
Abstract
NAD(P)H:quinone oxidoreductase 1 (NQO1; DT-diaphorase; DTD) is a cytosolic two-electron reductase, and compounds of the quinone family such as mitomycin C are efficiently bioactivated by this enzyme. The observation that DT-diaphorase is highly expressed in many cancerous tissues compared to normal tissues has provided us with a potentially selective target that can be exploited in the design of novel anticancer agents. Because of the relative lack of information about the cell-specific expression of DT-diaphorase, the purpose of this study was to map the distribution of this enzyme in normal human tissues. Fifteen tissue samples from normal human kidney were analyzed for expression of DT-diaphorase by immunohistochemistry (two-step indirect method). We found a specific high expression of DT-diaphorase in glomerular visceral epithelial cells (podocytes). These results suggest that a high expression of DT-diaphorase in podocytes could play a major role in the pathogenesis of renal toxicity and mitomycin C-induced hemolytic uremic syndrome, in which injury to the glomerular filtration mechanism is the primary damage, leading to a cascade of deleterious events including microangiopathic hemolytic anemia and thrombocytopenia. This observation has potential therapeutic implications because the DT-diaphorase metabolic pathway is influenced by many agents, including drugs, diet, and environmental cell factors such as pH and oxygen tension.
M
DT-diaphorase-rich cells are therefore more sensitive to MMC (Lee et al. 1993; Saeki et al. 1995; Mikami et al. 1996). A high expression of this enzyme has been demonstrated in many cancerous tissues compared to normal tissues (Cresteil and Jaiswal 1991; Malkinson et al. 1992; Belinsky and Jaiswal 1993; Smitskamp-Wilms et al. 1995). This allows the generation of high concentrations of reactive compounds specifically at the tumor site by the generation of DTD-activated prodrugs (Beall et al. 1995), decreasing the risk for toxicity to normal cells. Because reaction with DNA appears to be the primary event leading to cell death, these compounds are most toxic towards rapidly dividing cells. In standard doses, MMC-related common toxicities are nausea, vomiting, myelosuppression, and alopecia. In addition, MMC can cause renal toxicity (Hanna et al. 1981; Cantrell et al. 1985; Lesesne et al. 1989). Nephrotoxicity induced by MMC was first reported in 1971 (Liu et al. 1971). It has been described in up to 10% of patients treated with MMC, and is total-dose-dependent (Valavaara and Nordman 1985; Verwey et al. 1987).
Although there are increasing numbers of reports of MMC-associated renal toxicity, the key events that initiate or precede the glomerular injury have not yet been identified, and the reason why the renal glomerulus is apparently the major target for MMC-induced renal toxicity is still unknown.
Here we report our findings on the immunohistochemical detection of DT-diaphorase in human kidney.
Materials and Methods
Human Tissues
Archival samples of formalin-fixed, paraffin-embedded tissues were supplied by the Institute of Pathology of Southern Switzerland (Locarno, Switzerland). We analyzed 15 bioptic samples of normal human kidney taken from patients who underwent nephrectomy because of kidney carcinoma (hypernephroma). All these patients were chemotherapy-naive.
Methods
We performed an immunohistochemical analysis for detection of DTD on tissue sections using anti-DTD antibodies.
Antibodies and Reagents. Anti-DTD monoclonal antibody (IgG1)-secreting hybridomas (clones A 180 and B 771) were derived from a BALB-c mouse immunized with purified recombinant human DTD protein. All hybridoma cell lines were grown in spinner flasks in RPMI medium containing 50 U/ml penicillin, 50 μg/ml streptomycin, 1% L-glutamine, and 10% fetal bovine serum in 5% CO2 at 37C to a concentration of 106 cells/ml. Hybridoma tissue culture supernatants were prepared by centrifugation at 1800 rpm for 10 min and then stored at −80C.
Before use, supernatants were centrifuged at 14,000 rpm for 5 min. We demonstrated specificity of our proprietary antibody by preabsorbing it with antigen (human recombinant DTD) and showing that immunohistochemical staining was completely blocked. We also performed a Western analysis on tissue extracts to confirm immunohistochemical staining specificity.
Non-human reactive monoclonal mouse antibodies produced in tissue culture, subclass IgG1, were used as negative control reagent (Mouse IgG1 Negative Control Code No. X0931; DAKO, Glostrup, Denmark).
Immunohistochemical Method. Immunohistochemistry was performed on tissue sections (3 μm) cut from archival paraffin blocks. Sections were de-paraffinized in xylene and rehydrated through graded alcohol to running water, then placed in citrate buffer, pH 6.0, and microwaved for two 3-min cycles. Endogenous peroxidase activity was blocked by adding Peroxidase Blocking Agent (DAKO En Vision Kit; Carpinteria, CA). Nonspecific binding was blocked by adding 20% normal rabbit serum. Serial sections of each tissue sample were then incubated with either anti-DTD or control antibodies for 30 min at room temperature. The secondary antibody was added for 30 min (DAKOKIT, labeled Polymer HRP anti-mouse: peroxidase labelled polymer conjugated to goat anti-mouse immunoglobulins). Immunodetection was performed using a substrate-chromogen solution (DAKOKIT, hydrogen peroxide and 3,3′-diaminobenzidine chromogen). Slides were counterstained with hematoxylin, dehydrated through graded alcohols, mounted, and photographed.
Scoring of DTD Immunostaining. The intensity of immunostaining of DTD (brown staining) was visually scored as 0 (negative), +1 (very weak), +2 (weak), +3 (strong), or +4 (very intense), as previously reported for human lung cancers (Siegel et al. 1998).
Results
All samples analyzed were suitable for immunohistochemical analysis of DTD expression. There was no immunostaining in control sections when nonspecific antibodies were used.
DTD expression was absent in intertubular stroma (score 0), weak in epithelial cells of proximal convoluted tubules (score +2), strong in epithelial cells of distal convoluted tubules and cortical collecting tubules (score +3), and very strong in glomerular visceral epithelial cells (podocytes) (score +4) of all samples analyzed. Mesangial cells and glomerular parietal epithelial cells did not show any DTD staining (Figures 1A-1C).
Discussion
We report a high expression of DT-diaphorase in human glomerular visceral epithelial cells (podocytes). This implies that DTD-directed compounds, such as MMC, might be selectively bioactivated in podocytes and might thus explain the histological feature of glomerular injury that is central to the pathogenesis of MMC-induced kidney toxicity and hemolytic uremic syndrome. Podocytes are highly differentiated and quiescent cells that play a crucial role in the physiology and pathology of the kidney glomerulus. These cells, which envelop the glomerular capillaries, are an important component of the glomerular filter. The glomerular filtrate must traverse capillary endothelium, basement membrane and the podocyte layer before reaching Bowman's space. Podocytes also produce vascular endothelial growth factor (VEGF), which regulates glomerular endothelial permeability (Kretzler et al. 1998). What do we know about renal diseases secondary to podocyte injury? An acquired primary dysregulation of podocytes is described in collapsing idiopathic focal segmental glomerulosclerosis and in HIV-associated nephropathy (Barisoni et al. 1999,2000). A congenitally altered genotype and phenotype of podocytes is also known, such as in congenital nephrotic syndrome, an inherited autosomal recessive disease in which the gene mutated codes for nephrin, a cell surface protein of podocytes (Patrakka et al. 2000). Because cells with elevated DTD levels are generally more sensitive to bioreductive antitumor agents such as MMC, a high expression of DTD explains, in theory, a selective toxicity towards podocytes of DTD-bioactivated compounds. There are two clinically distinct patterns of MMC-induced renal damage (Hanna et al. 1981), an acute and often rapidly fatal glo-merular toxicity associated with microangiopathic hemolytic anemia and thrombocytopenia (cancer-associated hemolytic uremic syndrome; C-HUS), and a more chronic and progressive renal toxicity not associated with other C-HUS features. Both types show that an injury to the glomerular filter is central to the pathogenesis of renal damage (Hanna et al. 1981; Verwey et al. 1984; Jain and Seymour 1987). It is assumed that this precedes platelet activation and consumption in C-HUS. Histology shows typical renovascular damage characterized by necrosis and sclerosis of glomeruli, as well as mesangial interdeposition with double contouring of glomerular basement membranes and large atypical endothelial cell nuclei (Liu et al. 1971; Hanna et al. 1981; Verwey et al. 1984; Cantrell et al. 1985; Price et al. 1985; Sheldon and Slaughter 1986). These histological changes are similar to those observed in other glomerulopathies secondary to acquired or congenital podocyte dysfunction. It therefore seems reasonable to postulate that a high expression of DTD in human podocytes could be responsible for MMC-induced renal toxicity.
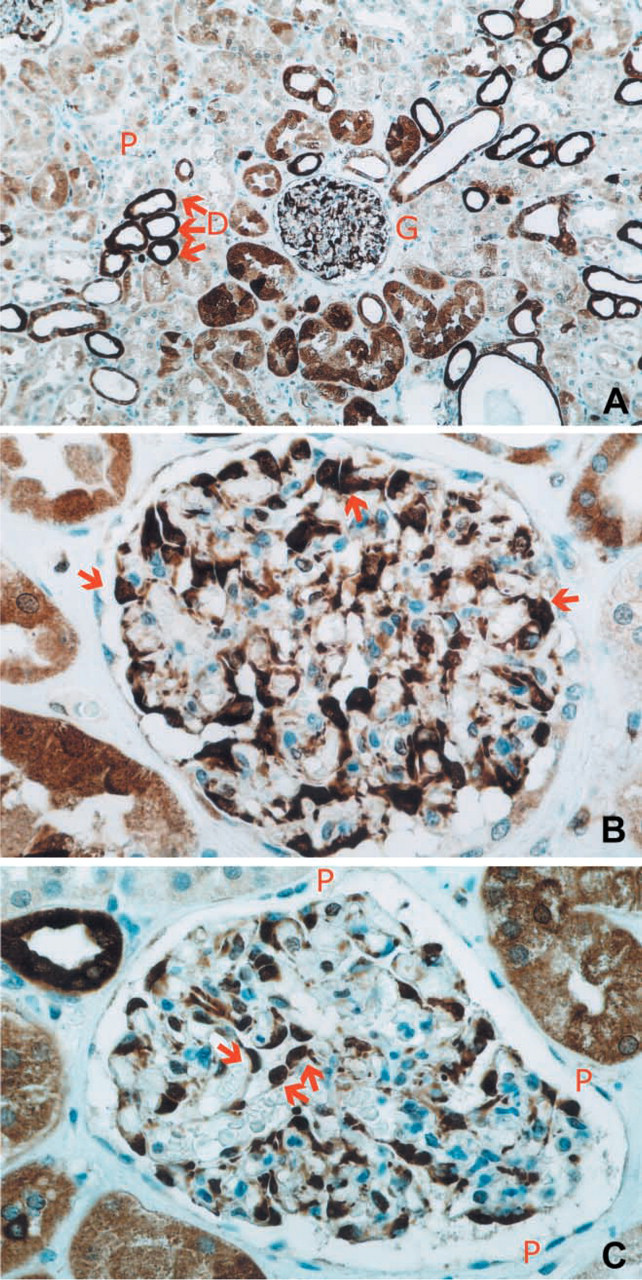
Immunoperoxidase staining (DAB) of formalin-fixed, paraffin-embedded human kidney sections using monoclonal antibodies to human DTD. Sections were counterstained with hematoxylin. (A) Weak expression of DTD in proximal convoluted tubules (recognized by a narrow lumen, P), strong expression of DTD in distal convoluted and collecting tubules (recognized by a larger lumen, D). One kidney glomerulus is visible in the middle of the section (G). (B) Higher magnification showing very strong expression of DTD in glomerular visceral epithelial cells (podocytes, brown stained, red arrows). (C) Higher magnification of podocytes (brown-stained, red arrows) covering glomerular capillaries. Glomerular parietal epithelial cells (P) do not express DTD.
If the high expression of DT-diaphorase in these cells is the key element in the pathogenesis of MMC-induced toxicity, other questions remain unanswered. Why is the risk for development of C-HUS after treatment with MMC between 4% and 15% and not higher? What is it that protects more than 85% of patients against this potentially lethal side effect? We propose the following mechanisms that might explain this individual susceptibility to MMC-related renal toxicity. (a) DT-diaphorase can be induced by a wide variety of compounds, including many dietary substances (Benson et al. 1980; O'Dwyer et al. 1996; Begleiter et al. 1997; Wang et al. 1999). For example, rapid induction of this enzyme has been demonstrated in subjects who regularly ingest coffee or broccoli (Sreerama et al. 1995). Moreover, in a previous study the major determinant of the variability of human lung DT-diaphorase was the cigarette-smoking history of the patient and the alcohol intake (Schlager and Powis 1990). Dietary or comportmental habits might increase the individual expression of DT-diaphorase enough to predispose to MMC-related toxicity. (b) Previous results indicate that DT-diaphorase activity is required for effective cytotoxicity of quinone-containing agents in human carcinoma cell lines (Lee et al. 1993; Sun and Ross 1996; Belcourt et al. 1998). However, cytogenetic heterogeneity within the tumor cell population and the various environmental factors that impact on bioreductive enzyme function, including pH and oxygen tension, may subvert this approach (Siegel et al. 1990; Begleiter et al. 1992; Begleiter and Leith 1993; Saeki et al. 1995). Individual variations in oxygen tension and pH (chronic lung diseases, metabolic or respiratory acid-basic status dysregulations) could play a role in modulating MMC cell metabolism. Kidney capillaries are located in a physiologically strategic pathway of acid-basic homeostasis, and this could influence DTD sensitivity to MMC. (c) DT-diaphorase is known to share two different functions, a bioactivating function that is exploited in antitumor activity and a more physiological phase II detoxifying function. The high expression of DT-diaphorase in podocytes could be related to a physiological detoxifying role, protecting these cells from potentially cyto-toxic or mutagen xenobiotics filtered in the glomerular urinary space from the bloodstream. A lack of this enzyme in podocytes would therefore render these cells more sensitive to the toxic effect of physiologically occurring or pharmacological bioreductive compounds. This “fragile” podocyte will be particularly sensitive to the toxic effect of DTD-directed antican-cer compounds. A polymorphism of the gene encoding DT-diaphorase has been described (Traver et al. 1997). This mutation (a C-to-T transition at base pair 609 coding for a proline to serine substitution) causes a loss of enzyme activity (Ross et al. 1996; Siegel et al. 1999) and resistance to anticancer agents that require reductive activation (Lambert et al. 1998). Individuals homozygous for this change (T/T) have no detectable DT-diaphorase activity. Previous studies indicate a significant ethnic variation in the occurrence of this point mutation (Edwards et al. 1983). A 4%-6% range of individuals homozygous for the mutation has been reported for the general population of the UK and among whites and African-Americans (Larson et al. 1999), and a higher incidence for Hispanics and Asians (Wiencke et al. 1997; Chen et al. 1999). An individual genetic variation in DT-diaphorase activity could at least partially explain the variation in susceptibility to MMC. Moreover, even if the two-electron pathway catalyzed by DT-diaphorase and unimpeded by molecular oxygen is probably the critical pathway for MMC antitumor activity, through a one-electron pathway catalyzed by NADPH-dependent cytochrome P450 reductase and impeded by molecular oxygen, semiquinone free radical intermediates are formed that may participate in MMC-induced toxicity (Bachur 1978; Lind et al. 1982; Siegel et al. 1990; Niedermeyer et al. 1999). In individuals lacking DT-diaphorase activity because they are homozygous for the point mutation, MMC metabolism preferentially follows the one-electron pathway catalyzed by the cytochrome P450 reductase, leading to minor antitumor activity but also to increased production of free radicals potentially responsible for cytotoxicity. A previous study suggests that DT-diaphorase can reduce MMC to potent cytotoxic species under aerobic conditions, and this activity predominates over the one-electron-reducing enzymes even in hypoxic conditions. In the absence of DT-diaphorase activity because of the point mutation in the NQO1 gene, MMC is reduced in hypoxic conditions by one-electron-reducing enzymes (Plumb and Workman 1994). Because of the similar range of incidence reported for individuals ho-mozygous for the NQO1 gene point mutation and for C-HUS and MMC-induced renal toxicity, the DTD-independent one-electron metabolism of MMC could be the critical pathway leading to renal toxicity, affecting only DTD-deficient individuals. Until now, no studies have focused on the possible relationship between the NQO1 gene mutation and the incidence of MMC-induced C-HUS.
We postulate that genetic, dietary, and environmental cell factors may be responsible for the individualism of susceptibility to DTD-directed anticancer agents. We also propose that MMC-induced hemo-lytic uremic syndrome and kidney toxicity could be related to the point mutation in the NQO1 gene, encoding an inactive protein. This report represents an attempt to explain the selective toxicity of quinones on kidney glomeruli, and proposes exciting new fields for further research.
Footnotes
Acknowledgements
Francesco Zappa is supported by an ESMO (European Society for Medical Oncology) grant.
Anti-DTD antibodies were a generous gift from Prof David Ross (Department of Pharmaceutical Sciences, School of Pharmacy and Cancer Center, University of Colorado Health Sciences Center; Denver, Colorado).