.5-E7).
Materials and Methods
Chicken Embryos
Fertile White Leghorn eggs (AA Laboratory Eggs; Westminster, CA) were incubated at 38C in a humidified incubator for 48–72 hr. Eggs were opened into 100-mm tissue culture Petri dishes and allowed to develop in a humidified tissue culture incubator at 38C for the times indicated.
Intravital Lectin Perfusion
Fluorescein isothiocyanate (FITC)- and rhodamine-conjugated lectins (Vector Laboratories; Burlingame, CA) were diluted in 1 × PBS and filtered using a 0.22-mm Millipore filter. The solution was warmed to 37C before perfusion. Intracardiac (before E5) or vitelline vein (after E5) injection was performed with the aid of a glass needle and perfusion was achieved by cutting the main yolk artery and allowing solutions to circulate for 3–5 min. A total of 0.2-2.5 μg of lectin was delivered in 2-6 μl depending on the developmental stage. The injection volume was kept to a minimum to avoid rupture of vessels due to volume overload. Pressure was maintained constant by use of a micropump (Harvard Apparatus; Holliston, MA): 50 μl/min before E3 and 100 μl/min thereafter. Lectins used included concanavalin A (ConA), Griffonia simplicifolia isolectin B4 (GSL I-B4), Euonymus europaeus (EE), Lens culinaris agglutinin (LCA), Lycopersicon esculentum (LE), peanut agglutinin (PNA), Ricinius communis agglutinin I (RCA I), Ulex europaeus agglutinin I (UEA I), and wheat germ agglutinin (WGA). When co-injection of two lectins was required, preincubation was performed and monitored under the microscope for possible precipitation. Two controls were performed to ensure lectin specificity: (a) perfusion of unlabeled lectin before injection of fluorophore-conjugated lectin, resulting in complete blockade of binding by fluorophore-conjugated lectin; and (b) tagged lectin together with the cognate hapten sugar moiety (as indicated in Table 1). For these competition experiments, a concentration of 0.2 M of the appropriate sugar was used either on sections (for 2 hr at room temperature) or by intravascular perfusions (1 ml of the particular sugar before lectin injection). In these experiments we saw either a significant decrease in staining or complete inhibition.
For the purpose of comparison, LM609 antibody (anti-αvβ3) (Chemicon; Temecula, CA) and anti-VEGFR2 (Eichmann et al. 1997) were included in these studies and visualized by incubation of secondary antibodies conjugated to Cy5. Anti-quail VEGFR2 antibodies (hybridoma supernatant) were also used on frozen sections (20 μm) of E2 embryos. Antibodies were applied overnight at 4C followed by biotinylated goat anti-mouse IgG1 and streptavidin-Cy5.
Fixation and Sectioning of Embryos
After perfusion, embryos were fixed in 4% paraformaldehyde overnight at 4C and then washed with 1 × PBS three times. Embryos at or younger than E4 were mounted on histological slides using Vectashield (Vector Labs). At E5 and thereafter, embryos were sectioned at 200-μm thickness using a Vibratome (Ted Pella; Redding, CA) and mounted with Vectashield. Evaluation was performed on a MRC-1024ES confocal system (Bio-Rad Laboratories; Hercules, CA) equipped with a Nikon E800 microscope and a krypton/argon laser.
Lectins used and their carbohydrate specificities
aModified from Damjanov (1987).
bModified from Goldstein and Poretz (1986).
Lectin Histochemistry
For evaluation of lectins by direct staining of sections, E7 embryos were fixed in 4% PFA overnight at 4C and sectioned at 200 μm using a vibratome. Sections were incubated with 0.3% Triton in 1 × PBS for 30 min and were then washed to remove excess of detergent. Lectins (5 μg/ml) were incubated for 1 hr at RT and washed extensively for 16 hr.
Results
Nine lectins (ConA, GSL I-B4, EE, LCA, LE, PNA, RCA I, UEA I, and WGA) were selected to evaluate their utility for labeling embryonic chicken vessels. Table 1 includes their known carbohydrate specificity and biological source.
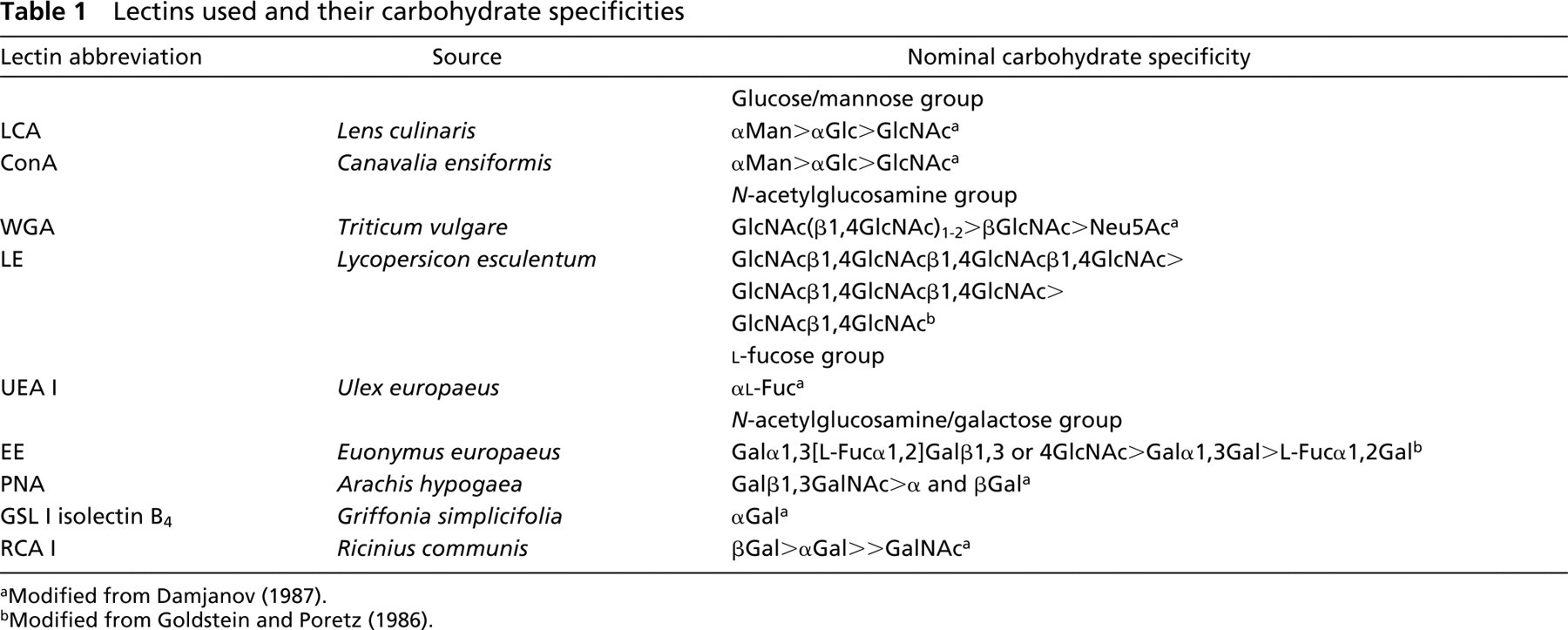
Initial screening was performed by intravascular perfusion of E2.5-E3 chicken embryos. In preliminary experiments, we found that LCA and WGA bound well to the luminal surface of vessels, in sharp contrast to UEA I and GSL I-B4, lectins previously known for their ability to label endothelial cells of human and murine vessels, respectively. In subsequent experiments we included either LCA or WGA in combination with other lectins to ensure that perfusion was complete and to compare patterns of binding. The selection of either LCA or WGA was based on compatibility with other lectins. For example, LCA mixed with RCA I resulted in the formation of insoluble precipitates that clogged small vessels during perfusion. Therefore, formation of precipitates was determined by microscopic evaluation and centrifugation before injection into the embryo. Figure 1 shows affinity of each lectin to the endothelium of chickens at E3. For evaluation of uniformity of binding to small- and large-caliber vessels, we decided to focus in the inter-somitic area. It should be stressed that although other areas are not shown, binding or lack thereof was uniform and consistent throughout the embryo. In addition to LCA and WGA, ConA (Figures 1B and 1C) and RCA I (Figures 1Q and 1R) bound to the vasculature of early chicken embryos quite well. LE, a lectin routinely used for perfusion of mouse embryos, did not bind as strongly as expected (Figures 1H and 1I). Because it was conceivable that this profile of binding changed over time, we performed similar experiments at later developmental stages (E4, E5, E6, and E7), as discussed below.
To compare lectin labeling efficacy with a set of known endothelial cell markers, embryos at the same developmental stage were also perfused with anti-αvβ3 and anti-VEGFR2 (Figures 1V and 1W). Although weak binding was detected, labeling was generally not as complete or as intense as that seen with LCA, ConA, WGA, or RCA I (Figure 1). Additional control experiments included perfusion with (a) unlabeled lectin before injection of fluorophore-conjugated lectin and (b) tagged lectin together with the cognate hapten sugar moiety (as indicated in Table 1). Both these controls resulted in complete and selective blockade of lectin binding (data not shown).
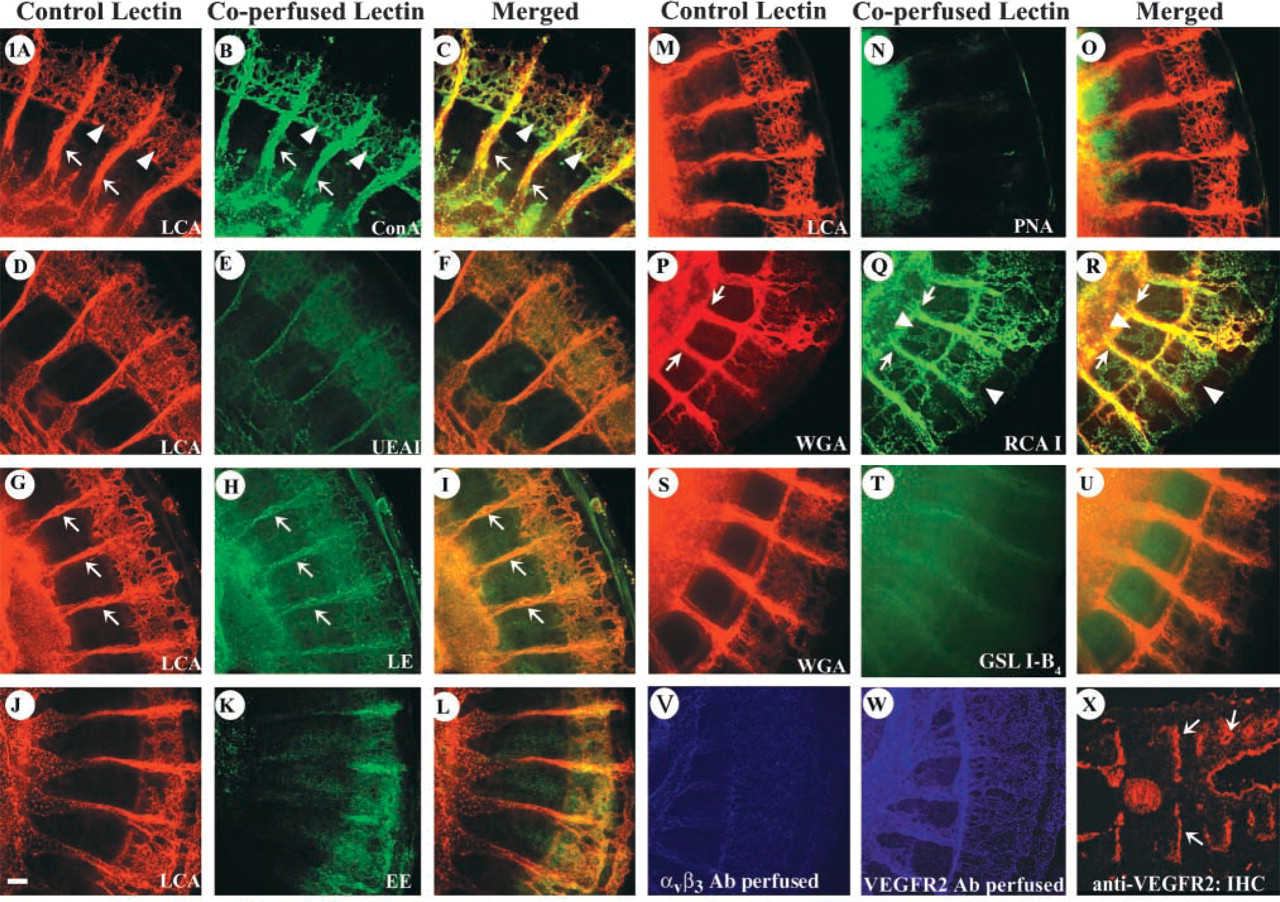
In addition to intravascular delivery, we tested the performance of all nine lectins on tissue sections (Figure 2). Although LE performed poorly when delivered intravascularly, it displayed effective binding of vessels on tissue sections of older embryos (Figure 2A). Surprisingly, WGA did not bind either large- or small-caliber vessels (Figure 2B). We believe that this was the result of delivery (i.e., on sections vs intravascular), as labeling with this lectin was seen on prefixed perfused vessels (Figure 2J). Lack of binding was also seen with UEA I and PNA (Figures 2C and 2D). RCA I, GSL I-B4, and EE bound to capillaries and larger vessels, albeit with lower affinity than LE (Figures 2E-2G). LCA and ConA also labeled vessels on sections (Figures 2H and 2I). Nevertheless, in all cases we observed significant background and frequent binding to either epithelium and/or extracellular matrix. Clearly, intravascular injection offered a more selective and intense vascular labeling.
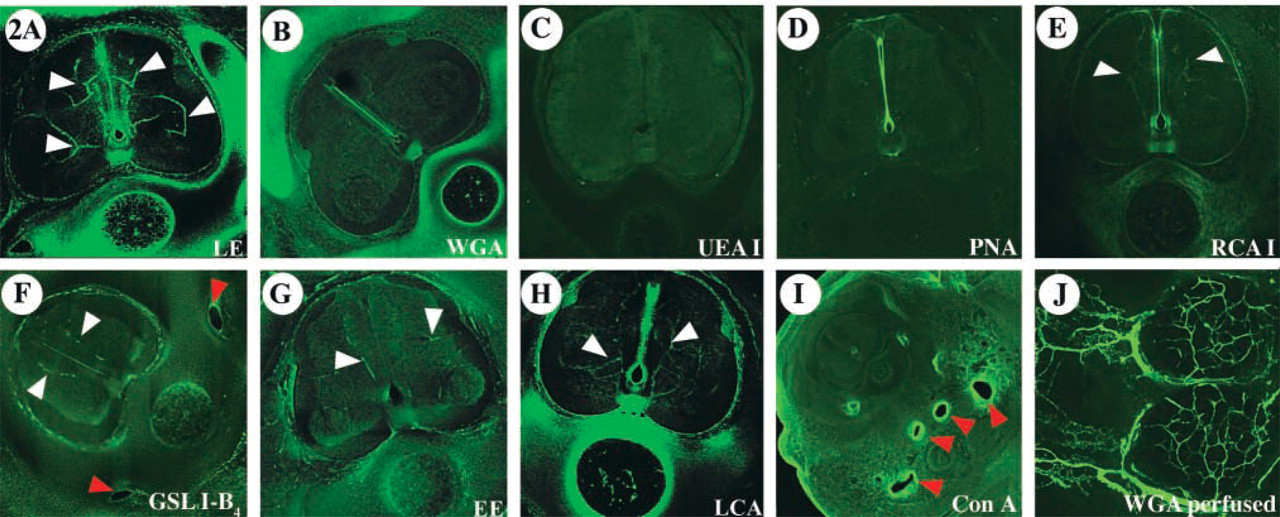
To identify possible variations in lectin affinity that might occur as a consequence of developmental changes, LCA and WGA binding patterns were compared at several stages. Co-injection of these two lectins was performed in embryos at 2.5, 3, 4, 5, and 7 days of development (Figure 3). During early stages of development (Figures 3A-3I), both lectins labeled the majority of vessels with equal affinity. However, as a result of vascular expansion, maturation, and remodeling during the developmental stages studied (E5 and E7), WGA showed a progressively more restricted affinity for larger vessels, while LCA continued to display comprehensive binding to the vasculature (Figures 3J-3O). Specifically, WGA exhibited higher affinity for some medium- and large-caliber vessels in detriment to small capillaries. Compared to WGA, LCA bound to the luminal surface of large and small vessels more uniformly and revealed the complete vascular network.
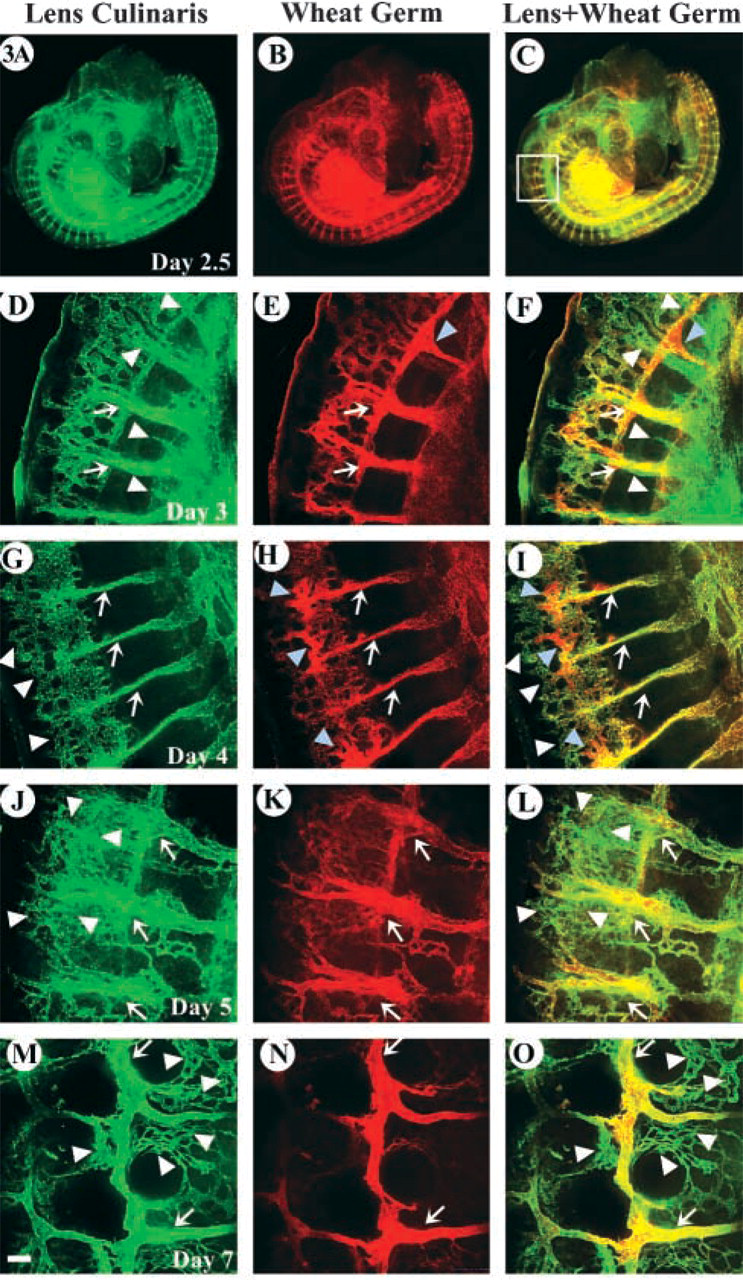
Efficiency of LCA and WGA binding in older embryos was determined on tissue sections after intravascular perfusion. Embryos (E7) were embedded in agarose for generation of 200-μm sections on a vibratome. Sections were mounted on slides and observed by confocal microscopy. Figure 4 shows a cross-section of E7 after co-injection with FITC-LCA and rhodamine-WGA. The evaluation confirmed previous results and demonstrated that smaller vessels are not as consistently labeled with WGA. In contrast, LCA was able to bind uniformly to endothelial surfaces of different hierarchic vascular networks. Furthermore, tissue sections allowed the clear identification of arteries and veins that appear to interact equally with LCA (Figure 4D). Interaction of venular endothelium with WGA, however, appeared much weaker than its binding to arterial endothelium at this developmental stage (Figure 4E).
Affinity of lectins for the luminal surface of embryonic chicken endothelium. (
Lectin staining of embryonic sections is less effective than intravascular perfusion for visualization of blood vessels. Transverse sections of chicken embryos (E7) were incubated with FITC-conjugated lectins and evaluated by confocal microscopy. Shown are images of the dorsal region, particularly the spinal cord and notochord. (
Binding of WGA and LCA to CAM vasculature is shown in Figure 5. These findings were similar to those in the intraembryonic vasculature. We found that LCA labeled CAM vessels uniformly (Figures 5A, 5D, and 5G). Binding was detected on vessels of different caliber and variable developmental stage. In contrast, WGA was recruited more selectively to larger vessels (Figure 5H) or to areas in which the vascular plexus appeared to undergo remodeling and fusion (Figures 5B and 5E). Incubation of E7 CAM vasculature with anti-αvβ3 showed binding to small-caliber vessels and adventitial capillaries of larger vessels (Figure 5J). Interestingly, terminal branches of small-caliber vessels were not recognized by αvβ3 as they were with LCA.
Discussion
Visualization of the vasculature in chicken embryos has met with some difficulties owing to the lack of specific reagents. Although PECAM is commonly used for both human and mouse vessels, the antibodies available are species-specific and do not crossreact with chicken vessels. The development of integrin antibodies has been particularly useful for functional analysis. For example, the monoclonal antibody LM 609 directed to the integrin αvβ3 was injected into developing quail embryos, resulting in significant disruption of capillaries and abnormal vascular patterning (Drake et al. 1995). This reagent, however, has offered limited applicability for comprehensive visualization of blood vessels. Currently, several molecules have been shown to be expressed selectively in arterial or venous endothelial cells and label the chicken embryonic vasculature. These include neuropilins (Herzog et al. 2001) and the transmembrane ligand ephrin B2 (Moyon et al. 2001; Othman-Hassan et al. 2001) for arteries. In contrast, EphB4 and Tie 2 were found to be expressed in veins (Moyon et al. 2001). Clearly, a marker that binds uniformly to all endothelial cells is lacking. Although efforts to generate chicken-specific antibodies to VEGFR2 are ongoing, these antibodies are not yet commercially available and recognition requires the use of frozen sections (Eichmann et al. 1997).
Lectins are proven tools for comprehensive visualization of the vasculature in several organs and animal species. The most successful approach has been intravascular delivery of the lectin. This method bypasses the problem of cell-type specificity because exposure of the lectin becomes limited to the luminal endothelial surface (Thurston et al. 1998,1999; Murphy et al. 1999). Lectins bind to sugars in either glycolipids or glycoproteins (Goldstein and Poretz 1986; Sharon and Lis 1989). Consequently, their affinity is broad and is defined by the monosaccharide that is able to inhibit binding (So and Goldstein 1967; Lloyd et al. 1969; Damjanov 1987). This type of chemical affinity explains the lack of consistency in the array of lectins that bind to the endothelium of different species (Fatehi et al. 1987; Roussel and Dalion 1988; Bankston et al. 1991; Rodriguez-Manzaneque et al. 2001). In addition, we have observed differences in vessel binding affinity when lectins were applied on sections vs intravascular perfusion. The reasons for altered affinity on sections vs in vivo (by intravascular perfusion) are multiple. For example, sections enable exposure to both surfaces of the endothelium and in some cases, interaction might occur with components of the basement membrane. In addition, postfixation might alter the conformation of binding sites that can consequently become more exposed due to crosslinking of aldehydes. We and others have found that binding of GSL I-B4 to paraffin sections of paraformaldehyde-fixed murine brain capillaries is enhanced if the tissue is previously digested with trypsin or proteinase K. In the case of sections from embryonic chicken, digestion did not appear to alter any of the binding patterns reported (data not shown). In this regard, the present study provides further strength to the notion that interactions of lectins with the endothelium are species-specific.
WGA binding profile is altered during development in contrast to LCA affinity, which remains constant. Embryos (E2.5) were co-perfused with FITC-LCA and rhodamine-WGA and either directly mounted on slides (E2.5-E4) or sagitally sectioned (
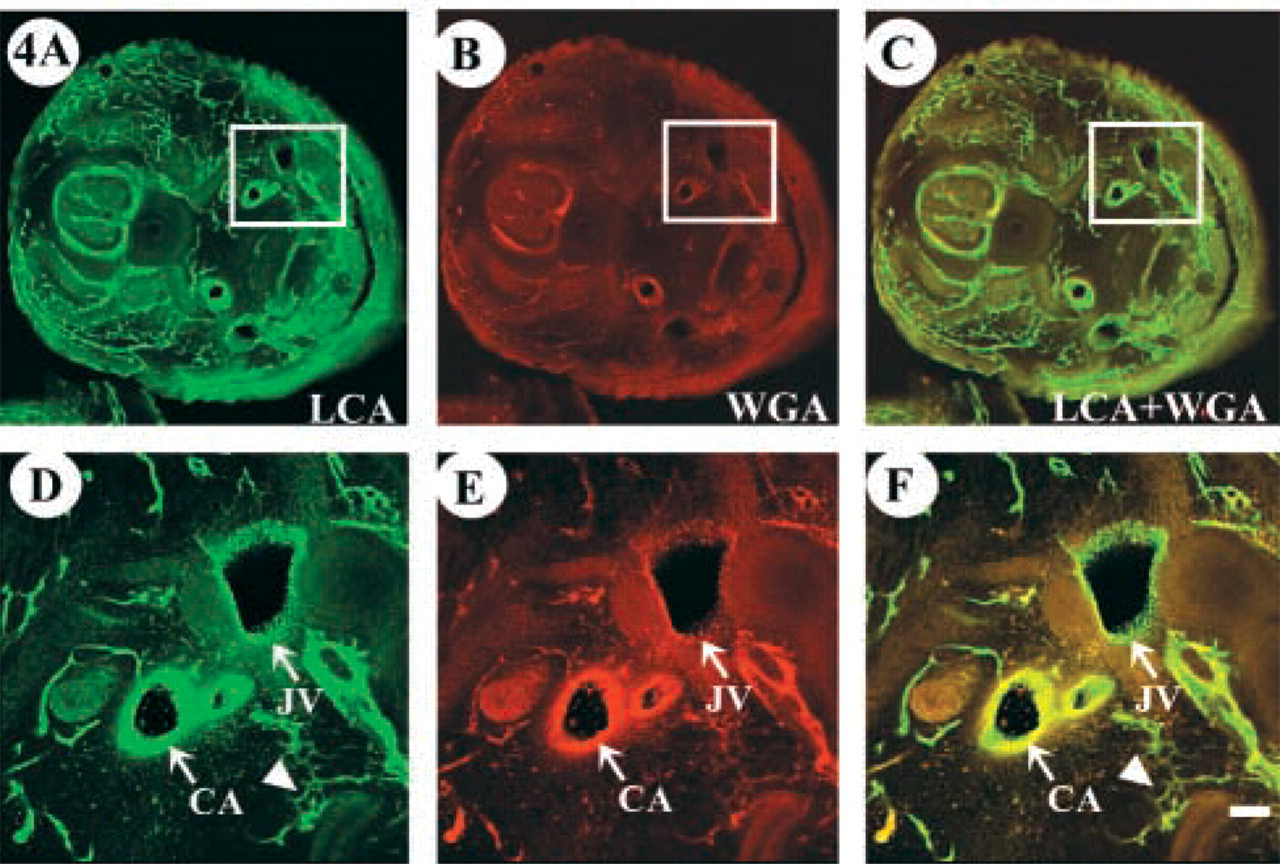
Although LCA comprehensively binds vein, artery, and capillaries, WGA appears to have a more limited affinity for different vascular hierarchies. Transverse tissue sections of cervical region of embryo (E7) after perfusion with the indicated lectins. (
It is unclear which molecule(s) harbors the binding of lectins on the surface of the endothelium. Experiments performed with bovine aortic endothelial cells have indicated that WGA is able to bind to VEGFR2 (Vaisman et al. 1990). We have been able to confirm these results with chicken endothelium using WGA and LCA in pull-down experiments (data not shown). Although VEGFR2 might be an important component of the array of proteins bound to the lectins, there are certainly many other molecules that facilitate lectin interaction with the endothelium. LCA is found to interact with glucose residues, whereas WGA binds N-acetyl-glucosamine as well as sialic acid linked to N-acetyl-glucosamine (Damjanov 1987; Thurston et al. 1996). Given this affinity, it is likely that multiple glycoproteins provide binding sites on the surface of the endothelium. Because of the binding of these lectins to the surface of sugar moeties in important receptors such as VEGFR2, as well as other glycoproteins, LCA and WGA should not be used for vital labeling of the vasculature because their binding will most likely affect the response to VEGF and vascular morphogenesis.
An interesting observation was the gradual change in the affinity of WGA from a more general capillary plexus in early stages of development, to an increasingly restricted binding to larger vessels. It has been recognized that the pattern of lectin binding sites on migrating endothelial cells is both increased and altered compared to quiescent cells (Augustin-Voss and Pauli 1992). Angiogenesis does encompass active migration, and it is likely that quiescent adult endothelium might not interact with the same types of lectins. In fact, changes in lectin specificity have been described in the human liver as a factor of development (Quondamatteo et al. 1997). Our studies have included a relatively limited sample of developmental stages (E 2.5-E7). Therefore, additional investigations should be performed to determine whether late developmental stages and/or adult chicken endothelium have the same properties as the ones described by the present study. Nevertheless, we hope that this study will enable investigators to comprehensively visualize chicken vasculature using commercially available means.
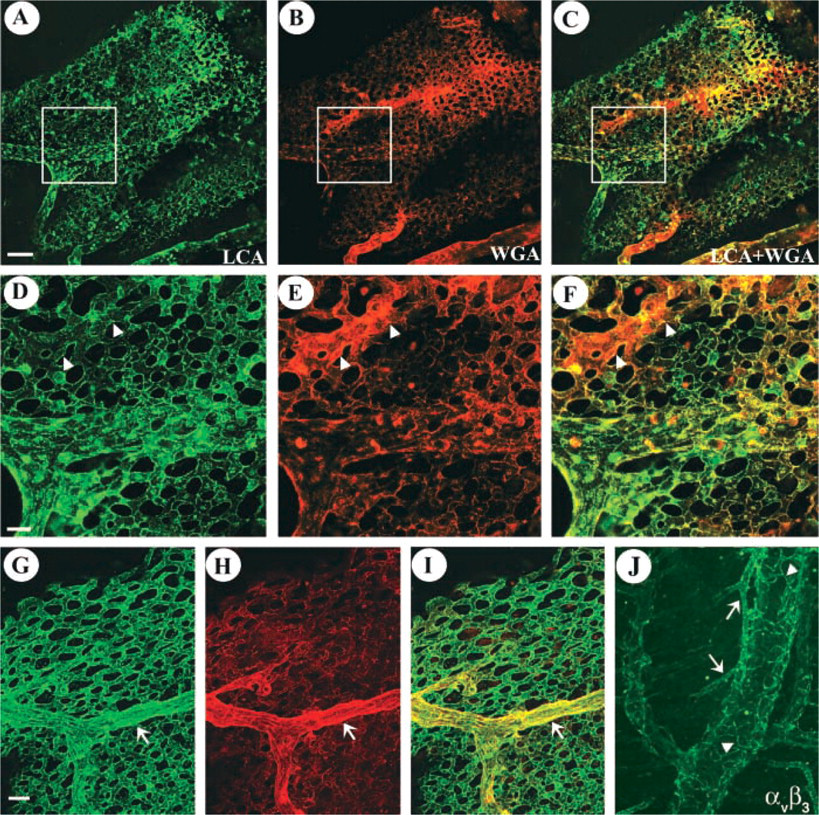
Differences in the intraembryonic vascular labeling profiles of LCA and WGA are also apparent in the CAM vasculature. Dual lectin perfusion, LCA (green) and WGA (red), was performed on embryos (E12) and labeling of CAM vessels was evaluated. (
Footnotes
Acknowledgements
Supported by a grant from the AHA Western affiliates to MLI-A.