
Abstract
Almost all secreted proteins pass through the endoplasmic reticulum (ER), an organelle that is equipped to tolerate and/or degrade misfolded proteins. We report here that yeast expressing the cystic fibrosis transmembrane conductance regulator (CFTR) concentrate the protein at defined sites in the ER membrane that are not necessarily enriched for the ER molecular chaperone BiP. We propose that these sites are Russell bodies, an ER subcompartment in which misfolded proteins are stored and can be targeted for degradation.
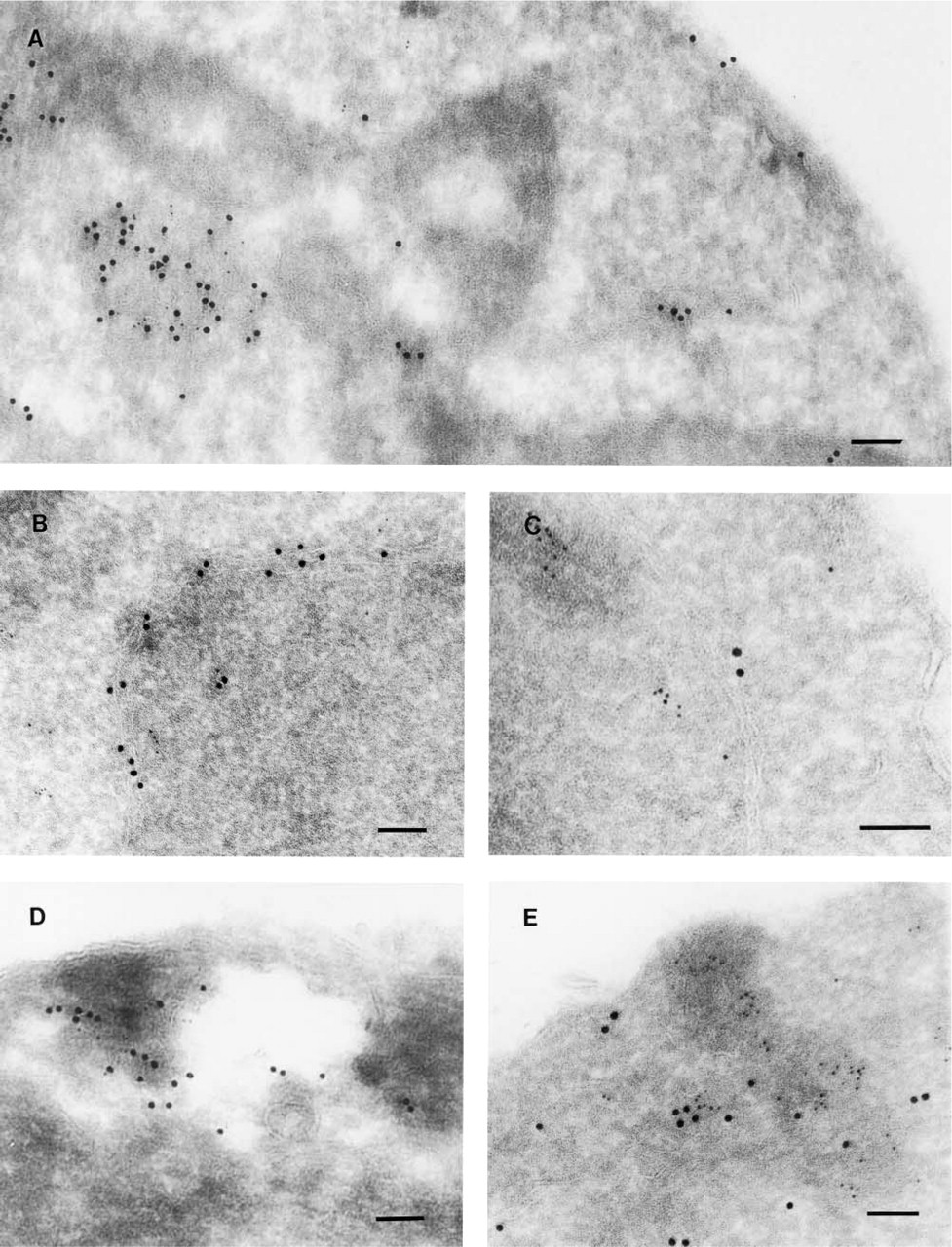
A
Among their many functions, molecular chaperones catalyze protein degradation. To determine whether ER-associated chaperones are required for wild-type CFTR degradation, we expressed the protein in the yeast S. cerevisiae and reported that the activity of the ER luminal Hsp70 chaperone BiP was dispensable for CFTR degradation, although indirect immunofluorescence (IF) microscopy suggested that CFTR resided in foci that co-localized with BiP (Zhang et al. 2001). To examine whether the CFTR foci represented “BiP bodies” or “Russell bodies”—ER subcompartments in which misfolded secretory proteins accumulate and in which BiP is either enriched or excluded, respectively (Nishikawa et al. 1994; Kopito and Sitia 2000)—we analyzed the localization of CFTR with respect to BiP by immuno-EM because of the limited resolution of IF in yeast.
To maximize epitope accessibility, cryosections of yeast expressing hemaglutinin (HA)-tagged wild-type CFTR were prepared with minimal fixation (Byers et al. 1991; Wright 2000). In brief, washed cells were fixed in cold 2% paraformaldehyde/0.01% glutaraldehyde in 0.1 M PBS for 1 hr, embedded in 3% gelatin, fixed for 30 min, and infused overnight either with 2.3 M sucrose/0.1 M PBS or with 20% polyvinylpyrrolidone/1.6 M sucrose/0.1 M PBS buffered with 0.055 M sodium carbonate. Blocks were frozen on specimen stubs in liquid nitrogen and 300-nm sections were cut on a Reichert Ultracut FCS and cryokit at −95C, stained with 0.5% Toludine Blue, and examined under the light microscope. Next, 65-nm sections were cut at −120C and mounted on 0.125% formvar-coated grids. The sections were rinsed in 0.1 M PBS and then 0.5% BSA in 0.15 M glycine, blocked with goat serum, and incubated with anti-HA (1:50; Roche, Chicago, IL) and anti-BiP antibodies (1:250; Brodsky et al. 1993) or anti-Pep12p antibodies (1:2000; Becherer et al. 1996) for 1 hr. Anti-HA and anti-BiP/Pep12p were visualized with 5-nm gold conjugated goat anti-mouse IgG and 10-nm gold-conjugated goat anti-rabbit IgG, respectively. The sections were fixed with 2.5% glutaraldehyde, counterstained with uranyl acetate, mounted in 1% methylcellulose, and dried. Labeled sections were examined on a JEOL 1210 transmission electron microscope.
BiP resided in intracellular membranes (Figure 1, large particles) and on occasion CFTR (small particles) co-localized with BiP (e.g., Figure 1A) at what may be ER translocation sites (Brodsky et al. 1993). More commonly, CFTR clusters excluded BiP (e.g., Figures 1B, 1C, and 1E). Morphometry of these and other images indicated that ∼72% of the CFTR-corresponding particles were clustered in regions lacking BiP. Because CFTR was membrane-associated or in more densely staining membrane-enriched regions, we conclude that CFTR does not reside in “aggresomes” in yeast, which are perinuclear cytoplasmic protein aggregates (Kopito and Sitia 2000). This conclusion is supported by data indicating membrane association of CFTR (Zhang et al. 2001). We also noted that CFTR residence was distinct from that of Pep12p, a protein that resides in the endosome/late-Golgi stack in yeast (Figure 2; Becherer et al. 1996). This result is consistent with the inability of CFTR to transit beyond the ER in yeast (Zhang et al. 2001).
(
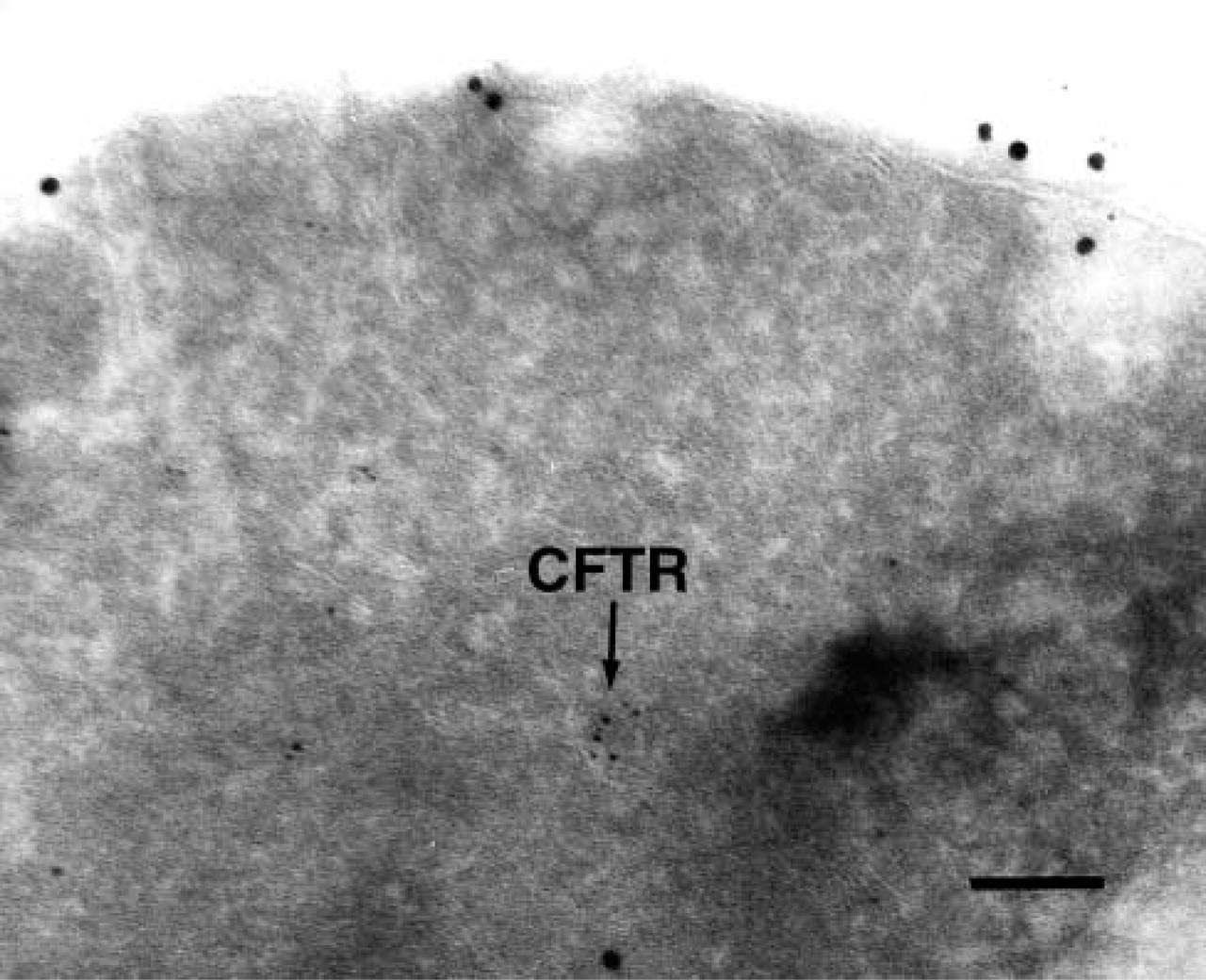
Distinct residence of CFTR and the endosome/late-Golgi Pep12 protein in yeast. Sections were prepared for immuno-EM as described in the text. Pep12p, 10-nm particles; CFTR, 5-nm particles. Bar = 0.1 μm.
To further examine BiP and CFTR residency, we fractionated yeast membranes on sucrose density gradients (Hong et al. 1996). ER-enriched fractions (Figure 3, Lanes 1–7) that contained CFTR, the Sec61p translocation channel and BiP, and plasma membrane fractions (Figure 3, Lanes 11–13) that contained the Pma1p ATPase were each concentrated by centrifugation at 100,000 X g for 30 min. The resulting pellets were then prepared for immuno-EM. Although we failed to detect CFTR or BiP in plasma membrane-enriched fractions (data not shown), thereby providing a negative control for these experiments, we identified several micrographs from the ER-enriched fractions that harbored CFTR but in which BiP was absent (see Figure 4 for one example).
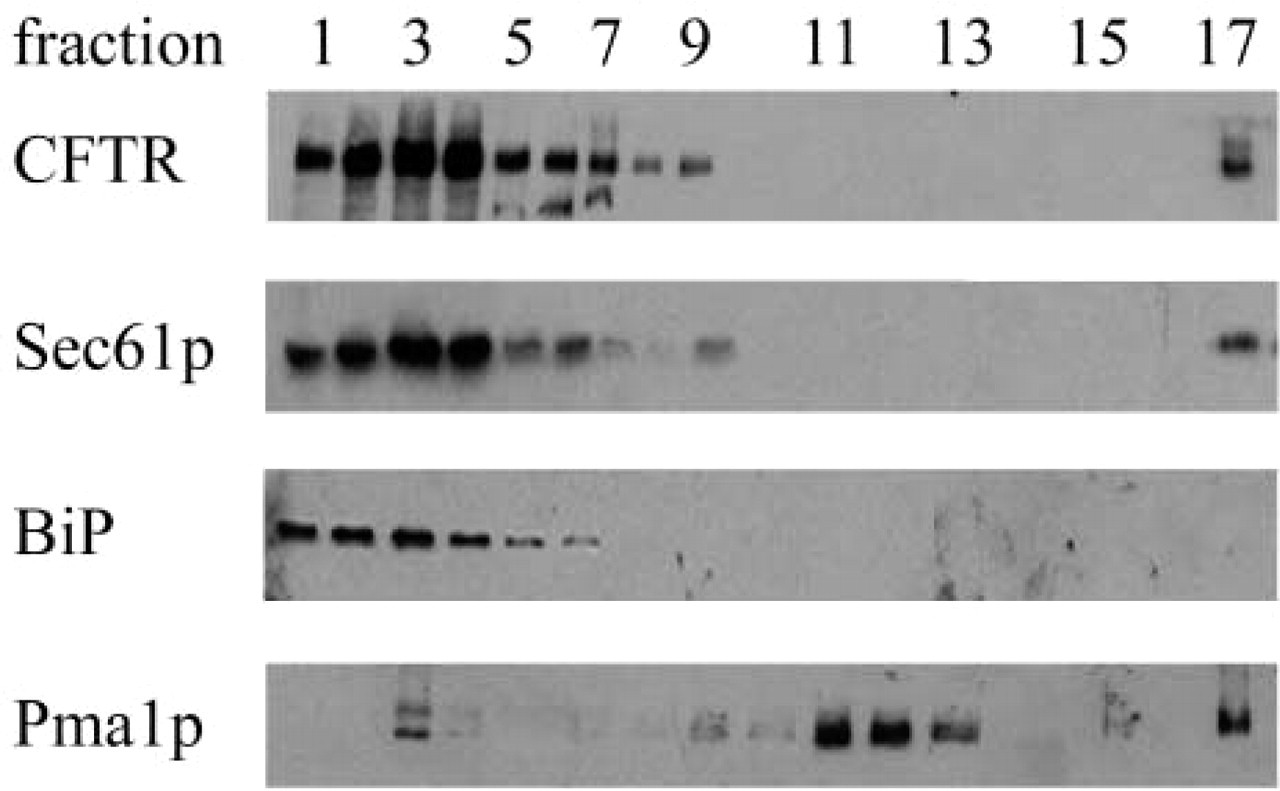
Lanes 1–17: Identification of ER- and plasma membrane-enriched fractions. Extracts from HA-CFTR-expressing yeast were subjected to sucrose gradient centrifugation. The gradient was fractionated and the migrations of CFTR, Sec61p, BiP, and Pma1p were determined by immunoblotting analysis (Brodsky et al. 1993; anti-Pma1p was provided by Dr. Carolyn Slayman, Yale University). Fraction 1 represents the top of the gradient.
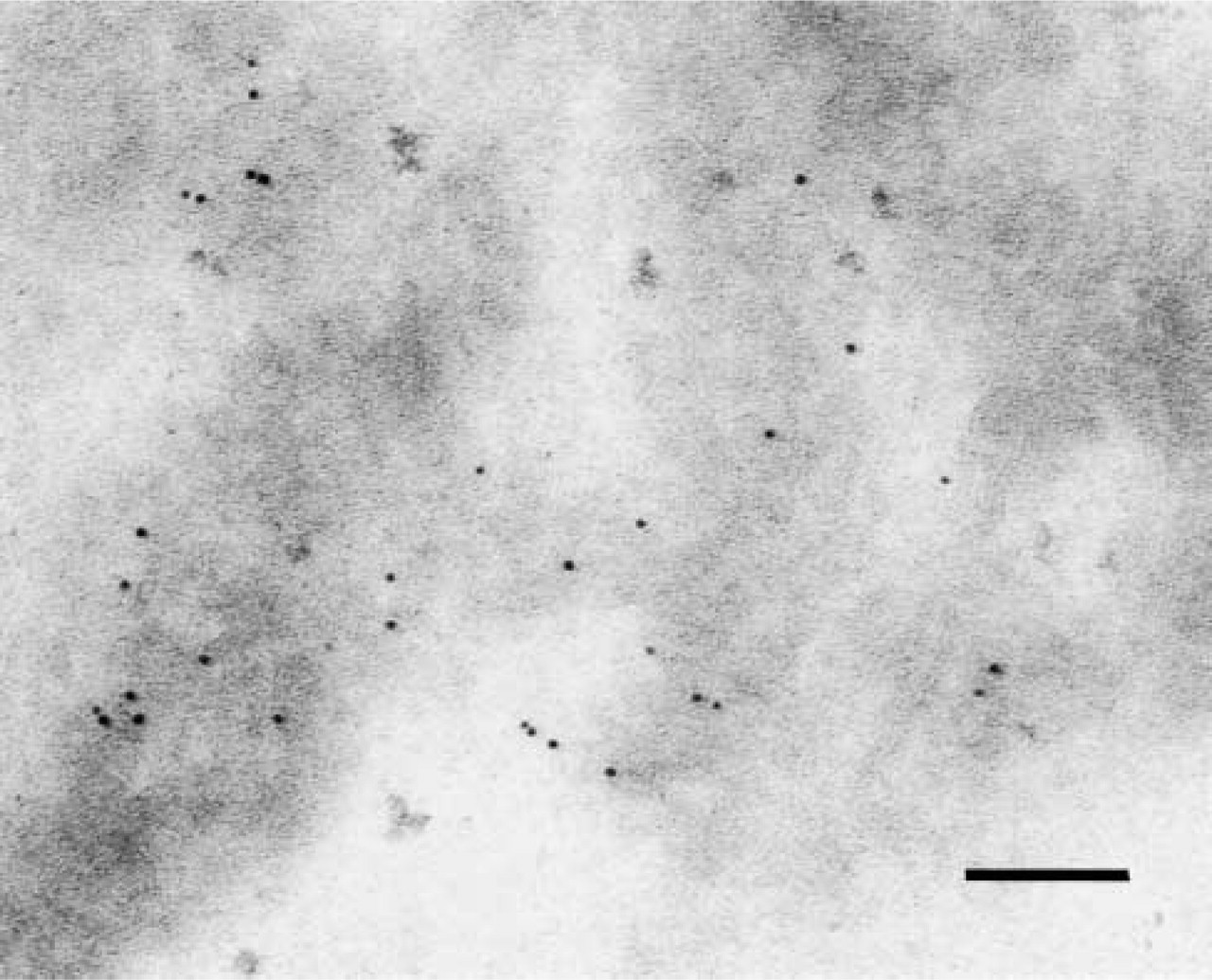
CFTR resides in ER-enriched fractions. A CFTR-positive image lacking BiP is shown. Bar = 0.1 μm.
On the basis of these data, we propose that CFTR concentrates at foci in the ER membrane but does not necessarily co-localize with BiP. This is in contrast to BiP bodies that form as a result of an ER-to-Golgi secretion block and that represent BiP-enriched, ER exit sites (Nishikawa et al. 1994). We suggest instead that CFTR resides in the yeast equivalent of Russell bodies that are ER subdomains in which BiP is absent and in which misfolded proteins accumulate (Umebayashi et al. 1997; Kopito and Sitia 2000).
Footnotes
Acknowledgements
Supported by a RDP grant to the University of Pittsburgh from the Cystic Fibrosis Foundation, by grant DK60385-01 from the National Institutes of Health, and by grant MCB-0110331 from the National Science Foundation.
We thank Y. Zhang for supplying yeast used for the initial micrographs, and E. Jones for providing anti-Pep12p antiserum.