
Abstract
We studied the level of the basal (constitutive) HSP70 expression (inducible and constitutive forms) in the central nervous system (CNS) of male and female rats from the postnatal period to maturity. HSP70 levels were analyzed by immunoblotting in five different areas (cortex, hippocampus, hypothalamus, cerebellum, and spinal cord). The highest levels of HSP70 were found in juvenile rats and decreased progressively until reaching baseline levels between 2 and 4 months. A slight and nonsignificant increase in aged (2-year-old) rats compared with adult subjects was observed in some cerebral areas (cerebral cortex, hippocampus, and cerebellum). In the first weeks of postnatal development, HSP70 immunoreactivity was distributed throughout CNS sections and no specific immunopositive cells could be clearly determined. In adult animals, strong immunostaining was observed in some large neurons (Purkinje neurons and mesencephalic and spinal cord motor neurons), some perivascular and subpial astrocytes, and ependymocytes. Immunoelectron microscopy revealed that HSP70 in these cells is located in the perinuclear area and in mitochondria, rough endoplasmic reticulum, and microtubules. In neurons, strong immunolabeling was also observed in synaptic membranes. The postnatal time course of HSP70 levels and the location and size of HSP70-immunopositive cells suggest that HSP70 constitutively expressed in the rat CNS may be mainly determined by the degree of development and metabolic activity of the neural cells.
C
The HSPs are usually classified according to their molecular weight. Among the 11 families described, attention has focused on the family of 70-kD heat shock protein (HSP70) because it is prominently induced in a variety of models of cell stress and is also the best conserved stress protein. HSP70 is a multigene family composed of constitutively expressed members (Hsc70) and stress-inducible members (Hsp70), and both can be basally expressed in the nervous system under control conditions (Manzerra et al. 1994; Foster and Brown 1996). We refer to the constitutive form of HSP70 as Hsc70 and the inducible form as Hsp70, and the general term of HSP70 implies both Hsc70 and Hsp70.
HSP70 binds and subsequently releases partially unfolded proteins in an ATP-dependent manner. More specifically, members of the HSP70 family protect nascent protein chains after synthesis, translocate proteins across membranes, and refold proteins after denaturation or during protein degradation (see reviews by Feige and Polla 1994; Kiang and Tsokos 1998; Mayer and Bukau 1998).
Expression of HSP70 proteins has been demonstrated in the central nervous system (CNS). The neural tissue has a high HSP70 constitutive expression compared with other organs and tissues (Tanguay et al. 1993). HSP70 expression is enhanced by different stress conditions, and the stress response differs in different CNS regions (Brown 1990; Manzerra and Brown 1992). HSP70 is expressed by both neurons and glial cells (Brown et al. 1989; Nowak et al. 1990). Astrocytes have a different (Nowak et al. 1990) and more vigorous (Nishimura et al. 1991; Satoh and Kim 1994; Marcuccilli et al. 1996) response to stress than neurons, although contradictory results have been reported (Pardue et al. 1992; Li et al. 1995). HSP70 expression in neural tissue appears to be cell-specific and to depend on the stressor (Krueger et al. 1999). Moreover, HSP70 expression may correlate with the constitutive location in CNS of heat shock transcription factors (HSFs) (Stacchiotti et al. 1999).
Most of the studies in the literature have analyzed the response to stress, with little attention to HSP expression under unstressed conditions. The aim of the present study was to examine the constitutive (basal) expression of HSP70 in the normal rat CNS from the postnatal period to maturity. Because the inducible form of HSP70 can also be constitutively expressed in the normal CNS (see above), we have used an anti-HSP70 antibody that recognizes both constitutive and inducible forms. Moreover, the immunohistochemical (IHC) study was made to identify the type of neural cells involved in HSP70 expression and the immuno-electron microscopic study was also made to localize HSP70 at the subcellular level.
Materials and Methods
Male and female Wistar rats (Rattus norvegicus L.) were analyzed at different postnatal ages (7, 14, and 21 days, 1, 2, 4, and 8 months, 1 and 2 years). They were maintained on a 12-hr dark/light cycle, housed at 20 ± 1C, 50% relative humidity, and fed and watered ad libitum. Animal care procedures were conducted according to the directive 86/609/EEC adopted by the Council of the European Communities for the protection of animals used for experimental and other scientific purposes.
Immunoblotting
Rats (four male and four female rats per time point) were anesthetized, brain and spinal cord removed, placed in PBS 0.1 M, pH 7.4, and frozen at −70C for subsequent analysis. Pieces were homogenized for 30 sec at 20,000 rpm (Ultra-Turrax T25; IKA Labortechnik, Staufen, Germany), sonicated (three times for 10 sec, setting 18) (Microson XL; Mix onix, Farmingdale, NY) at 4C in 1 ml PBS, and centrifuged at 18,000 × g for 10 min at 4C (Avanti J25; Beckman, Palo Alto, CA). The pellets were discarded and a sample (200 μl) of supernatant was analyzed to determine protein concentration (Bradford method).
Supernatant samples (7 μg) were prepared for electrophoresis on polyacrylamide slab gels transforming their buffer in a solubilization buffer (0.06 M Tris, 5% β-mercaptoethanol, 2% SDS, 10% glycerol, pH 6.8). The amount of protein per sample was obtained from the linear ranges of standard curves. Stacking gels contained 4% and separating gels contained 10% acrylamide. After electrophoresis proteins were transferred to nitrocellulose (HSI; San Francisco, CA). The nitrocellulose sheets were cut into strips, washed in Tris buffer (0.05 M, pH 7.6) for 5 min, and incubated with agitation for 15 min at room temperature (RT) in Tris buffer with 5% (w/v) nonfat powdered milk (Blotto buffer). Without washing, membrane strips were incubated overnight at 4C with a monoclonal primary antiserum against HSP70 (clone BRM-22; Sigma, St Louis, MO) diluted 1:5000 in Tris buffer. This antibody recognizes both constitutive and inducible HSP70 forms. After three 5-min washes with agitation (once in Blotto buffer and twice in Tris buffer), strips were incubated in peroxidase-conjugated goat anti-mouse IgG (Sigma) diluted 1:600 for 1 hr at RT. Strips were then rinsed once in Blotto buffer and twice in Tris buffer. The peroxidase reaction was carried out in a solution of 0.05 M Tris buffer containing 0.1% 3,3-diaminobenzidine (Sigma) and 0.05% H2O2. Control strips were used to test antibody specificity, one with plasma proteins and another in the absence of primary serum. These control strips showed no immunoreactive product. A videodensitometric procedure (Microm; Barcelona, Spain) was used for quantification of immunoblotting immunoreactivity. Data are expressed as the mean ± SEM (n = 4, in duplicate).
Immunohistochemistry
Rats (three animals per time point) were transcardially perfused with 3% paraformaldehyde in PBS and postfixed for 2 hr (spinal cord) and 4 hr (brain). After fixation the pieces were washed in PBS, dehydrated in ethanol, and paraffin-embedded. Transvere section slices 9 μm thick were obtained. Dewaxed sections were incubated for 30 min in goat normal serum at 1:30 dilution. Without washing, sections were incubated for 1 hr at RT and overnight at 4C with a primary anti-HSP70 monoclonal antibody (MAb) diluted 1:2000 in Tris buffer. After three 10-min washes in Tris buffer, the sections were incubated at RT for 1 hr in peroxidase-conjugated goat anti-mouse immunoglobulin (Sigma) at 1:200 dilution and rinsed twice in Tris buffer. Peroxidase activity was revealed using 0.1% 3,3-diaminobenzidine in Tris-buffer with 0.05% H2O2 for 10 min. Sections then were washed with distilled water, dehydrated in graded series of ethanol, and mounted in DePeX. Control sections were incubated with normal serum instead of primary antibody and showed no immunoreactive product.
Immunoelectron Microscopy
Four-month-old rats (two males and two females) were transcardially perfused with 3% paraformaldehyde and 0.5% glutaraldehyde in PBS and postfixed for 2 hr (spinal cord) or 4 hr (brain). After fixation, the pieces were washed for 2 days in PBS at 4C and 150-μm sections were obtained in a vibratome (Micro-Cut H1200; Biorad, Hertfordshire, UK). Free-floating sections were washed in PBS (twice for 5 min), incubated with normal goat serum diluted 1:30 in PBS for 30 min and then, without washing, with anti-HSP70 MAb diluted 1:1000 in PBS with 0.1% Triton X-100 for 1 hr at RT and overnight at 4C. Then the sections were washed in PBS (three times for 10 min) and incubated at 20C for 1 hr in peroxidase-conjugated goat anti-mouse antibody diluted 1:100 in PBS and rinsed in PBS (three times for 10 min). The peroxidase reaction was carried out as described above. Sections were then rinsed in PBS (twice for 5 min) and postfixed for 10 min in 1% OsO4 in PBS. Finally, sections were dehydrated in graded concentrations of acetone and embedded in Araldite. Small fragments of those sections were dissected and mounted on Araldite blocks to obtain ultrathin sections. These sections were stained with uranyl acetate and lead citrate.
Statistical Analysis
Data on immunoreactivity obtained from the immunoblot stripes were statistically evaluated using the Statgraphics program (Microsoft). Graphics for HSP70 immunoreactivity were done using mean ± SEM for every age and CNS region. The 95% LSD method, a multiple-range test, was used to determine the basal level of HSP70 expression. This method applies a multiple comparison procedure to determine which means are significantly different from others and, in our study, serves to form homogeneous groups (considering the mean) in every studied area of the CNS. The 95% CI for mean of the homogeneous group with lowest HSP70 expression was used to define the range of basal expression in every CNS area.
Results
The postnatal time course of the level of HSP70 constitutively expressed (basal expression level) in different cerebral areas and the spinal cord of normal rats, analyzed by Western blotting, is shown in Figures 1 and 2. Because no sexual dimorphism was observed in most areas analyzed, data from male and female rats were pooled. However, the hypothalamus showed sexual differences in HSP70 level, and therefore data from males and females were considered separately. Using a multiple-range test, it was possible to make a homogeneous group with the lowest HSP70 levels in each CNS area. We took the 95% CI for the mean of this group as the range of baseline HSP70 level. Baseline HSP70 level was reached in cerebral cortex, cerebellum, and hippocampus between 2 and 4 months, in the spinal cord between 1 and 2 months, and in the hypothalamus between 1 and 2 months for females and between 4 and 8 months for males. The baseline level of HSP70 constitutively expressed was similar in all the areas studied except in female hypothalamus and spinal cord, which had a higher baseline HSP70 level. The highest constitutive HSP70 level was consistently found at 7 days, except in the male hypothalamus, where it was reached at 21 days. No significant differences were observed between adult and aged rats, although a slight increase in aged (2-year-old) animals compared with adult animals was observed in some cerebral areas (cerebral cortex, hippocampus, and cerebellum).
During the first 3 weeks of postnatal development, HSP70 immunoreactivity was distributed throughout all CNS sections analyzed and no specific cells could be clearly determined. Later, from the age of 2 months on, a general loss of immunoreactivity was observed (Figures 3I and 3J). However, some specific types of neural cells showed strong immunoreactivity: Purkinje neurons (Figure 3A), motor neurons in the spinal cord (Figure 3B) and mesencephalon (Figure 3C), some astrocytes, especially subpial or perivascular astrocytes (Figures 3D and 3E), and ependymocytes (Figures 3F and 3G). Some glial cells in the white matter and the axons of small myelinated fibers also showed intense immunostaining. Some glial cells had strong nuclear immunoreactivity, particularly in 2-month-old animals (Figure 3H).
Neurons and astrocytes (Figure 4A) were the most prominently immunostained cells, with immunoreactivity usually distributed throughout the cell cytoplasm, sometimes forming a perinuclear rim. Ependymocytes also showed strong immunostaining, whereas oligodendrocytes and endothelial cells (Figure 4B) were poorly immunolabeled. No immunostaining was detected in microglial cells. In both neurons and astrocytes under electron microscopy, peroxidase reaction products were especially abundant on the external surface of mitochondria (Figures 4D and 4E), rough endoplasmic reticulum (Figure 4B), microtubules (Figure 4C), external membrane of the nucleus (Figures 4A), and dispersed throughout the cytosol. Immunoprecipitation also was found in mitochondria and microtubules of some axons (Figure 4C) and synapses (Figures 4D and 4E).
Discussion
Postnatal Time Course of HSP70 Immunoreactivity
The present results demonstrate that the level of constitutively expressed HSP70 (basal expression level) changes during the postnatal development of the normal rat CNS. Because the antibody used recognized both constitutive (Hsc70) and inducible (Hsp70) forms of HSP70, our data on HSP70 immunoreactivity level are always the sum of Hsc70 plus Hsp70, although in the CNS the constitutive expression of Hsp70 is very low compared with the constitutive expression of Hsc70 (Manzerra et al. 1994; Foster and Brown 1996; D'Souza and Brown 1998; Suzuki et al. 1999).
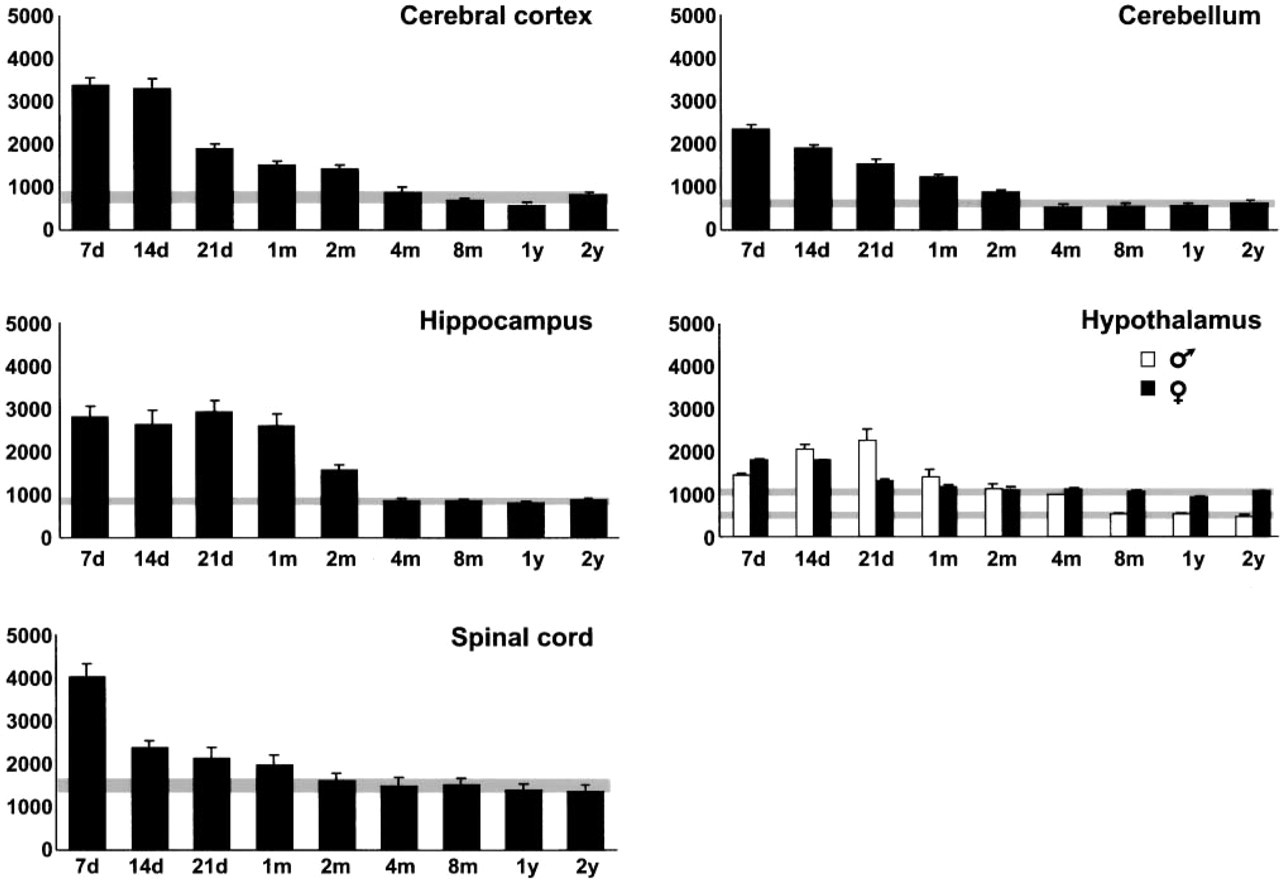
Graphics of the postnatal time course of the level of constitutively expressed HSP70. HSP70 immunoreactivity (arbitrary units) is represented in the Y-axis and time (d, days; m, months; y, years) is represented in the X-axis. The gray stripe represents baseline HSP70 level and is the 95% CI for mean (considering the homogeneous group with lower HSP70 level established using a multiple-range test). Bars represent mean ± SE; n=8 (four males and four females). Female (n=4) and male (n=4) rats were considered separately in the hypothalamus.
The level of constitutively expressed HSP70 was high in the first month of postnatal development, especially at the first week, and decreased slowly to reach baseline levels in adult animals. Abundant HSP70 protein has also been demonstrated throughout postnatal neural development (D'Souza and Brown 1998) and in late chicken embryos (Voellmy and Bromley 1982), and a pronounced decrease in Hsc70 has been described during postnatal development of the kidney (D'Souza and Brown 1998). Lower amounts of Hsc73 have been found in the brains of adult mice than in young (11-day-old) mice (Tanguay et al. 1993). However, the basal level of constitutively expressed HSP70 in 2-year-old rats was significantly higher in some parts of the brain (pons, medulla, striatum, and thalamus) than that in 6-month-old rats, suggesting that HSP70 plays a role in suppressing protein denaturation with aging (Unno et al. 2000). Our study demonstrated a lower level of HSP70 constituvely expressed in both adult and aged rats than in young rats. Although no significative differences between adult and aged rats have been observed, a slight increase in aged animals was observed in cerebral areas such as cerebral cortex, hippocampus, and cerebellum. In support of our results, similar results were obtained by Unno et al. (2000) in the same cerebral areas, and the basal HSP70 mRNA is increased in the kidney of young and old rats compared with adult animals (Maiello et al. 1998), suggesting that the prevalence of anabolic activity can trigger the increased basal production of HSP70 in young animals and that the accumulation of post-translationally modified proteins can enhance HSP70 synthesis in the kidney of aged animals.
Using the 95% LSD statistical method, we defined the baseline level of HSP70 constitutively expressed for each CNS region studied. The baseline HSP70 level in different areas of rat CNS observed in our study was comparable to that found in adult rabbit brain (Foster and Brown 1996). Baseline HSP70 level was reached between 2 and 4 months of postnatal development in the cerebral cortex, cerebellum, and hippocampus, and it was reached earlier, between 1 and 2 months, in the spinal cord. It is usually accepted that rats reach maturity at 3 months. The growth rate slows at this age, and growth consists mainly of adipose tissue accumulation. The correlation coefficient r between the body growth rate of Wistar rats (data obtained in the animal house of the Alcalá de Henares University) and HSP70 level in the CNS (mean of five CNS areas for each age) was 0.954. This figure indicates a high correlation between the growth rate of body weight and the level of HSP70 constitutively expressed in the CNS, since the growth rate of body weight decreases, although body weight itself increases with age.

Immunoblot of male cerebral cortex (d, days; m, months; y, years). Note the specificity of antibody and the progressive decrease of the immunostaining with age.
Considering that (a) normal rat CNS reaches the baseline level of HSP70 constitutively expressed between 2 and 4 months of postnatal development, and (b) the close correlation between the growth rate of body weight and CNS HSP70 level and (c) the function of HSP70 in protein synthesis, it is possible that the constitutive expression of HSP70 is determined mainly by the developmental growth of rat CNS under control (unstressed) conditions. HSP70 has been described as having a causal role in these developmental processes (Angelier et al. 1996; D'Souza and Brown 1998), but the high HSP70 level in the first weeks of postnatal development may indicate that this enhanced level of HSP70 is a developmental effect rather than a cause.
The postnatal time course of HSP70 level in different CNS areas varies with age. Cerebral cortex, cerebellum, hippocampus, and spinal cord showed similar patterns of HSP70 content, whereas the male hypothalamus showed a different pattern. The baseline level of constitutively expressed HSP70 was higher in the female than in the male hypothalamus. This could be attributable to the effect of sex hormones because males and females differ in the regulation of HSP70 expression in the hypothalamus (Olazábal et al. 1992) and because increased hypothalamic HSP70 expression has been demonstrated after estradiol treatment (Mydlarsky et al. 1995).
Different studies have analyzed the possible correlation between developmental patterns of HSP and HSF levels in recent years (Loones et al. 1997; Rallu et al. 1997; Brown and Rush 1999). Our findings in cerebellum confirm those of Brown and Rush (1999) for HSF2 expression, demonstrating a decline in HSF2 levels during postnatal cerebellar development. On the other hand, it has been demonstrated that the induction of HSP70 expression by a variety of stressors declines with age because of a decrease in HSF levels (Heydari et al. 1994; Gutsmann-Conrad et al. 1998) or disturbances in the activation of HSF1 (Heydari et al. 2000).
Currie et al. (1983) did not detect any increase in HSP70 synthesis in brain after hyperthermia until rats were more than 2 weeks old. Moreover, tissues having a high constitutive expression of HSP70 show less HSP70 induction after stress (Yu et al. 1994; Brown 1995). Because the level of HSP70 constitutively expressed in the rat CNS varies with age, it is clear that the age of the animal should be considered in studies of HSP70 induction by stress.
HSP70-immunopositive Cells
We expected that the IHC methods used to analyze HSP70 distribution would immunostain all cells in a section because HSP70 is a ubiquitous protein (for HSP70 functions see reviews by Kiang and Tsokos 1998 and Nover and Scharf 1997). Accordingly, we observed a generalized immunostaining of sections (cells and neuropil) in the first weeks of postnatal development. However, in adult animals only certain cells showed intense immunostaining, such as motor neurons, subpial astrocytes, and ependymocytes. A similar IHC pattern has been demonstrated for heat shock transcription factors (HSFs) in the rat spinal cord (Stacchiotti et al. 1999).
In vivo studies have demonstrated that constitutive expression of HSP70 is mainly neuronal and is particularly located in large neurons (Foster and Brown 1996; Stacchiotti et al. 1997). This has been correlated with a higher level of demand of proteins for structural and metabolic functions in large neurons than in small neurons (Stacchiotti et al. 1997). A similar explanation can be suggested for the intense constitutive HSP70 immunoreactivity shown by some ependymocytes and astrocytes in adult animals in this study. Intensely HSP70-immunostained astrocytes are usually located in subpial or perivascular areas, where they have special functions, such as metabolic interchange with blood vessels (Coles 1995) and/or basement membrane secretion (Bernstein et al. 1985). Strong HSP70 immunoreactivity has also been detected in perivascular astrocytes in the hippocampus of lead-exposed rats (Selvín-Testa et al. 1997). Ependymocytes also can have an active metabolism in mature brain because they act as a metabolic barrier between the brain and cerebrospinal fluid, as well as in secretory processes (Del Bigio 1995; Bruni 1998). The very low or null HSP70 immunostaining of oligodendrocytes, endothelial cells, and microglial cells observed in this study coincides with the results of other studies (Stacchiotti et al. 1997).
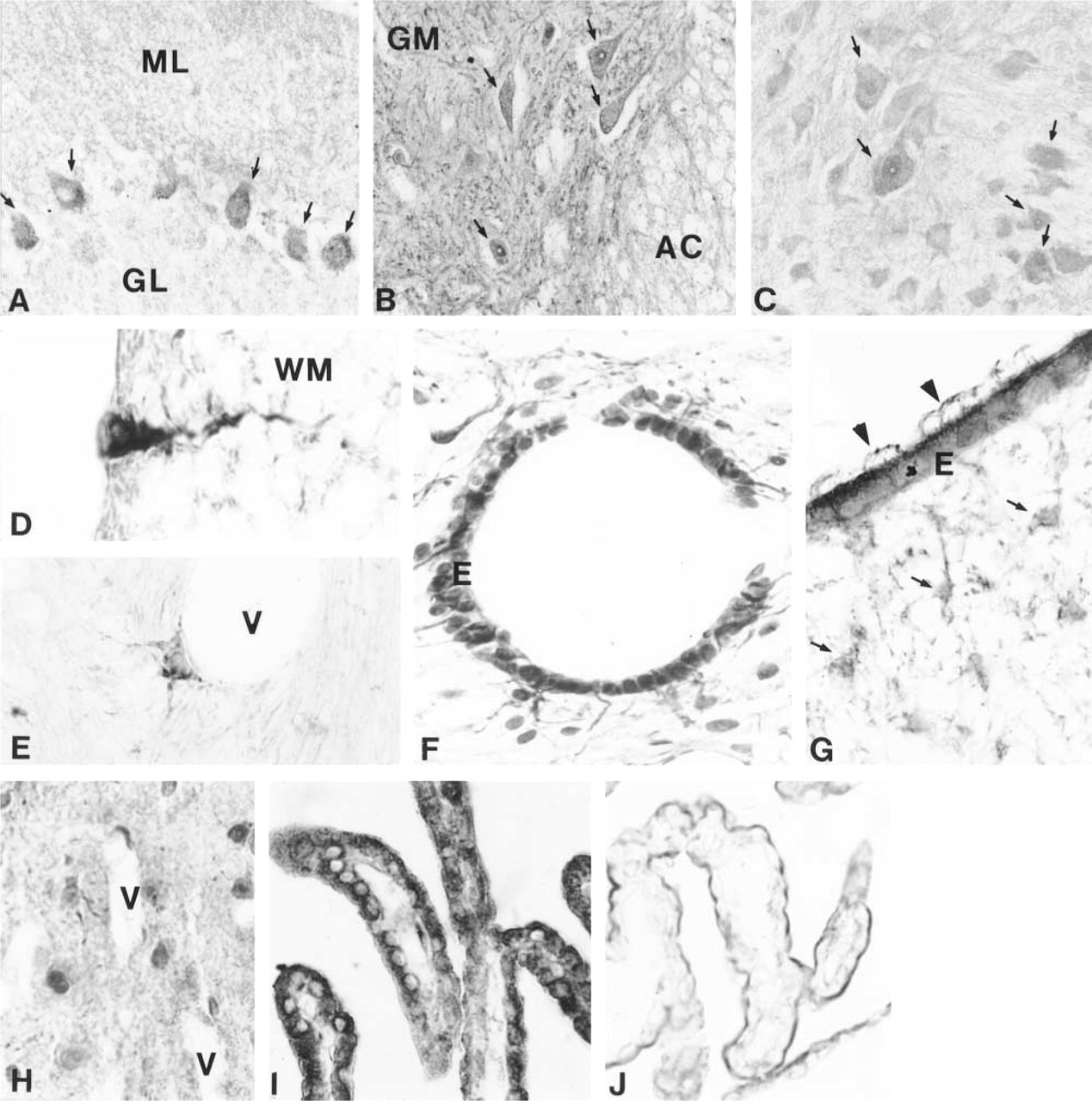
HSP70 immunohistochemistry.
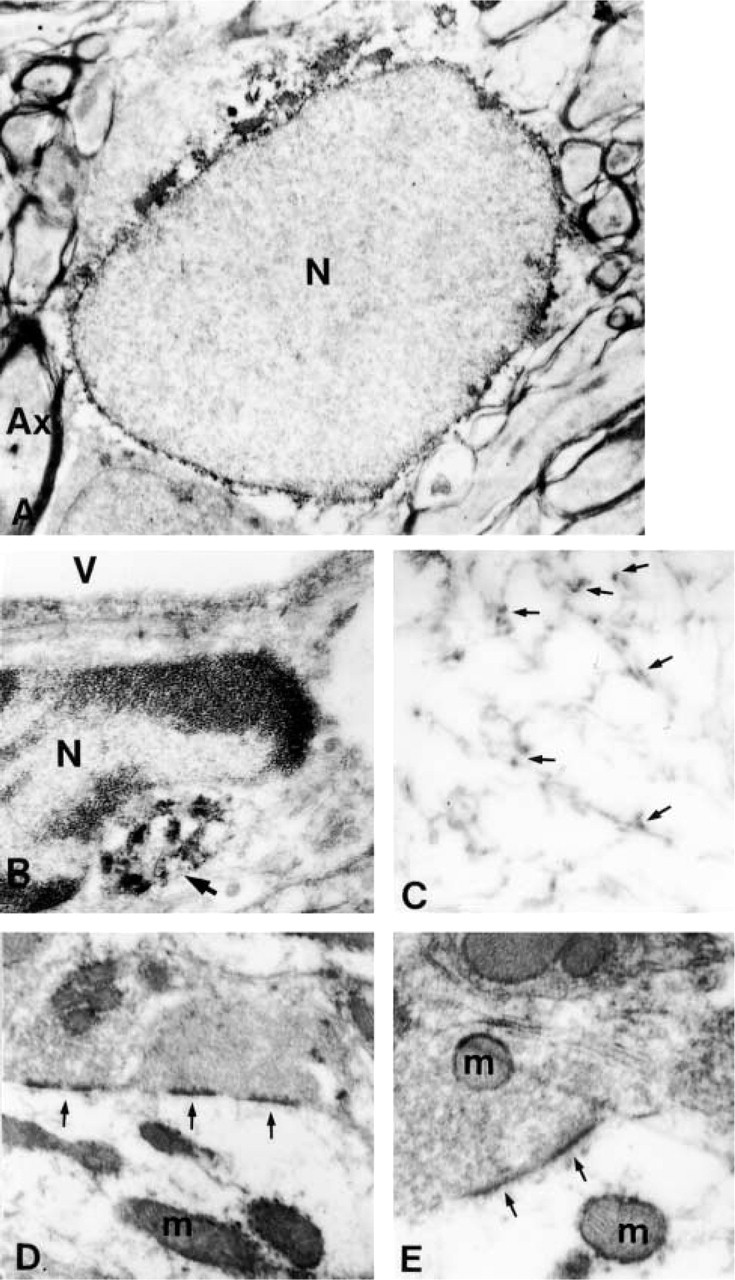
HSP70 immunoelectron microscopy; all micrographs were obtained from 4-month-old animals.
HSP70-immunopositive Subcellular Structures
Our electron microscopic study demonstrated that HSP70 immunoreactivity is located mainly in the perinuclear region and associated with microtubules, mitochondria, rough endoplasmic reticulum, and synapses. The association of HSP70 with microtubules has been demonstrated by biochemical methods (Clark and Brown 1987; Williams and Nelsen 1997). Indeed, HSP70 appears to play an important role in tubulin nucleation and polymerization (Marchesi and Ngo 1993; Williams and Nelsen 1997).
HSP70 acts as a molecular chaperone, assisting in protein synthesis and protein translocation across membranes. This function could explain its presence in mitochondria, rough endoplasmic reticulum, and the nuclear periphery. Perinuclear accumulation has been described previously and associated with cytoplasmic ribosomes (Welch and Suhan 1986). Moreover, Hsc70 is involved in the nuclear transport of karyophilic proteins (Okuno et al. 1993).
The synaptic location of HSP70 has been demonstrated in the brain under control conditions (Suzuki et al. 1999) and after hyperthermic stress (Bechtold et al. 2000). It has been suggested that, under control conditions, HSP70 could be involved in postsynaptic mechanisms of local protein synthesis related to synaptic plasticity and/or could act as a messenger from synapse to nucleus (Suzuki et al. 1999). Its presence in synapses suggests a reparative and/or protective effect related to synaptic proteins as has been postulated under stress conditions (Bechtold et al. 2000).
Footnotes
Acknowledgments
Supported by in part DGICYT grant PB94-0366-C02-01.
Thanks to Angel Blasco (Departamento de Matemáticas) and Antonio Priego (Servicio de Microscopía Electrónica) for their help in the statistical analysis and electron microscopy. We also thank Dr Julio Pérez for discussion and critical comments.