
Abstract
We investigated by transmission electron microscopy the cellular route in tumor MCF7 cells of DNA labeled with digoxigenin, carried by cationic liposomes (Lip+) prepared from TMAEC-Chol [3β(N-(N',N',N'-trimethylaminoethane)-carbamoyl)cholesterol iodide] and TEAPC-Chol [3β(N-(N',N',N'-triethylaminopropane)-carbamoyl)cholesterol iodide], two cholesterol-based cationic lipids containing a quaternary ammonium. In a previous work we showed the pathway of cationic lipid/plasmid complexes from the beginning of endocytosis until their entry into the perinuclear area. Beyond this limit, unlabeled exogenous plasmids cannot be distinguished with nuclear DNA. This work dealt with the cellular fate of cationic liposome-vectorized plasmids labeled with digoxigenin using an immunogold procedure. Early after the beginning of transfection (30 min, 1 hr, 5 hr), gold particles were observed only in the cytoplasm and in endosome-like vesicles, whereas after 24 hr gold particles were densely present in the nucleus. These results demonstrate the nuclear localization of plasmids vectorized by the cationic liposomes used. The results are discussed in comparison with transfection efficiency measurements.
Non-viral carriers for gene transfer, especially cationic lipids, are more and more often investigated (Zabner 1997; Schatzlein 2001). It was shown that DNA can be efficiently transferred by cationic lipids and cationic liposomes in a variety of cell lines in culture. Attempts for transfection in vivo were successful, and cationic lipid-mediated transfection has shown promise for vaccination and gene therapy clinical trials (Behr et al. 1989; Gao and Huang 1991; Felgner et al. 1994; Nabel et al. 1994; Ruysschaert et al. 1994; Schwartz et al. 1995; Lewis et al. 1996; Vigneron et al. 1996; Gregoriadis et al. 1997; Liu et al. 1997; Okayama et al. 1997; Fife et al. 1998). Although the ultimate goal was to have more efficient lipids and to find the best conditions for their use, investigations devoted to understanding the parameters from which the efficiency can result attract workers.
There are an increasing number of works to study the cellular pathway of DNA/lipid complexes (Zhou and Huang 1994; Tarrason et al. 1995; Wrobel and Collins 1995; Zabner et al. 1995; Friend et al. 1996; Lappalainen et al. 1997; McLean et al. 1997; Escriou et al. 1998; El Ouahabi et al. 1999). Although several experimental proofs were reported, there are still questions about the cellular fate of DNA vectorized by cationic lipids, calling for investigations to elucidate the pathways of DNA/lipid complexes after their internalization in cells and to understand the expression observed. For passage of these complexes across the cell membrane, most reports favor endocytosis (Friend et al. 1996; Lappalainen et al. 1997; El Ouahabi et al. 1999; Cao et al. 2000). However, after internalization in cells, there are divergences between the reports about the route of DNA. Investigating the pathway of oligonucleotides carried by the monocationic liposomes DDAB/DOPE (dimethyldioctadecylamonium bromide/dioleoylphosphatidylethanolamine) and the polycationic liposomes DOSPA/DOPE (2,3-dioleyloxy-N-(2(spermine carboxamido)ethyl)-N,N-dimethyl-1-propanaminium trifluoroacetate), Lappalainen et al. (1997) have observed microscopic patterns showing different routes for oligonucleotides depending on whether DDAB or DOSPA was used. For plasmids carried by DOTMA/DOPE (N-(1-(2,3-dioleyloxy)propyl)-N,N,N-trimethylammonium chloride), Friend et al. (1996) described vesicular and reticular intranuclear membranes probably resulting from the fusion of liposomes with the nuclear envelope. Here also, it appears that the cellular pathway of DNA/lipid complexes depends on the nature of the lipid used and on the cells to be transfected. In this paper the presence of DNA in the nucleus was not mentioned, whereas with various techniques plasmids were observed only in the perinuclear area (Zabner et al. 1995; Escriou et al. 1998; El Ouahabi et al. 1999). This important question remained obscure because for most studies using reporter genes transferred into cells by cationic liposomes, the expression was generally observed. It is clear that in the nucleus containing chromatin, exogenous DNA must be labeled to be distinguished. For long circular plasmids, the labeling is known but is still rarely used. Recently, plasmid DNA can be marked with fluorochrome for detection by fluorescence microscopy (Neves et al. 2000) or with BrdU (bromodeoxyuridine), allowing its immunogold detection at the ultrastructural level by electron microscopy (El Ouahabi et al. 1999).
In this work we used liposomes prepared from two cholesterol-based cationic lipids, TEAPC-Chol [3β(N-(N',N',N'-triethylaminopropane)-carbamoyl)cholesterol iodide] and TMAEC-Chol [3β(N-(N',N',N'-trimethylaminoethane)-carbamoyl)cholesterol iodide]. The efficiency of these liposomes has been shown in a previous work (Cao et al. 2000), in which we reported that liposomes interact with DNA, the complexes penetrate into cells, and functional plasmids are delivered. With transmission electron microscopy (TEM), we observed the penetration of complexes into cells by endocytosis and their internalization in endosomes, followed by their release in the cytosol and their presence in the perinuclear area. However, the observation of unlabeled plasmids in nuclei was not possible. The aim of this work is to report the delivery into the cell nuclei of functional plasmids observed by immunogold detection of digoxigenin-labeled plasmids and attested to by expression of the transgene. Up to now, digoxigenin has been used only for labeling of oligonucleotides (Tarrason et al. 1995; Friend et al. 1996).
Materials and Methods
Materials
Dioleoyl phosphatidylethanolamine (DOPE) was furnished by Aventi-Polar Lipid and was used without other purification. Reagents were analytical grade and were purchased from Aldrich-Sigma (St Louis, MO), Carlo Erba (Milano, Italia), and Fluka (Buchs, Switzerland). The digoxigenin DNA labeling kit was purchased from Roche Diagnostics (Meylan, France). Ultrasmall gold-labeled anti-digoxigenin sheep antibody Fab fragments, acetylated BSA (bovine serum albumin) (BSA-C), and the silver enhancement reagent were from Aurion (Wageningen, The Netherlands).
Cationic Lipids
The cationic lipids TMAEC-Chol and TEAPC-Chol used for this work were synthesized and characterized by Fourier transform infrared spectroscopy. The synthesis of TMAEC-Chol has been previously described (Geromel et al. 2001). TEAPC-Chol differs from TMAEC-Chol by three ethyl groups instead of three methyl groups in the polar head.
Liposome Preparation
The cationic lipids TMAEC-Chol or TEAPC-Chol and DOPE (1:1) were dissolved and mixed in chloroform and the solution was dried in a rotating evaporator. Solvent trace was removed under vacuum overnight. The following day, sterile water (Millipore; Bedford, MA) was added and the mixture was sonicated for 1 hr in cycles of 15 min (13 min followed by 2 min rest) to clarity. The liposome preparations were controlled by negative staining electron microscopy and their size was characterized by quasi-elastic light scattering (QLS) using an apparatus constructed by our laboratory. Polystyrene latex spheres of 109 nm in aqueous solution were used for the calibration.
Plasmid
The plasmid used for this work was a plasmid of 7.164 kb including the β-galactosidase reporter gene sequence under the control of the promoter of cytomegalovirus CMV (pCMV-β), furnished by Promega (Madison, WI) and propagated according to the standard method using a maxiprep kit purchased from Qiagen France (Courtaboeuf, France) and conditioned in Tris—EDTA buffer, pH 8.
Preparation of Digoxigenin-labeled Plasmids
In a sterile vial, 0.4 μl of reactive digoxigenin (DIG-Chem-Link from Roche, Meylan, France) was added to 2 μg of pCMV-β plasmid in a final volume of 20 μl. After incubation for 30 min at 50C in a waterbath, the reaction was stopped by adding 5 μl of the stop solution and the solution was stored at −20C before use.
Formation of Liposome—Plasmid Complexes
Plasmids (1 μg) and liposomes with known concentrations were separately diluted in sterile water to have 10 μl each, and then mixed. The plasmid—liposome complexes formed instantaneously and were used within 30 min for transfection.
Gel Electrophoresis
Control of Digoxigenin-labeled Plasmids. One μg of unmodified or modified plasmid was loaded on an agarose gel (1%) containing ethidium bromide in 1 X TBE (Tris buffered EDTA) buffer and analyzed electrophoretically under an electric voltage of 100 V (about 400 mA).
Retardation Control of DNA—Liposome Complex Formation. Formation of complexes between plasmid DNA and cationic liposomes results in a retardation of DNA movement and can be monitored by a gel retardation control prepared at various cationic lipid/DNA molar charge ratios. After 15 min of incubation at room temperature (RT), aliquots of complexes were mixed with the loading buffer (Blue 6X loading dye; Promega) and analyzed electrophoretically under the same conditions.
Cell Culture
Established adherent MCF7 cells (human breast carcinoma cells) were grown on plastic ware at 37C in a humid atmosphere containing 5% CO2 in air. The culture medium DMEM was supplemented with fetal calf serum (10%), penicillin/streptomycin (50 U/ml), and glutamine (2 mM).
Observation of Labeled Plasmids by Immunogold Electron Microscopy
Cells dissociated from the wells of the plate were fixed at 4C for 20 min with 1% paraformaldehyde and 0.1% glutaral-dehyde in 0.1 M phosphate buffer, pH 7.4 (PB). After washing the cells were treated for 10 min with 0.05% NaBH4 and 0.1% glycine in PB, to block the free aldehyde groups and to permeabilize the cells. The cells were then rinsed in PB and pre-incubated in 0.2% acetylated BSA (BSA-C) in PB for 30 min. They were then incubated for 2 hr at RT with diluted ultrasmall gold-labeled anti-digoxigenin antibody Fab fragments in 0.2% BSA-C in PB. The cells were rinsed in 0.2% BSA-C in PB, then in PB, and refixed with 2.5% glutaraldehyde for 10 min. After washing in PB, the samples were postfixed in 0.5% OsO4 in PB for 20 min, washed in de-ionized water, and treated with the silver enhancement reagent. After washings in distilled water, samples were stained with 0.5% uranyl acetate in 50% ethanol, dehydrated in a graded series of alcohols, embedded in Epon, sectioned, and examined with a Philips EM300 transmission electron microscope operating at 60 kV.
Transfection Efficiency
The efficiency of the transfection in cells was estimated by the X-gal test. The presence of expressed β-galactosidase was observed by the blue staining with X-gal reagent. For this test, a 4-well permanox Lab-Tek plate (Nunc) was seeded with 4.104 MCF7 cells/well. Plasmids pCMV-β (0.5 or 1 μg) and cationic liposomes at the molar charge ratio X = 2 were complexed and incubated with cells for 48 hr at 37C in 5% CO2. After the incubation, cells were washed with PBS, fixed with paraformaldehyde (1%) and glutaraldehyde (0.02%) for 3 min before the X-gal reagent (1 mg/ml X-gal, 5 mM potassium ferrocyanide, 5 mM potassium ferricyanide, 2 mM MgCl2) was added. Microscopic observation was performed 6 hr later.
Results
Characterization of Liposomes and Efficiency of the Plasmid—Liposome Complex Formation
Characterization of Cationic Liposomes. QLS results indicated a size distribution of liposomes prepared with TMAEC-Chol/DOPE (1:1) or TEAPC-Chol/DOPE in water with mean sizes around 124 nm and 104 nm, respectively, and a polydispersity factor of 0.33 and 0.23. These liposomes were very stable. TEM showed a thickness of about 3 nm for the liposomal membrane, indicating a single bilayer, and demonstrated that liposomes are unilamellar.
Control of Plasmids Labeled with Digoxigenin. Plasmids labeled with digoxigenin were controlled by gel electrophoresis in comparison with native plasmid, as indicated in Figure 1. In this figure there is no difference between the migration of digoxigenin-labeled plasmid (Figure 1, Lane 2) and the native plasmid (Figure 1, Lane 3), suggesting that the circular supercoiled state was preserved.
Efficiency of the Formation of Plasmid—Liposome Complexes
After mixing of the same amount of plasmids with various amounts of liposomes, the solution containing pure plasmids and the solution containing the complexes were exposed to electrophoresis for a gel retardation observation.
Complexing of plasmids pCMV-β with TMAEC-Chol/DOPE liposomes is shown in Figure 2. Lane 1 indicates molecular weight markers. Free plasmids migrated in Lane 2, whereas Lanes 3, 4, and 5 indicate plasmids mixed with TMAEC-Chol/DOPE liposomes in various charge molar ratios, X = 0.5, 2, and 4, respectively. In Figure 2, Lane 3 there is a trace of plasmids not entirely retained by liposomes but there is no print of free plasmids in Lanes 4 and 5, indicating a complete complexing of plasmids with liposomes when the molar charge ratio of lip(+)/DNA is greater than 2.
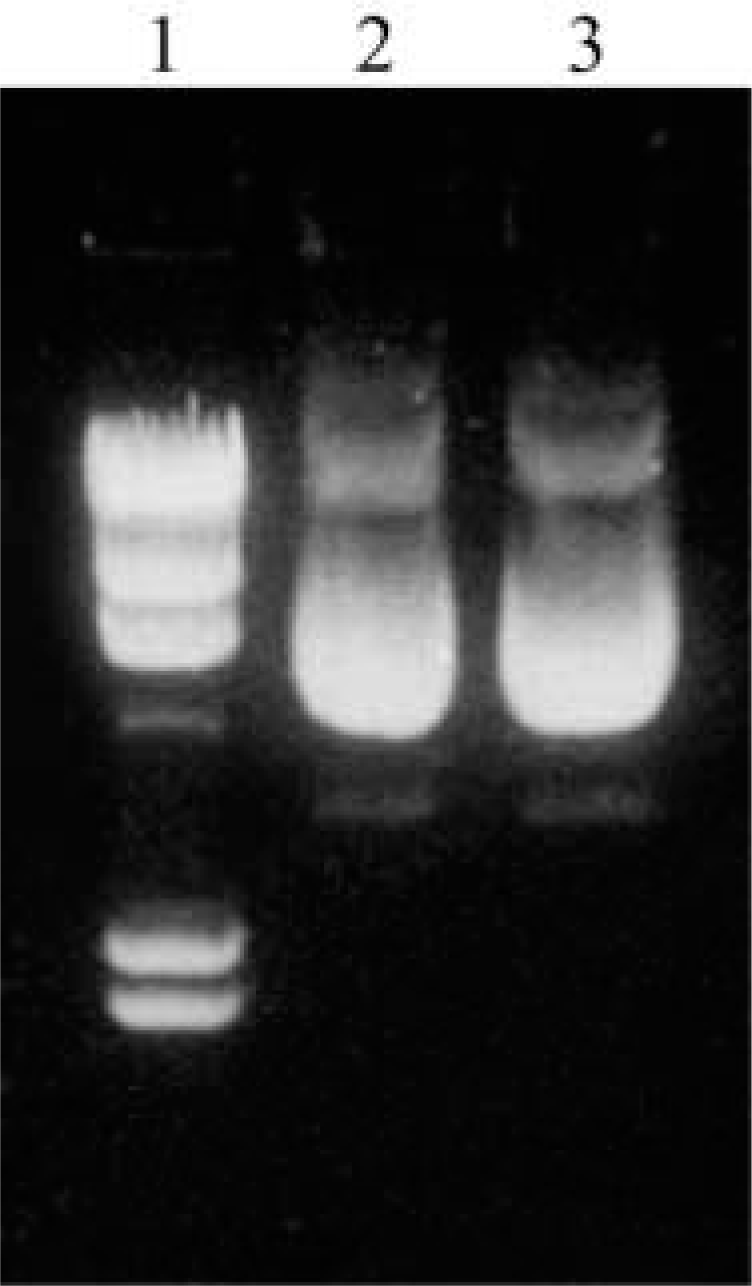
Electrophoresis control of digoxigenin-labeled plasmid. Lane 1, molecular weight markers with λ Hind III; Lane 2, plasmid DNA labeled with digoxigenin as described in the text; Lane 3, unlabeled plasmid for comparison.
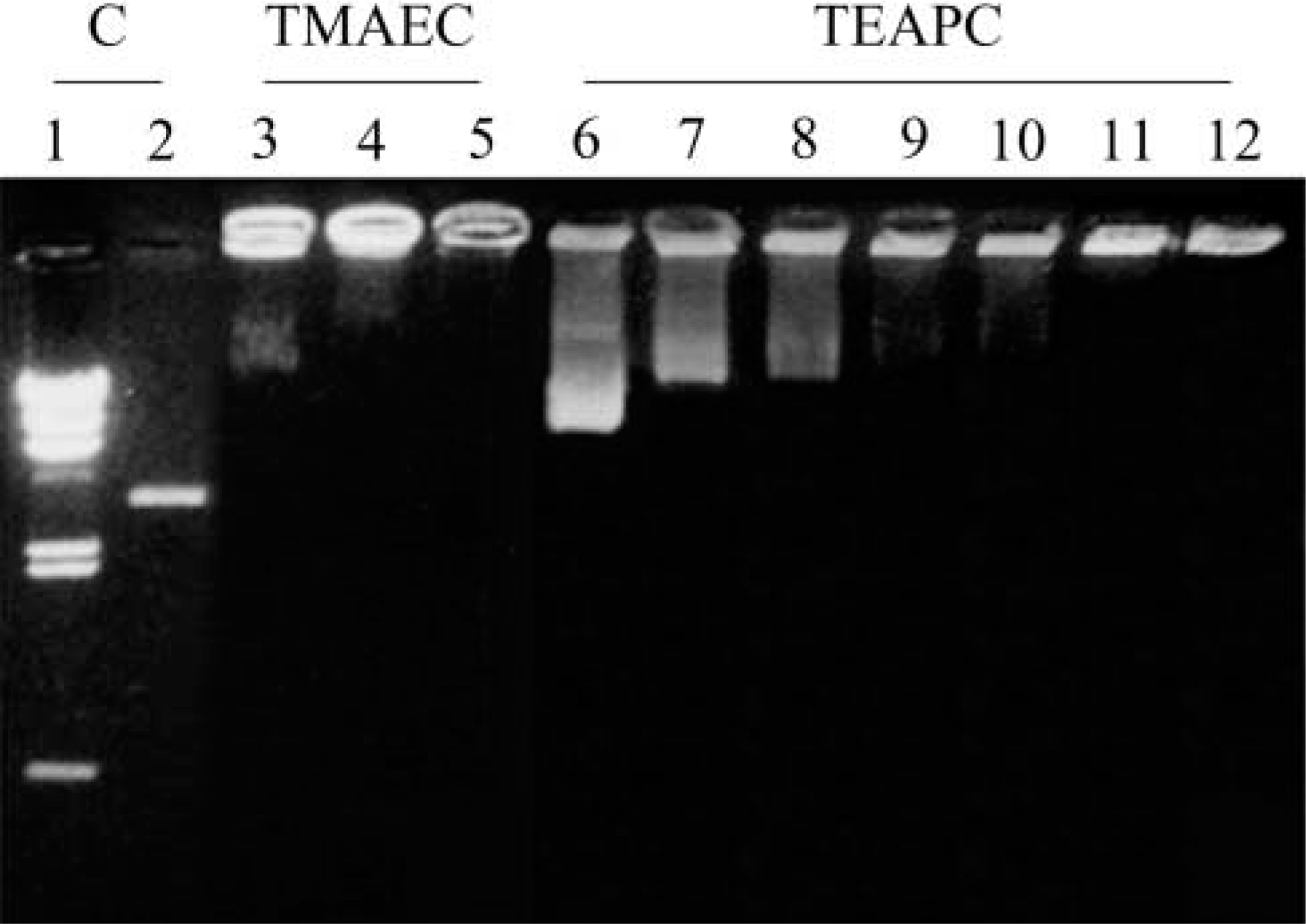
Assay on efficiency in complex formation observed by gel retardation of plasmids pCMV-β complexed with TMAEC-Chol/DOPE and TEAPC-Chol/DOPE (1:1) liposomes. Lanes 1 and 2 for control. Lane 1, molecular weight markers with λ Hind III; Lane 2, plasmid DNA alone. Lanes 3–5 represent the migration of DNA as a function of the molar charge ratio X between cationic lipid TMAEC-Chol and DNA. Lane 3, X = 0.5; Lane 4, X = 2; Lane 5, X = 4. Lanes 6–12, TEAPC-Chol. Lane 6, X = 0.5; Lane 7, X = 1; Lane 8, X = 1.3; Lane 9, X = 1.5; Lane 10, X = 1.7; Lane 11, X = 2; Lane 12, X = 4.
It was the same for pCMV-β complexed with TEAPC-Chol/DOPE liposomes. As shown in Figure 2, Lanes 6–12 show plasmids when mixed with liposomes in various charge molar ratios 0.5, 1, 1.3, 1.5, 1.7, 2, and 4 in the absence of serum. In Lane 10 (X = 1.7) there is a trace of plasmids not entirely retained by liposomes but there is no print of free plasmids in Lanes 11 (X = 2) and 12 (X = 4), indicating a complete complexing of plasmids with liposomes when the molar charge ratio of lip(+)/DNA is greater than 1.7. DNA can be retained 100% when the molar charge ratio exceeds this value.
These results can be explained as follows. One nucleotide (with one negative charge) was bound to one cationic lipid molecule (with one positive charge) on the external surface of the liposomes and therefore twice the number cationic lipid molecules is necessary if the liposomal structure was conserved. This result clearly indicates the electrostatic behavior of the binding between DNA to TMAEC-DOPE liposomes. In all cases, the yield of the formation of liposome—DNA can reach 100% for plasmids when the molar charge ratio is appropriately chosen.
Delivery of Vectorized DNA into MCF7 Cells
Transfection Efficiency. The transfection efficiency was estimated by X-gal (5-bromo-4-chloro-3-indolyl β-
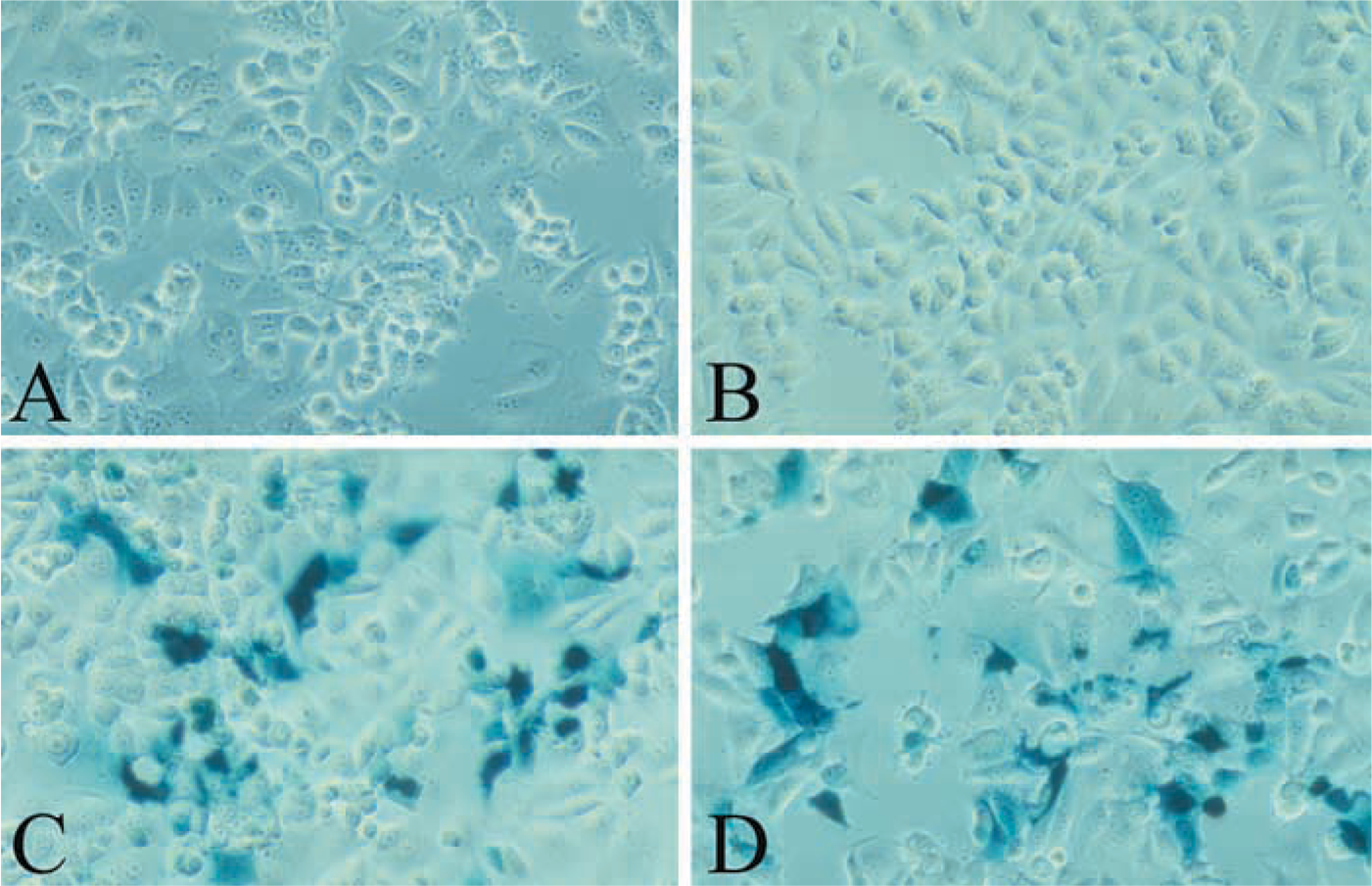
Transient transfection efficiency in MCF7 cells observed by the blue staining with X-gal. Plasmid DNA was complexed with TMAEC-Chol/DOPE liposomes at the molar charge ratio X = 2. Incubation time 48 hr. (
Pathways into MCF7 cells of plasmids pCMV-β vectorized by cationic liposomes TMAEC-Chol/DOPE (1:1) observed by TEM. Plasmids were labeled with digoxigenin and detected by antibodies conjugated to gold particles enhanced by colloidal silver (arrows). (
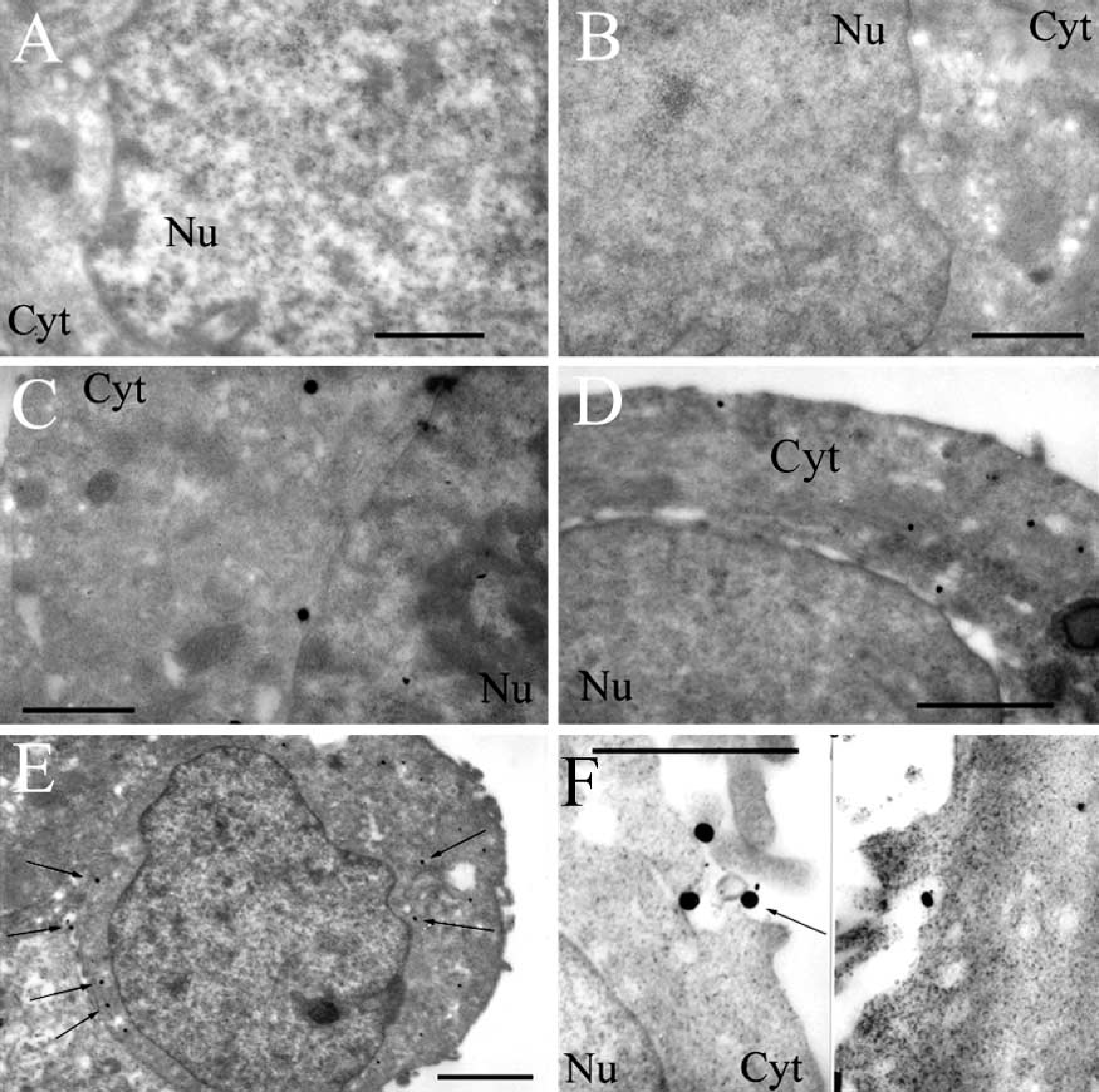
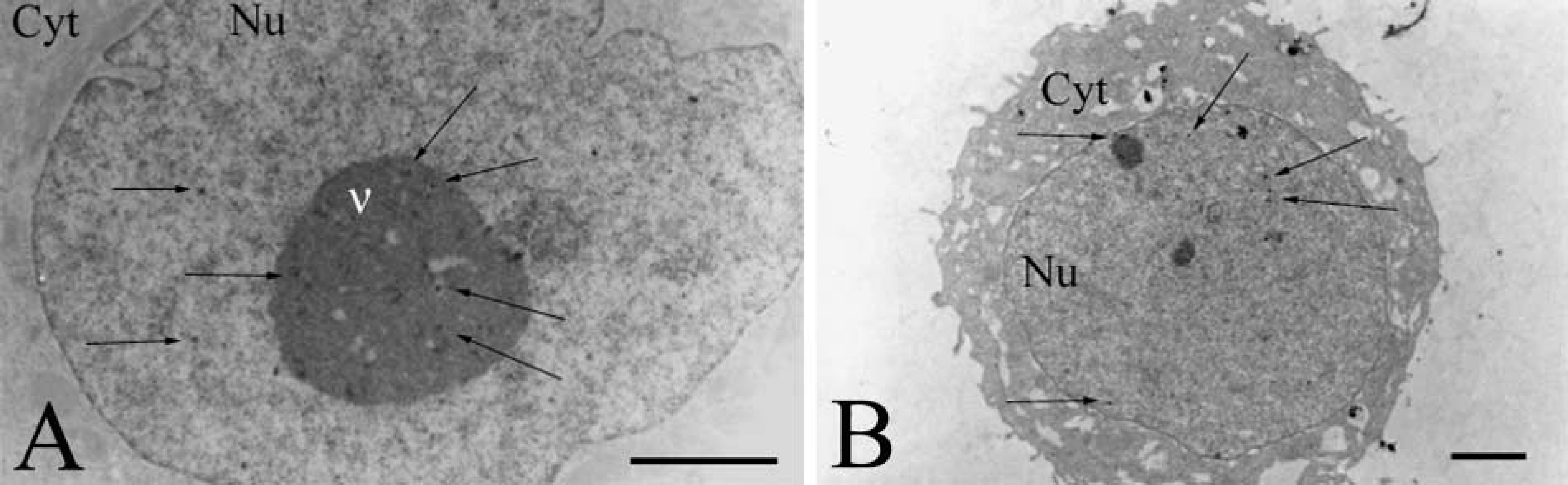
Observation of Plasmids by Immunogold Labeling. To follow the route of plasmids, the latter were labeled with digoxigenin and detected by anti-digoxigenin Fab coupled to ultrasmall gold particles. The size of gold particles was increased by a silver enhancement reagent. Figures 4A and 4B show controls by untransfected cells treated with silver alone or with antibodies followed by silver. The micrographs do not show any dark staining and clearly indicate that, in the absence of plasmids labeled with digoxigenin, there are no dark spots. Figure 4C shows a cell 24 hr after transfection with plasmid pCMV-β unvectorized by liposomes. Few gold particles are observed in the cytoplasm, indicating the presence of rare plasmids.
Figures 4D–4F show cells exposed to plasmids for 15 min and 30 min. At 15 min after incubation of cells with the complexes, black gold particles were seen only at the cell surface or in the cytoplasm but no particles were observed in the nucleus (Figure 4D). At 30 min after incubation, as at 15 min, gold particles were observed only in the cytoplasm and not in the nucleus (Figures 4E and 4F). Cell components lightly stained by uranyl acetate do not contribute to these very dark patterns.
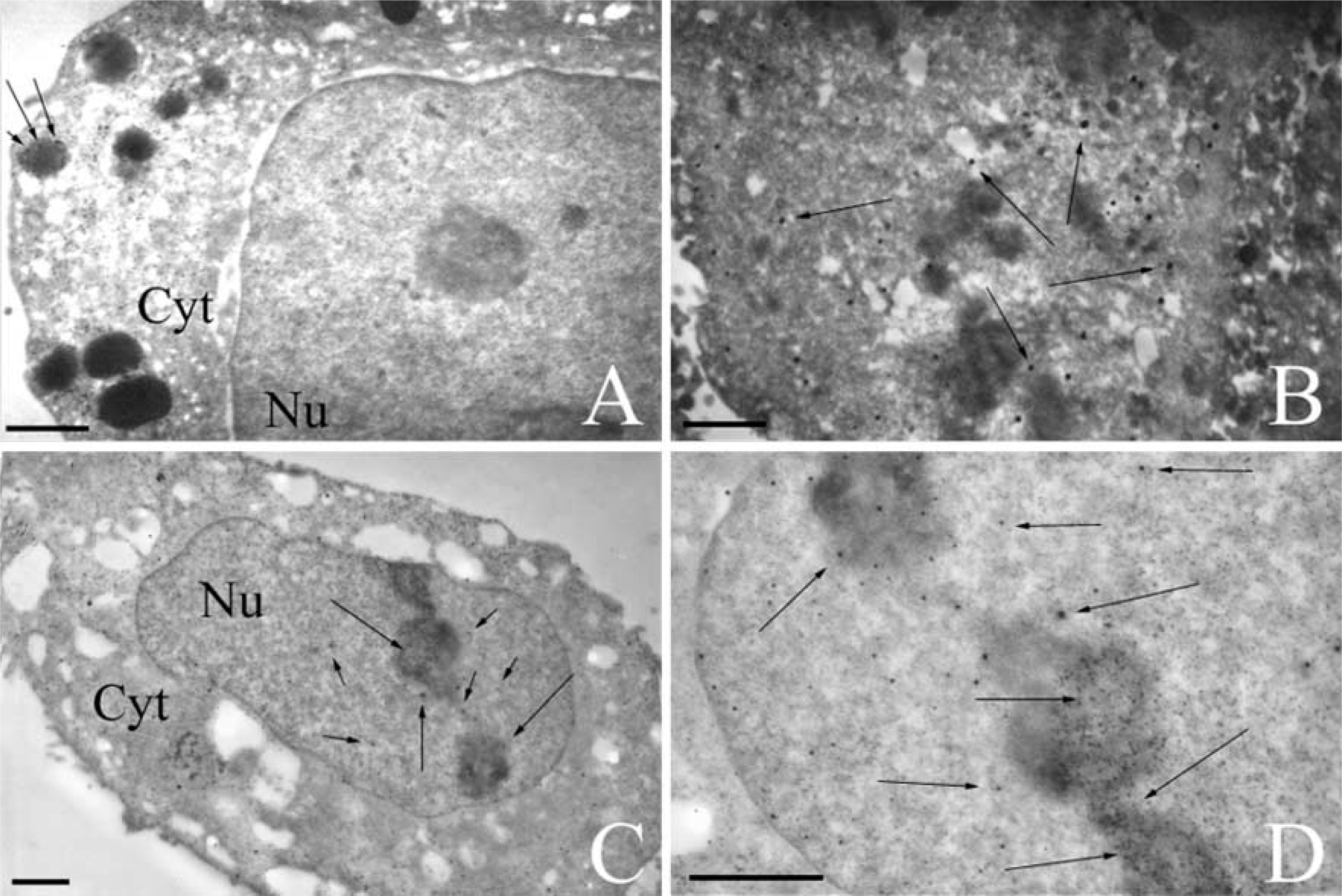
Pathways into MCF7 cells of plasmids pCMV-β vectorized by cationic liposomes TMAEC-Chol/DOPE (1:1) observed by TEM. Plasmids were labeled with digoxigenin, detected by antibodies conjugated with gold particles enhanced by colloidal silver (arrows). (
The micrograph in Figure 5A of an MCF7 cell transfected for 1 hr shows many gold particles but only in the cytoplasm and in electron-dense vesicles. No gold particles were seen in the nucleus. This indicates that plasmids, or at least complexes of plasmid-lipid, are already internalized in the cell. Gold particles in the dense vesicles may represent endosomes containing plasmid-lipid complexes. In the micrographs showing cells after 5 hr (Figure 5B), particles are still present in the cytoplasm and a few begin to appear in the nucleus of some cells. For cells observed at 24 hr after transfection there are many more plasmids in the nucleus, although some gold particles still exist outside the nucleus (Figures 5C and 5D). Occasionally, gold particles are still present in protrusions of the cell surface. Magnifications of some portions of the nuclei show several gold particles in the nucleolus and nucleus (Figures 5C and 5D). After 24 hr of incubation many but not all cells have immunopositive nuclei.
Liposomes prepared from cationic lipid TEAPC-Chol were also used to carry plasmids. As early as 6 hr after addition of complexes to MCF7 cells, plasmids were observed in the nucleus, as shown by a high-magnification field showing gold particles in nuclear compartments (Figure 6A). At 24 hr after incubation, low magnification of an entire transfected cell revealed gold labeling especially in the nucleus, as for TMAEC-Chol/DOPE liposomes (Figure 6B). This indicates the similar behavior of the series of cationic lipids containing a quaternary ammonium head group.
Discussion
The most important result in this work is that, with the immunogold technique, digoxigenin-labeled plasmids transferred by cationic liposomes have been visualized in the nucleus of the transfected cell. Until now, digoxigenin was used only for labeling of oligonucleotides (Tarrason et al. 1995; Lappalainen et al. 1997).
Several controls were performed to ensure the validity of this technique. First, the Dig label of plasmid did not modify the structural integrity, as assessed by electrophoresis. Second, no black staining was observed in cells when samples were submitted to the silver enhancement treatment alone. Third, cells not incubated with the Dig plasmid-liposome complexes but treated with gold anti-Dig antibodies and silver showed no immunoreactivity in the cytoplasm or in the nucleus, indicating that there was no nonspecific binding in cell preparations. When the cells were incubated with the Dig plasmid without cationic liposomes as carriers and further treated by the anti-Dig antibodies and silver, there was no immunoreactivity in most cells. Occasionally in some cells, rare gold particles were observed as shown in Figure 4C. Therefore, with these negative controls the presence of dark particles ensures the presence of gold particles and of Dig-labeled plasmids when plasmids are transferred by cationic liposomes (Figure 5).
Pathways into MCF7 cells of plasmids pCMV-β vectorized by cationic liposomes TEAPC-Chol/DOPE (1:1) observed by TEM. Plasmids were labeled with digoxigenin and detected by antibodies conjugated with gold particles enhanced by colloidal silver (arrows). (
The kinetics of plasmids carried by TMAEC-Chol/DOPE liposomes was studied and showed that, after the cytoplasmic step, TEM can prove that DNA penetrates inside the nucleus, as revealed by immunogold labeling. The time course is respected: at 15 min after transfection only very few plasmid copies were observed in the cytoplasm, and then 24 hr later there are many more plasmid copies, especially in the nucleus. Since we know that previous works have demonstrated lipids only in the cytoplasm and not inside the nucleus, it cannot be doubted that DNA was dissociated from lipids.
To understand these results, it is useful to recall some results of our previous work concerning the ultrastructure of lipoplexes and their cellular pathway (Cao et al. 2000). It was shown by confocal microscopy that, with these liposomes as carriers, after entry into MCF7 cells oligonucleotides were dissociated from the vectors and penetrated into the cell nuclei. For plasmids and with staining with uranyl acetate, TEM indicated the passage of DNA—liposome complexes across the cell membrane by endocytosis, internalization into endosomes, and release into the cytosol. In the present work liposomes were not labeled, but the clusters of gold particles in the electron-dense vesicles observed, as in Figure 5A, undoubtedly correspond to the endosomes observed previously. It appears that the membrane of endosomal compartment was destabilized, allowing the release of DNA from the complexes and then the entry of DNA into the cytosol, as shown by separated gold particles revealing the presence of labeled plasmids. As in most formulations of cationic liposomes (Farhood et al. 1995; Zabner et al. 1995), DOPE was used with TMAEC-Chol and TEAPC-Chol. This destabilization of endosomal membranes may be achieved by cationic lipids (Xu and Szoka 1996; Zelphati and Szoka 1996) or by the co-lipid DOPE (Farhood et al. 1995; Fasbender et al. 1997). In the present case it was not possible to visualize the dissociation of plasmids and liposomes as has been observed with oligonucleotide—liposome complexes in previous work. However, knowing previously that liposomes did not penetrate into the nucleus, it is obvious that, with plasmids detected in the nucleus here, dissociation did occur. On this point, we should emphasize that the dissociation from lipids and the entry into nuclei were observed with modified oligonucleotides (Marcusson et al. 1998) as well as with unmodified oligonucleotides transferred by TMAEC-Chol/DOPE liposomes into MCF7 cells (Cao et al. 2000).
The passage into the nucleus is a necessary step for the observed expression of the gene. This passage across the nuclear envelope may be due to various mechanisms, such as mitosis (Mortimer et al. 1999; Tseng et al. 1999), or by a mechanism that enables the association of plasmids with nucleus-targeted proteins that carry DNA into the nuclei. A “facilitated process” implying a role of karyophilic proteins has been also suggested (Dowty et al. 1995). Transport across nuclear envelopes may also be stimulated by signal-mediated factors (Nigg 1997).
It is worth noting that in previous works (Zabner et al. 1995; Friend et al. 1996; Escriou et al. 1998; El Ouahabi et al. 1999), using various techniques to visualize plasmids, only perinuclear localization was observed, so that the expression of the genes used in these works remain unclear. The passage of plasmids observed in the nucleus here is in full accordance with the expression of a gene encoding β-galactosidase in MCF7 cells observed by the X-gal test and luminometric measurements described previously (Cao et al. 2000).
Finally, it is important to emphasize that high efficiency can be obtained in the complexing of DNA and TMAEC-Chol/DOPE or TEAPC-Chol/DOPE liposomes. This high efficiency is confirmed with the plasmid, as demonstrated by gel retardation, that complete complexing occurred when the molar ratio was higher than 1.7 or 2, depending on the lipid.
In conclusion, the results of this work show that the digoxigenin-labeled plasmid DNA immunogold procedure is a useful method to detect exogenous DNA in cells by transmission electron microscopy. With this technique, we showed that plasmids are efficiently carried into MCF7 cells by cationic liposomes prepared from TMAEC-Chol and TEAPC-Chol. The functionality of the reporter genes carried by these liposomes was preserved inside the nuclei, resulting in the observed expression of β-galactosidase in accordance with the passage of plasmids into the nuclei.