
Abstract
Bone morphogenetic proteins (BMPs) are characterized by their ability to induce osteoblastic differentiation. However, the mechanism of osteo-induction by BMPs has yet to be determined. Using differential display we previously identified AJ18, a zinc finger transcription factor, as an immediate-early response gene to BMP-7. AJ18 was shown to bind to the osteoblast-specific element2 (OSE2) and to modulate transactivation by Runx2, a master gene in osteoblastic differentiation. Here we describe the temporal and spatial expression of AJ18 in developing mouse tissues. AJ18 mRNA expression was observed in most tissues, except liver, and was generally highest early in embryonic development, decreasing markedly after parturition. Consistent with immunohistochemical analysis, AJ18 mRNA expression was highest in the brain, kidney, and bone of 17 dpc (days post coitum) embryos. In endochondral bones of embryonic and 4-week-old mice, immunostaining for AJ18 was strong in the nuclei of proliferating and pre-hypertrophic chondrocytes, and osteoblasts, whereas there was low or no staining in hypertrophic chondrocytes. In teeth of embryonic and 4-week-old mice, nuclear staining was observed in precursor and mature ameloblasts, odontoblasts, and cementoblasts, respectively. In addition, in 4-week-old mice staining of AJ18 was observed within alveolar bone cells and periodontal ligament cells. In general, the spatial expression of AJ18 in skeletal and non-skeletal tissues of mouse embryos showed striking similarity to the expression of BMP-7 mRNA. Therefore, the expression of AJ18 is consistent with its perceived role as a transcriptional factor that regulates developmental processes downstream of BMP-7.
Organogenesis involves temporospatially regulated morphogenesis of cells that differentiate to form organ-specific structures in the appropriate location (reviewed by Peters and Balling 1999). At the earliest stages of organ development, coordinated sequential and reciprocal interactions occur between adjacent epithelial and mesenchymal tissues, and it is evident that growth factor-mediated signaling is necessary for the regulation of the epithelial-mesenchymal interactions. The majority of these growth factors are encoded by a small number of conserved multigene families, which include the bone morphogenetic proteins (BMPs). Although BMPs provide the primordial signals for bone formation, the importance of BMPs in epithelial-mesenchymal interactions has also been demonstrated. For example, BMP-4 expression in dental epithelium induces its own expression along with that of transcription factors Msx1, Msx2, and Egr1 in mesenchyme (Vainio et al. 1993). Moreover, BMP-7 is expressed in both the ureteric epithelium and the undifferentiated mesenchyme of the kidney, and is required for the continued proliferation and differentiation of mesenchymal and/or epithelial cells (Dudley et al. 1995; Luo et al. 1995). BMP-7 is also required for the development of mesenchyme and epithelium during eye development.
AJ18, a member of the growing family of KRAB/C2H2 zinc finger genes, was originally identified in fetal rat calvarial cells (FRCCs) as a target gene for BMP-7 (Jheon et al. 2000). Subsequent studies have shown that the protein is differentially regulated during osteogenic differentiation, that it has DNA-binding activity, and that it can modulate Runx2-mediated transactivation through the OSE2 (Jheon et al. 2001).
To determine the physiological significance of AJ18 in bone development, preliminary analyses of AJ18 mRNA expression were performed in rat tissues (Jheon et al. 2001). Whereas expression of AJ18 was relatively strong in embryonic bone, expression was also observed in other tissues, including brain and kidney. However, no expression was observed in liver tissues. In our studies, a combination of Northern hybridization and immunohistochemistry (IHC) was used to analyze the temporal and spatial expression of AJ18 in mouse embryos to provide a more comprehensive and detailed analysis of tissues and cells that express AJ18 during embryonic development. The isolation and sequencing of mouse AJ18 confirmed that polyclonal antibodies raised against rat AJ18 would also recognize the mouse protein. Mouse AJ18 was later found to be the same gene as Kid3 and, as shown for AJ18, is expressed highly in brain and is differentially expressed during kidney development (Watson et al. 2000). The studies presented here have revealed an expression pattern of AJ18 similar to that reported for BMP-7 (Helder et al. 1995), which affects tissue and organ development during embryogenesis. These observations further support that AJ18 is a downstream target of BMP-7.
Materials and Methods
Mouse Genomic Library Screen
To characterize the mouse AJ18 gene, a mouse genomic library was prepared from 129SvJ mouse spleen DNA assembled into λ-phage FIX II vector (Stratagene; La Jolla, CA) and screened by plaque hybridization (Sambrook et al. 1989). The library was screened with a [32P]-dCTP-labeled cDNA probe, which was digested from rat AJ18 cDNA by PstI, generating a 461-bp fragment. After tertiary screening, positive clones were isolated, purified, and characterized by restriction endonuclease digests. EcoRI digested fragments were separated on an agarose gel, subcloned into pBluescript II SK (Stratagene), and sequenced. Mouse AJ18 cDNA was deduced from sequence comparison with rat AJ18 cDNA.
Mouse Tissue Preparation
Tissues from brain, heart, lung, skeletal muscle, cartilage, liver, thymus, kidney, calvaria, and long bones were dissected from mice at embryonic stages 15 and 17 dpc and at Days 2, 7, 15, and 23 after birth. Mouse embryos at 16 dpc and mandible and tibia from a 4-week-old mouse were isolated and fixed in 4% paraformaldehyde-PBS at 4C. The mandible and tibiae were demineralized in 12.5% (w/v) EDTA (pH 7.4), with the solution changed other day for 2–3 weeks. The embryos and tissues were embedded in paraffin. Serial 12-μm-thick sections were mounted on Superfrost/Plus glass slides (Fisher Scientific; Nepean, Canada) and stored at 4C until use.
RNA Extraction
Total RNA was isolated from mouse tissues using the thiocyanate-phenol-chloroform extraction method as described by Chomczynski and Sacchi (1987).
Northern Blot Hybridization
Northern blot hybridization was performed as described previously (Jheon et al. 2001). A cDNA fragment of 700 bp within the 3′-UTR of AJ18 was PCR-amplified from the isolated mouse λ-clone using primers 5′-GGCACACCTTTCATCTACGGCTCATCC-3′ and 5′-ACGCCTGTATCCATCCCCACTGTTAAG-3′, and was used as the template for probe synthesis. The intensity of the AJ18 mRNA bands was visualized with a PhosphorImager (Molecular Dynamics; Sunnyvale, CA) and quantified using the ImageQuant software (Molecular Dynamics). Relative expression levels of AJ18 mRNA were determined by normalization to expression levels of glyceraldehydephosphate dehydrogenase (GAPDH).
Anti-AJ18 Polyclonal Antibodies
Polyclonal antibodies were raised against peptides specific for rat AJ18 and affinity-purified as described in Jheon et al. (2001).
IHC Analysis
Immunoperoxidase staining for AJ18 protein in tissue sections from 11, 12, 13, 14, 15, and 16 dpc mouse embryos and from 4-week mouse mandible and tibia was performed using the Vectastain ABC kit (Vector Laboratories; Burlingame, CA) according to the manufacturer's instructions. Tissue sections, prepared as described above, were rehydrated through incubation in graded ethanol to water and incubated in blocking solution (5% BSA, 2% normal goat serum) for 1 hr. Affinity-purified anti-AJ18–1 or anti-AJ18–2 antibodies were applied and tissue sections were incubated for 1 hr. The sections were washed and treated with biotinylated anti-rabbit IgG for 30 min, followed by incubation with peroxidase-labeled streptavidin for 30 min, and subsequently incubated with diaminobenzidine tetrahydrochloride (DAB) and H2O2 for 15 min. All incubations were performed at 21C. Various sections were counterstained with hematoxylin. All sections were visualized under a light microscope (Eclipse 400; Nikon Canada, Mississauga, Canada) and photographed using a Coolpix 950 digital camera (Nikon Canada). All immunostaining was performed at least three times on sections obtained from at least three individual mice (embryos and 4-week-old).
Results
Identification of Mouse AJ18
The sequence of the mouse AJ18 open reading frame (ORF) was deduced from an ∼10-kb λ-clone that was isolated from mouse genomic screens. Because of the high conservation of the nucleotide sequence between mouse and rat AJ18, the mouse AJ18 gene was readily identified and the sequence of the ORF determined from a comparison of the mouse genomic sequence to the rat cDNA. This was simplified, in part, because the 11 C2H2 zinc finger motifs are over 97% identical and are encoded by a single exon. In total, the translated protein sequences of rat and mouse AJ18 share 91% identity and 94% similarity (Figure 1). Whereas the N-terminus, KRAB domain, and zinc finger regions of mouse and rat AJ18 are almost perfectly conserved, showing between 97% to 100% identity, the linker sequence between the KRAB domain and zinc finger motifs and the C-terminus show relatively low conservation with 72% and 79% identity, respectively.
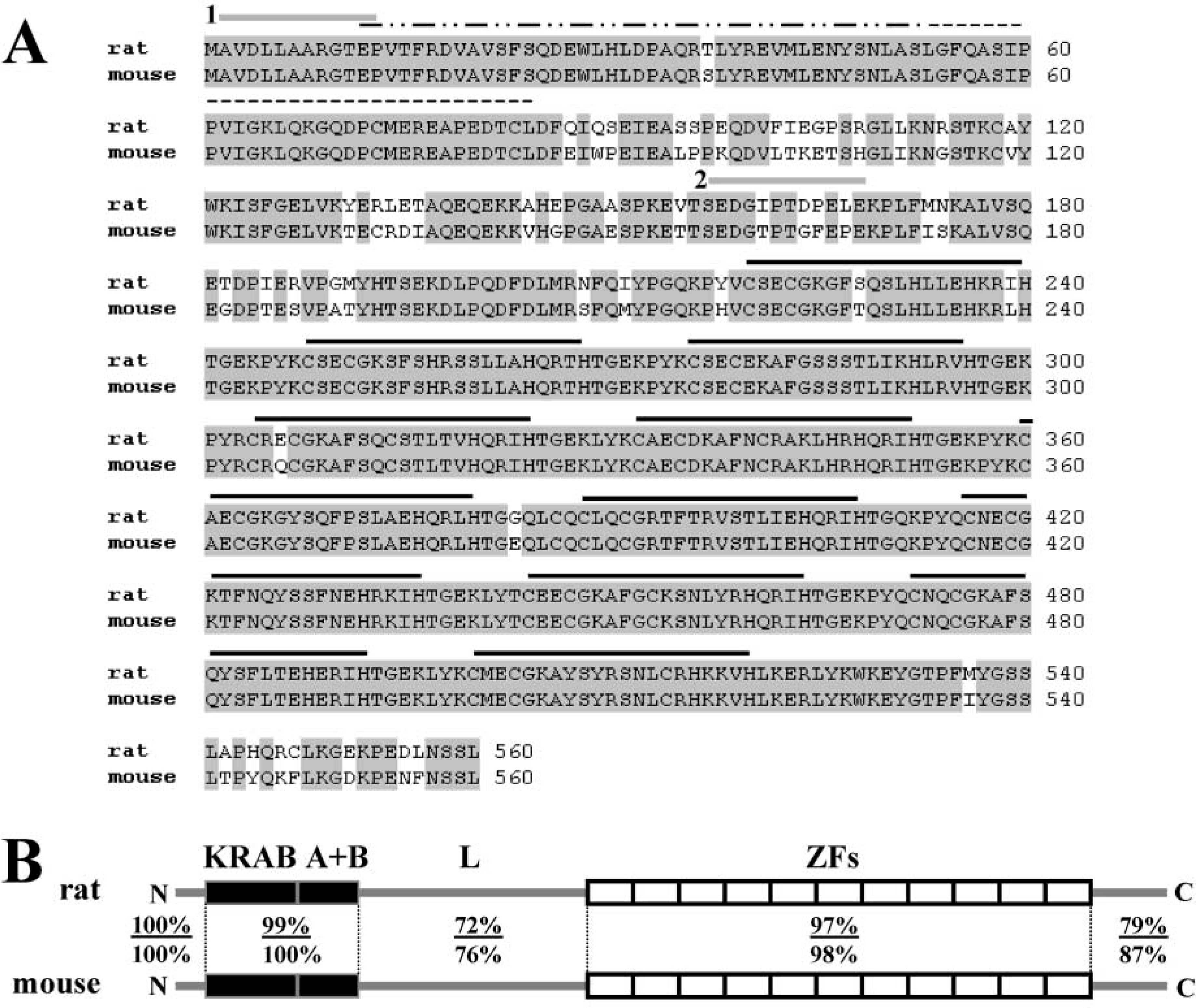
Sequence alignment of rat and mouse AJ18. (
Northern Blot Hybridization
In previous studies, expression of AJ18 mRNA in whole mouse embryos was first detected in 11 dpc embryos, reached a peak at 15 dpc, and decreased in 17 dpc embryos (Jheon et al. 2001). To determine the developmental expression of AJ18 mRNA in different mouse tissues, total RNA was extracted from tissues dissected at various stages of embryonic and neonatal development, Northern blot analyses performed, and the relative intensities of AJ18 mRNA normalized to expression levels of GAPDH mRNA were plotted on a histogram (Figure 2). In 15 dpc embryos, expression of AJ18 mRNA was evident in all tissues and organs analyzed, except the liver. Relatively high expression of AJ18 mRNA was observed in brain and bone tissues. In 17 dpc embryos, expression of AJ18 mRNA remained high in the brain but was lower in the other tissues analyzed compared to levels in 15 dpc embryos. Thereafter, in Day 2 and Day 7 neonates (Figure 2), in 15- and 23-day old mice (data not shown), and in adult rats (Jheon et al. 2001), AJ18 mRNA was expressed highly in the brain, with expression barely detectable in bone and other tissues. Notably, AJ18 mRNA was not detected in liver at any of the developmental stages analyzed (Figure 2; Jheon et al. 2001).
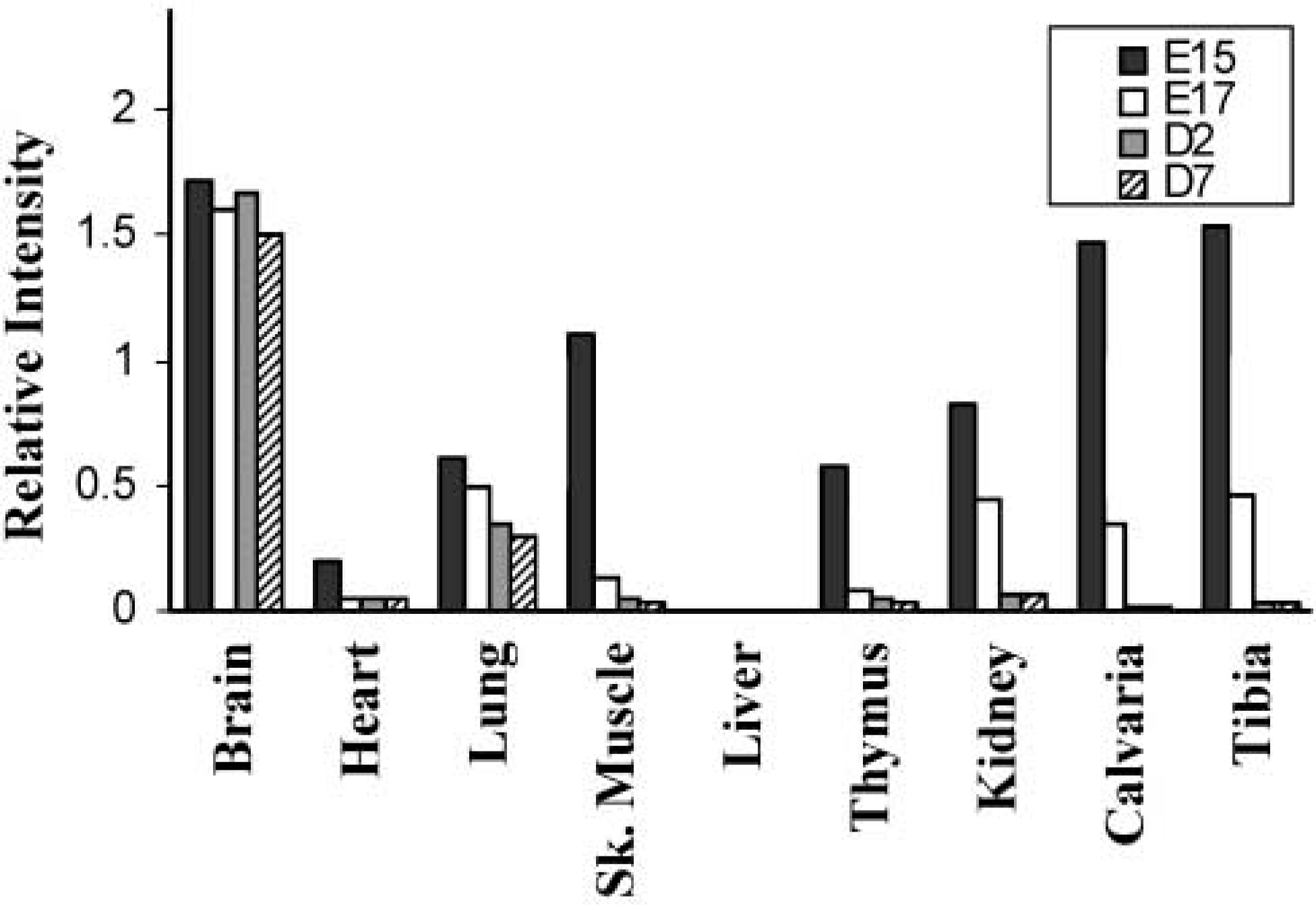
Preliminary Northern blot analyses of AJ18 mRNA expression during mouse development. Various tissues were dissected from mouse embryos and neonates and total RNA extracted for Northern blotting analyses. Blots were hybridized with radiolabeled probes synthesized from AJ18 cDNA. A cDNA probe for glyceraldehydephosphate dehydrogenase (GAPDH) was used as a control for RNA loading. Mouse embryos 15 dpc, E15; 17 dpc, E17; mouse neonates Day 2, D2; Day 7, D7.
IHC Staining
In concert with Northern blot analyses, IHC analyses were performed on tissue sections of whole mouse embryos at various developmental stages and in neonatal tissues. Because the mouse and rat AJ18 sequences are highly conserved, the anti-rat AJ18 polyclonal antibodies raised to synthetic peptides 1 (residues 2–13) and 2 (residues 158–169) should recognize both rat and mouse AJ18 protein. Affinity-purified anti-AJ18–1 and anti-AJ18–2 antibodies recognized mouse AJ18 protein (data not shown), although anti-AJ18–1 showed higher immunoreactivity, consistent with the 100% conservation of sequence between residues 2 and 13 (Figure 1). Therefore, all the tissue sections presented hereafter were immunostained using affinity-purified anti-AJ18–1 antibody. Strongest staining was obtained for all tissues at Day 16 dpc, at which time skeletal tissues could be analyzed at different developmental stages in the ribs. Consequently, the results present here are primarily for tissues at this stage (Figures 3–5, 7, 8, and 10). Although staining for AJ18 was observed in the same tissues beginning at 13–14 dpc mouse embryos, staining intensity was lower relative to 16 dpc mouse embryos, and the differential expression within and between tissues was less clear. To analyze expression of AJ18 in further developed tibial bone and teeth, sections from a 4-week-old mouse were immunostained (Figures 6 and 9).
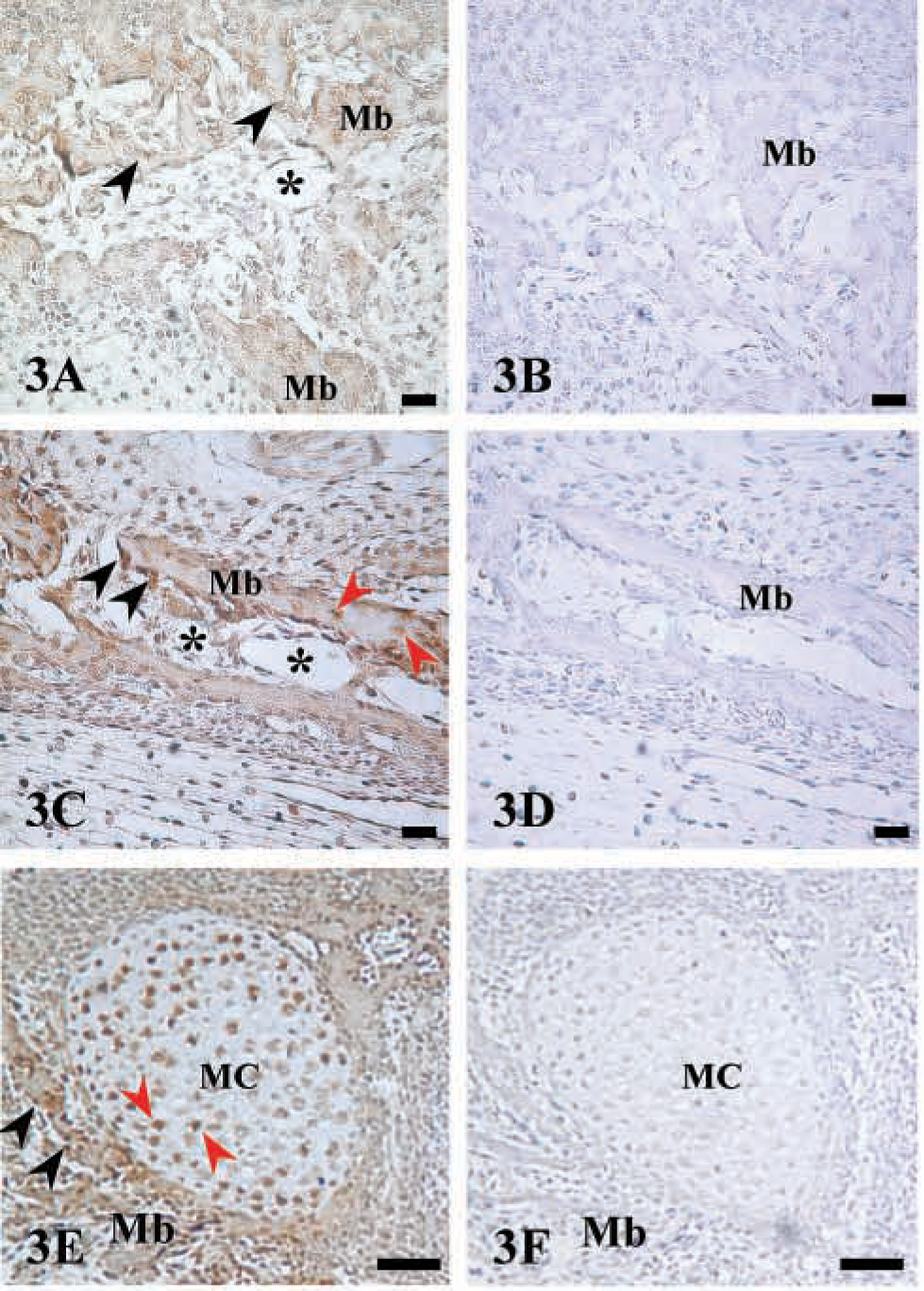
Immunostaining of AJ18 in jaw bones and Meckel's cartilage. Light micrographs of maxillar and mandibular bones of a 16 dpc mouse embryo immunostained for AJ18. (
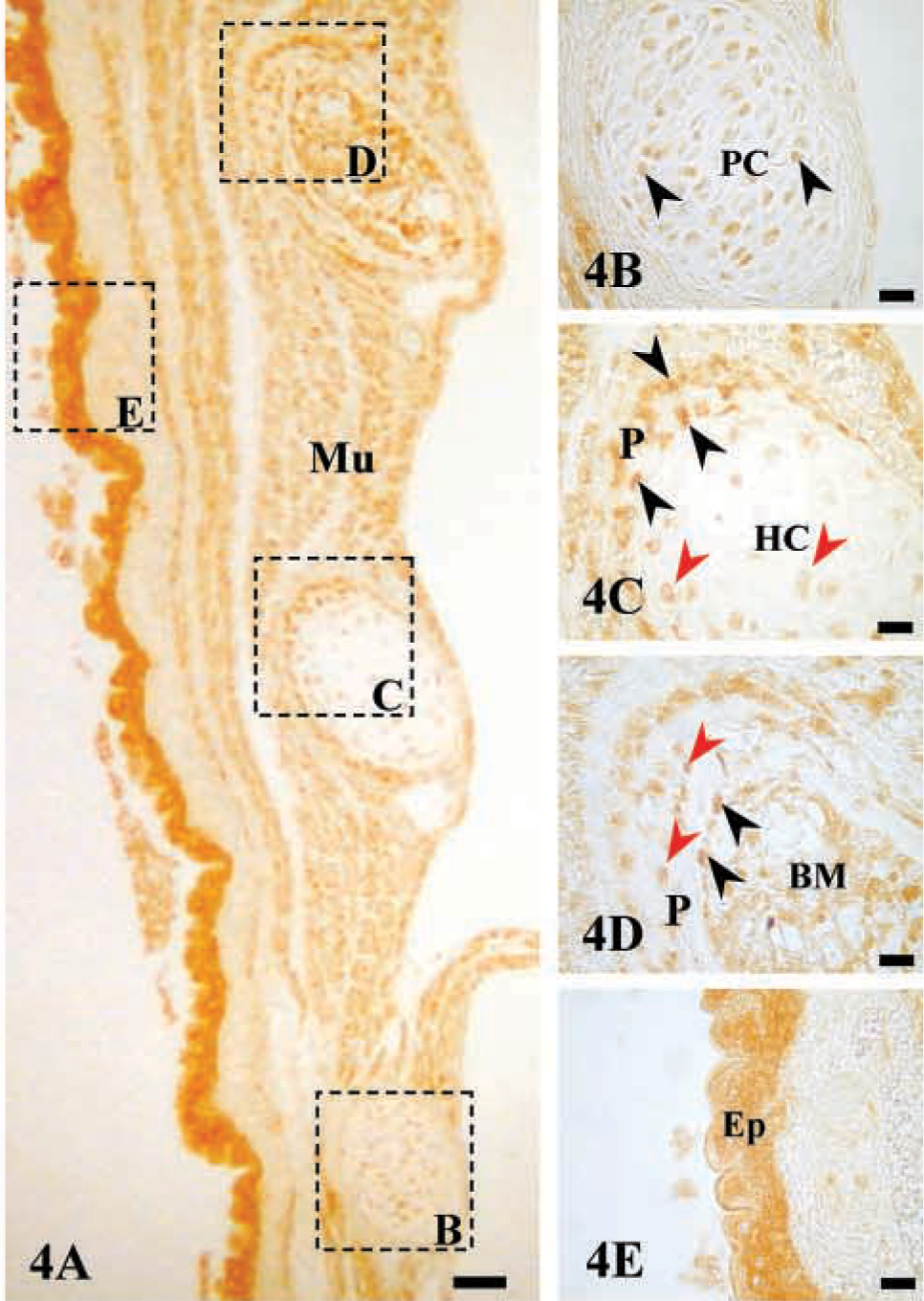
Immunostaining of AJ18 in developing rib bones. Light micrographs of developing rib bones of a 16 dpc mouse embryo immunostained for AJ18. (
Immunostaining of AJ18 in an embryonic rib bone. (
Immunostaining of AJ18 in long bone of 4-week-old mouse. (
Expression of AJ18 Protein in Cartilage and Bone
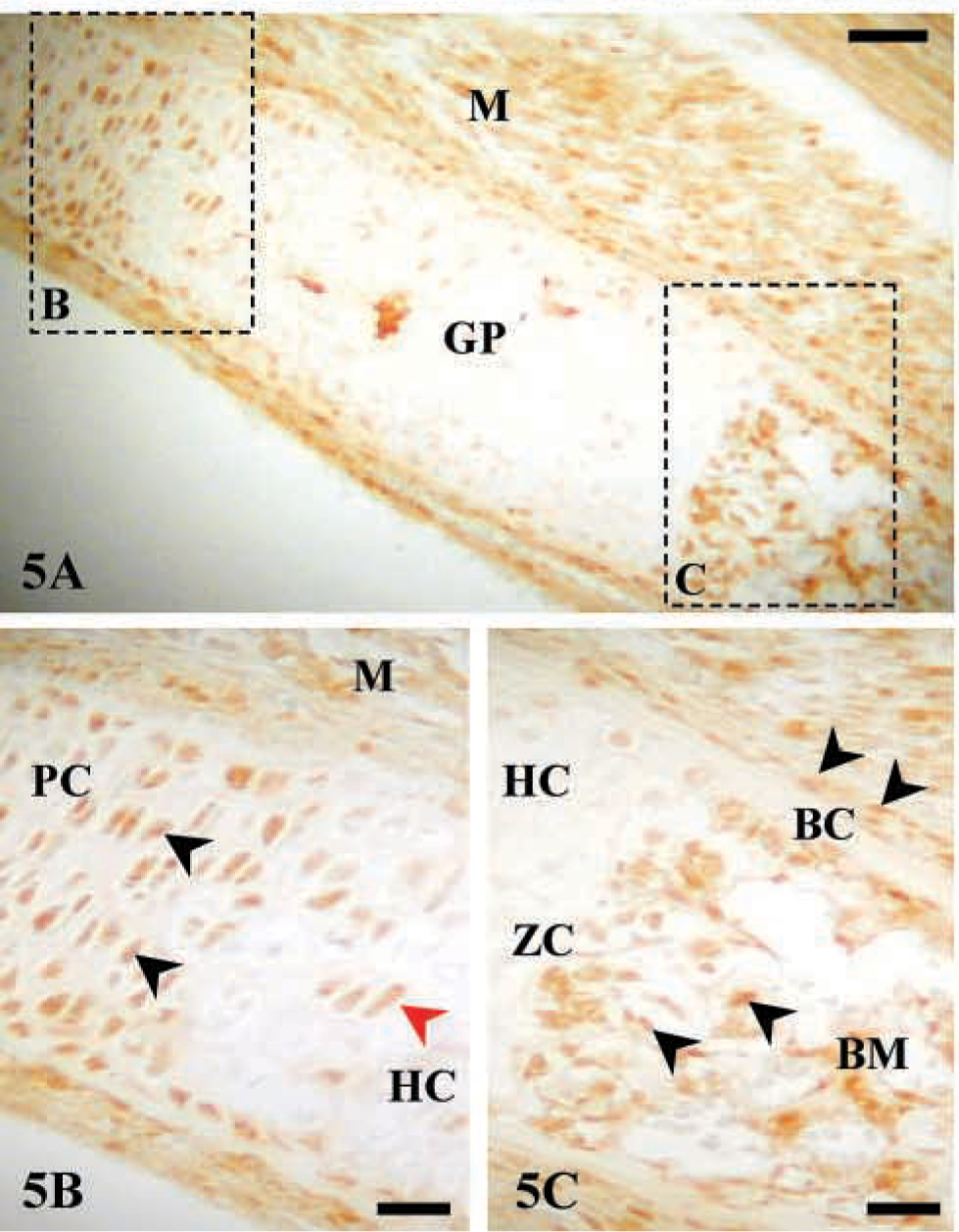
Immunostaining of AJ18 was high in skeletal tissues of 16 dpc mouse embryos, as shown for bone of the maxilla and mandible, Meckel's cartilage, and ribs (Figures 3–5). During embryonic development, mesenchymal cells in the facial region condense to form nodules of differentiated osteoblasts that form the maxillar and mandibular bones of the upper and lower jaws, respectively. Expression of AJ18 was evident in the nuclei of mesenchymal cells adjacent to the bone surfaces, with stronger staining of the osteoblasts lining mineralized bone surfaces (Figures 3A and 3C). AJ18 was also detected within some newly formed osteocytes, but no staining was apparent in the endothelial cells surrounding blood vessels or in loose connective tissue (Figure 3C). AJ18 was detected within the nuclei of the small chondrocytes of Meckel's cartilage, which forms the template for mandibular bone (Figure 3E). In identically prepared control sections incubated without the primary antibody, no signal was observed (Figures 3B, 3D, and 3F). Because the individual ribs form at different times, a progression of endochondral bone development could be seen at a single time point in a transverse section through a series of ribs. Thus, staining for AJ18 in the 10th, 11th, and 12th ribs showed protein in nuclei of cartilage and bone cells (Figure 4A). In the 12th rib, nuclear staining of AJ18 was evident in pre-hypertrophic chondrocytes (Figure 4B). In the 11th rib, which is further developed, the cartilage cells had undergone hypertrophy and cell nuclei were stained sporadically for AJ18 (Figure 4C). At this stage, when the cartilage had started to mineralize and the osteogenic periosteal cells had condensed, AJ18 was present in the nuclei of cells on the surface of the mineralizing bone. In the more advanced 10th rib, endochondral bone had formed centrally, replacing the mineralized cartilage, and the formation of periosteal bone was well established (Figure 4D). Staining for AJ18 was observed in cells present in bone marrow, in the periosteal cells on the surface of the rib bone, and in osteocytes present within periosteal bone. In these sections, comparable staining was also present in skeletal muscle tissue surrounding the ribs (Figure 4A) and was particularly strong in the outer epithelial layer of the embryo (Figure 4E). In skeletal muscle and especially in the epithelium, diffuse staining was seen in the cells, with nuclear staining in the epithelium being strongest in the basal cell layer.
A longitudinal section through the growth plate of the 3rd rib from a 16 dpc mouse embryo showed the developmental progression of cartilage cells through the formation of columnar proliferating chondrocytes to hypertrophic chondrocytes in the mineralized cartilage that is subsequently replaced by epiphyseal bone (Figure 5A). The strongest staining was seen in the proliferating chondrocytes with lower expression in hypertrophic chondrocytes (Figures 5A and 5B), consistent with the staining in the 11th and 12th ribs. However, staining was high in the osteogenic cells in the primary spongiosa immediately below the calcified cartilage and in the subperiosteal bone collar formed by periosteal cells (Figure 5C). Staining in skeletal muscle was also observed.
Immunostaining of AJ18 in developing teeth. Light micrographs of molar tooth development from the maxilla and mandible of a 16 dpc mouse embryo. (
Immunostaining of AJ18 in molar and incisor of 4-week-old mouse. Light micrographs of the mandible from a 4-week old mouse. (
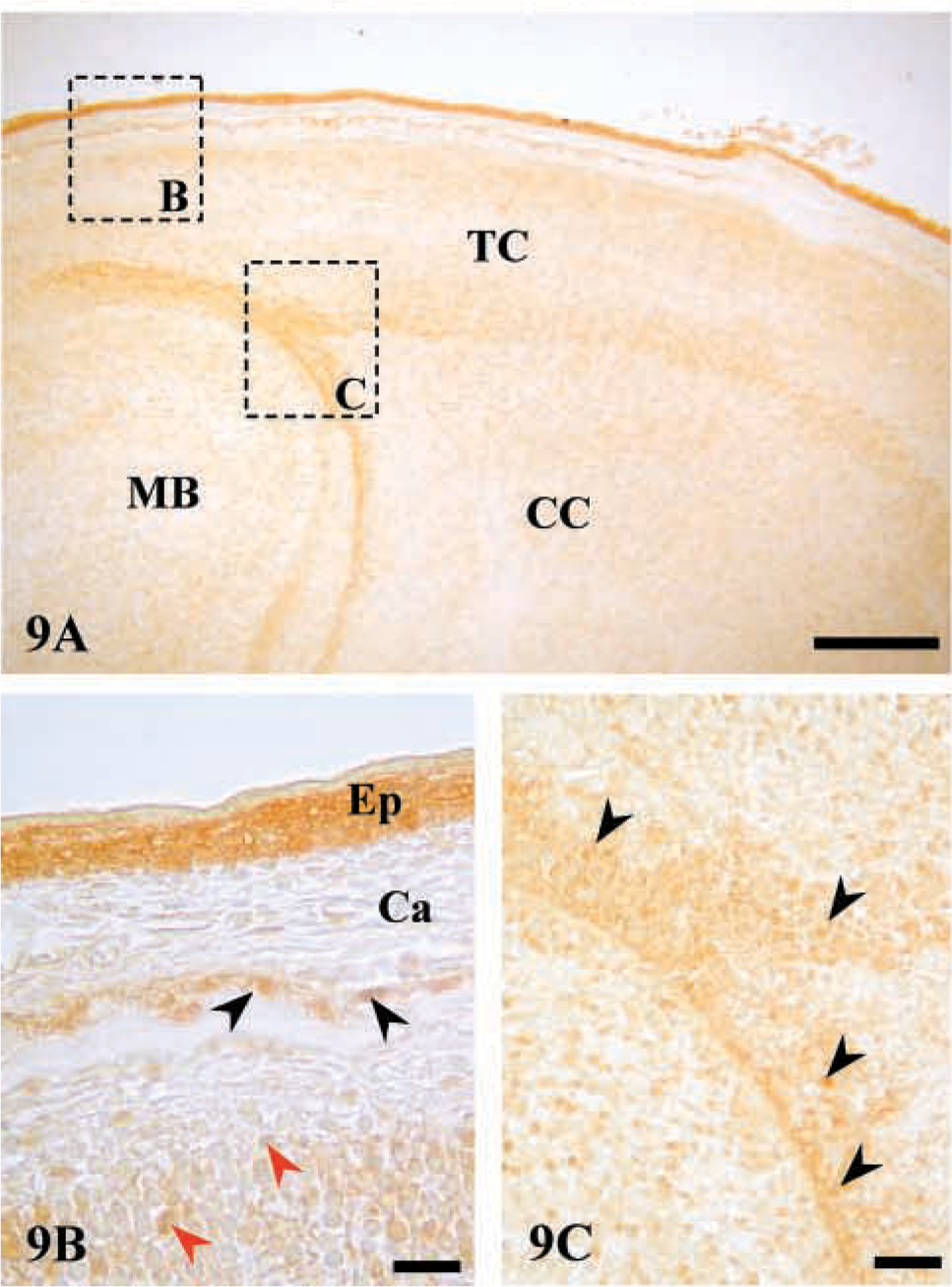
Immunostaining of AJ18 in the brain. (
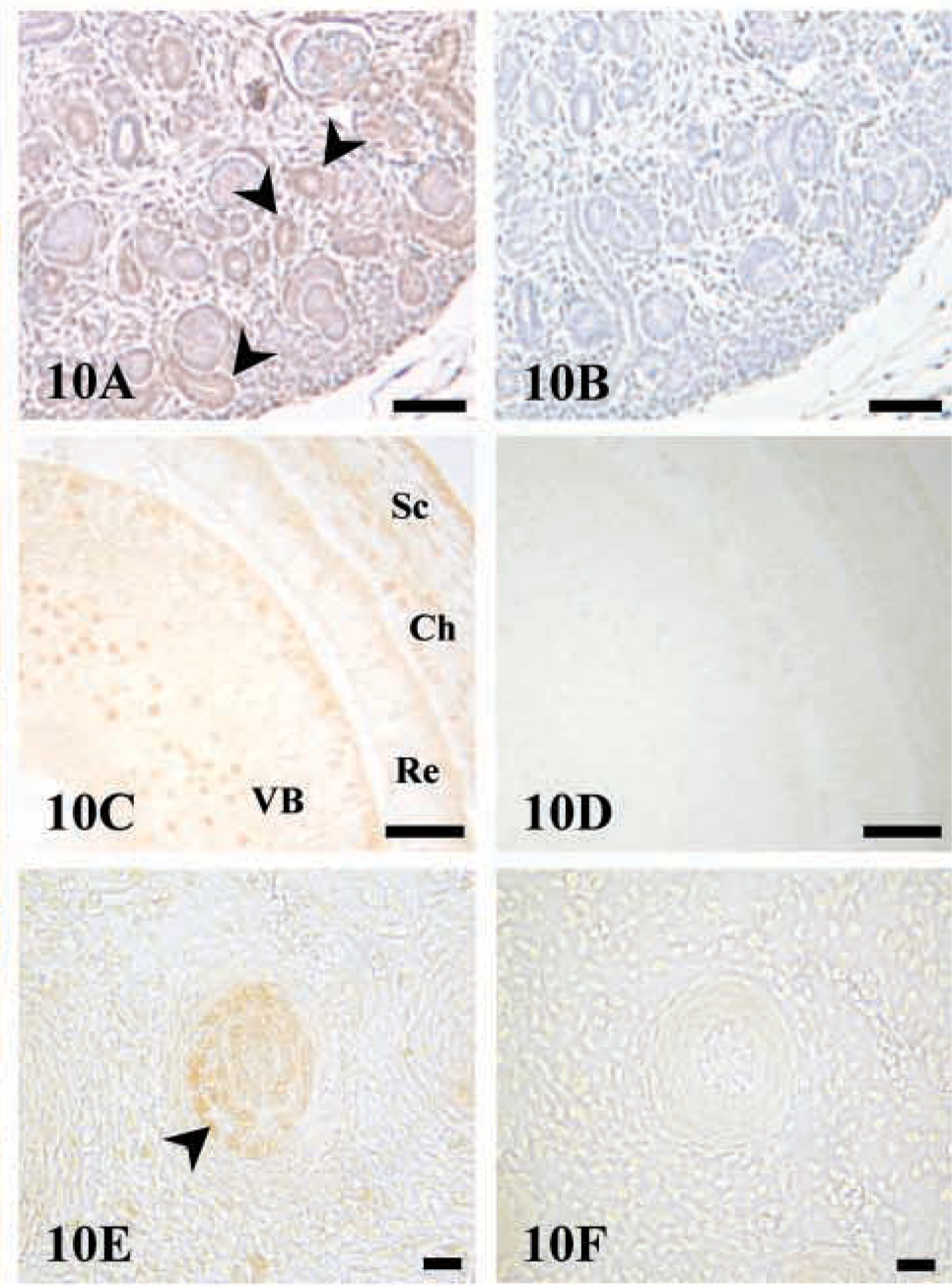
Immunostaining of AJ18 in tissues that express BMP-7. Light micrographs of 16 dpc mouse embryo sections. (
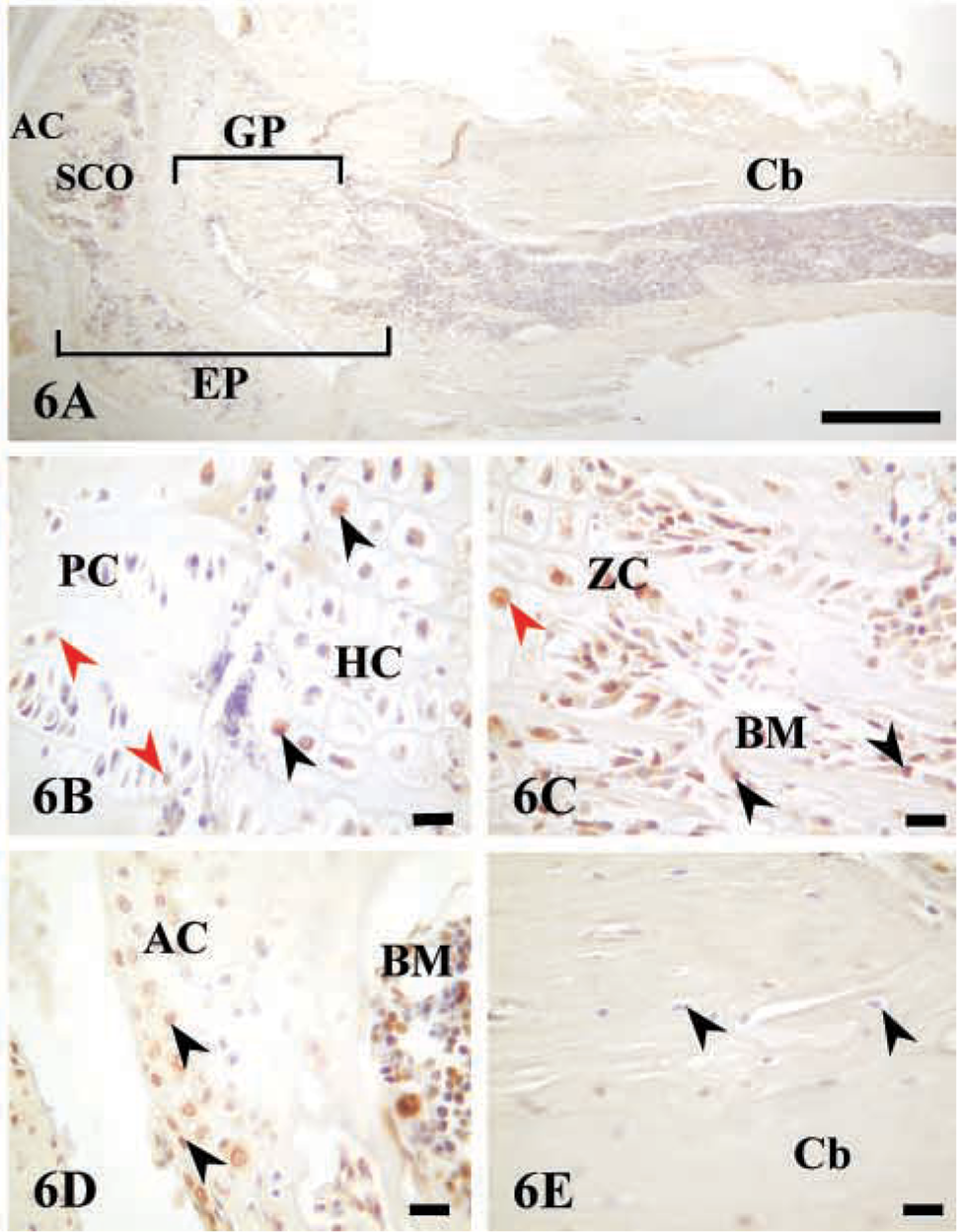
To examine AJ18 expression at later stages of endochondral bone formation, sections of tibiae from a 4-week-old mouse were analyzed (Figure 6A). AJ18 staining was observed in some pre-hypertrophic and hypertrophic chondrocytes (Figure 6B), with strong staining visible in osteoblasts (Figure 6C). Staining for AJ18 was also present in the cartilage cells at the periarticular surface (Figure 6D). No staining for AJ18 was observed in osteocytes embedded within cortical bone (Figure 6E). These results were similar to those obtained in tibiae from a 4-week-old rat (Jheon et al. 2001).
Expression of AJ18 Protein in Developing Teeth
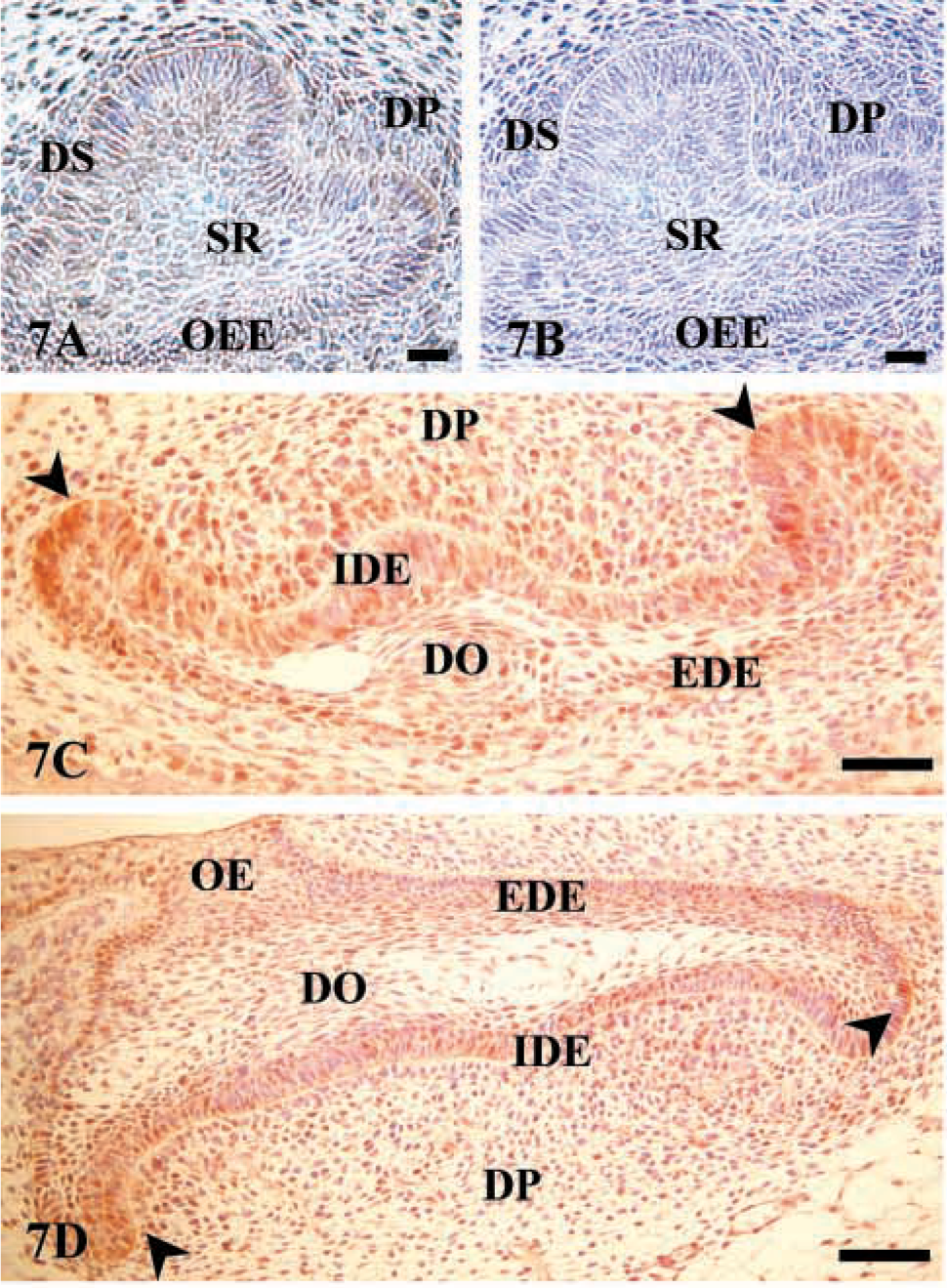
Three principal stages (bud, cap, and bell) are recognized in the formation of teeth (reviewed by Ten Cate 1994). The bud stage, which begins with the incursion of epithelial cells into the mesenchyme of the jaw, is followed by the cap stage, at which time cells condense and the dental papilla and dental follicle are formed. The outer epithelial ingrowth resembles a cap sitting on the dental papilla and is called the dental organ. The dental organ, dental papilla, and dental follicle constitute the tooth germ and give rise to enamel, dentin-pulp, and supporting structures of the tooth, respectively. Continued growth of the tooth germ produces the bell stage named for the resemblance of the dental organ to a bell as the crescent-shaped area of the epithelial cap deepens. At the early bell stage the epithelial cells start to differentiate, forming the internal and external dental epithelia. Immunostaining for AJ18 was weak before the bell stage, at which time AJ18 was expressed primarily in the nuclei of the internal dental epithelium, especially around the cervical loop region of the dental organ (Figure 7). In addition, AJ18 was expressed in the external dental epithelium, which is contiguous with the oral epithelium, and in some of the mesenchymal cells in the dental papilla.
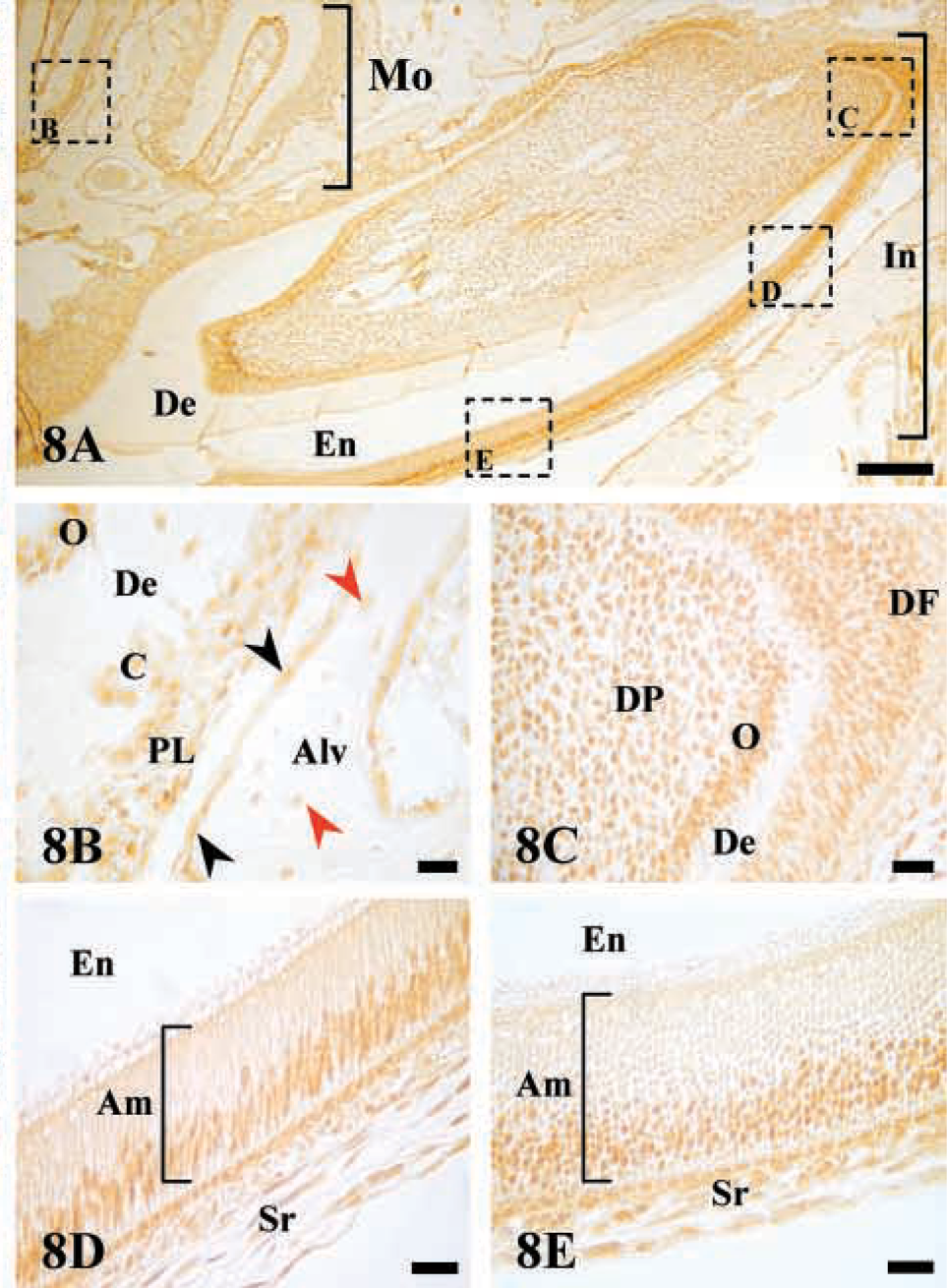
Expression of AJ18 was also studied in the incisors and fully developed molars of a 4-week-old mouse (Figure 8A). In sections through the molar roots, staining for AJ18 was seen in the dental papilla, with strong staining in the odontoblasts lining the dentin. Strong staining was also apparent in the cells of the periodontal ligament and in osteogenic cells lining the surface of the alveolar bone. However, nuclear staining of osteocytes was weak (Figure 8B). In the continuously erupting incisor tooth, a continuum of cell differentiation associated with the formation of dentin and enamel was present only on the labial side of the mouse mandibular incisor. AJ18 staining was observed in the cells of the dental papilla, with strong staining in the newly differentiating odontoblasts (Figure 8C), which lose the staining after the formation of dentin has started. Anterior to this region, the nuclei of ameloblasts with their elongated cell bodies were strongly stained (Figures 8D and 8E). Moreover, the staining intensity appeared to be retained as the ameloblasts progressed from the secretory stage, in which they appear highly ordered (Figure 8D), to the post-maturation stage, in which they undergo apoptosis and the columnar arrangement becomes disorganized (Figure 8E).
Expression in the Brain and Epithelium
Expression of AJ18 mRNA was highest in brain in the embryo and especially after birth, when expression in other tissues declined markedly (Figure 2). However, immunostaining sections of brain in 16 dpc embryos was variable, with the strongest staining in condensing cells that form the future cerebral cortex, midbrain, and tentorium cerebelli (Figures 9A and 9C). In general, staining was not as strong as that seen in the osteogenic cells forming the calvarial bone and was much lower than the staining seen in the epithelium covering the cranium (Figure 9B). Because AJ18 mRNA expression appeared stronger in brain compared to bone, it is possible that protein translation in brain cells is less efficient than in bone cells, or that AJ18 may be highly expressed in specific regions of the brain that were not included in the sections analyzed. Relatively strong staining was observed in skin epithelia (Figure 9B). Although nuclear staining was present in the basal cells of epithelia, staining in the outer layers was diffuse, possibly due to cell death.
Expression in the Kidney, Eye, and Whisker Follicles
Northern blot analyses showed AJ18 mRNA expression in the kidney of mouse embryos at 15 and 17 dpc. Consistent with these results, relatively strong immunostaining for AJ18 was observed in cells within developing glomeruli of the kidney (Figure 10A). In the eye, specific staining was found in cells within the sclera, choroids, and vitreous body, with less staining in the retina (Figure 10C). AJ18 staining was also observed in the epithelial root sheath of developing whisker follicles (Figure 10E). The strong staining of nuclei in the whisker follicles is consistent with the general staining of epithelial cells, including the epidermal cells (Figures 3 and 9), and of epithelial cells lining the intestine and lung alveoli (results not shown).
Discussion
Previous studies have characterized AJ18 as a zinc finger transcription factor that is upregulated in osteogenic cells induced to differentiate with BMP-7. Consistent with the expression of AJ18 mRNA during osteogenic differentiation by FRCCs in vitro (Jheon et al. 2001), AJ18 mRNA was expressed early during the formation of endochondral and intramembranous bones (tibia and calvaria, respectively) in mouse embryos. Expression of AJ18 mRNA was also detected in other tissues and organs, with maximal expression during embryonic development. Whereas significant expression of AJ18 occurred postnatally in brain, which together with mineralized tissues has the highest expression of AJ18, expression was lost in other tissues. As expected for a transcription factor, IHC staining for AJ18 protein was largely restricted to cell nuclei, with the relative intensity of staining in the tissues approximating the level of mRNA determined by Northern blot hybridization.
Comparison of the protein sequences for AJ18 in rat and mouse revealed high conservation of amino acids (97–100%) in the region of the C2H2 zinc fingers and the KRAB domain, signifying the functional importance of these regions in mediating DNA binding and transcriptional activity. In the linker region between the KRAB domain and the zinc fingers, and at the carboxy-terminal region, sequence conservation was much lower (72–79%), indicating that the precise structure of these regions is of lesser functional importance. In previous studies (Jheon et al. 2001) the C2H2 zinc finger motifs of AJ18 has been shown to bind to DNA with a preference for sequences containing 5′-CCACA-3′, which is present within the OSE2 through which Runx2 mediates transcriptional activity (Ducy et al. 1997). AJ18 was found to suppress Runx2-mediated transactivation through the OSE2. Although gene targets for AJ18 have not been identified, forced expression of AJ18 in C3H10T1/2 fibroblast-like cells induced towards osteogenesis by BMP-7 suppressed the expression of alkaline phosphatase, an early marker of osteogenic differentiation.
Although BMPs are characterized by their bone-inductive activity (Rodan and Harada 1997) as members of the TGF-β superfamily of cytokines, they have a broader role in developmental processes involving different tissues and organs (Hogan 1996). Therefore, it is of interest that BMP-7, which was originally used to identify AJ18 (Jheon et al. 2000), has an expression pattern similar to that of AJ18 in mouse embryos. BMP-7 mRNA is expressed in the glomeruli of kidney, in the eye, and in whisker follicles, and its expression is absent in the liver of 17.5 dpc mouse embryos (Helder et al. 1995). Our studies show that the expression of AJ18 in 16 dpc mouse embryos corresponds with the results reported by Helder et al. (1995). In addition, BMP-7 is expressed in the brain, in mineralized tissues such as in hypertrophic chondrocytes, osteoblasts, and periosteum, and in the inner dental epithelium, with expression in a few cells in the dental pulp (Helder et al. 1995). These are all tissues in which AJ18 showed positive staining. In 12–18-day mice, BMP-7 is expressed in all bony tissues and in the odontoblasts and ameloblasts of teeth (Thomadakis et al. 1999). Although AJ18 mRNA expression was markedly reduced in all postnatal tissues, except brain, and was difficult to detect by Northern hybridization, AJ18 protein was detectable by immunostaining in 4-week-old mice. AJ18 was present in the nuclei of chondrocytes and osteoblasts in tibiae, and in the odontoblasts and ameloblasts in molar and incisor teeth. The spatial expression of AJ18 at various developmental stages therefore closely mimics the expression of BMP-7 in mouse tissues, consistent with the original identification of AJ18 as an immediate-early responsive gene to BMP-7 (Jheon et al. 2000). Similar to BMP-7, therefore, AJ18 may modulate the differentiation of bone and cartilage cells, but its expression in various tissues suggests functional roles in the development of other tissues, which may be revealed by generating mice with a targeted disruption of the AJ18 gene. Although significant developmental effects have been observed in BMP-7 knockout mice in only three organs (eyes, kidneys, and bone; Dudley et al. 1995; Luo et al. 1995), other BMPs may compensate for the absence of BMP-7 in the formation of other tissues.
From the IHC analysis of endochondral bone in 16 dpc mouse embryos, it is evident that AJ18 is initially expressed by proliferating chondrocytes and pre-hypertrophic chondrocytes, with sporadic expression in hypertrophic chondrocytes. The formation of cartilage in endochondral bones is controlled by a negative regulatory feedback loop involving PTH-related protein (PTHrP) and Indian hedgehog (Ihh) (Lanske et al. 1996; Vortkamp et al. 1996). Ihh is expressed in the pre-hypertrophic chondrocytes present in the growth plate, where it regulates the rate of hypertrophic chondrocyte differentiation (Vortkamp et al. 1996). Ihh induces the expression of PTHrP in the periarticular perichondrium, which signals to its receptors expressed in pre-hypertrophic chondrocytes and inhibits the expression of Ihh, thereby blocking hypertrophic chondrocyte differentiation (Lanske et al. 1996; Vortkamp et al. 1996). Thus, ablation of the PTH/PTHrP receptor results in mice with accelerated differentiation of chondrocytes within the growth plate of bone (Lanske et al. 1996). The inhibition of Ihh by PTHrP appears to be indirect and through the direct downregulation of BMP-6 (Grimsrud et al. 1999). The spatial expression pattern of AJ18 within the growth plate of bone and the apparent functional redundancy of BMPs suggest that AJ18 may be a possible downstream target for BMP-6 in this cartilage negative feedback loop.
BMP-7 has also been shown to inhibit the terminal differentiation of chondrocytes in the periarticular region, independent of the Ihh/PTHrP/BMP-6 negative feedback loop via unknown inhibitory factors (Haaijman et al. 1999). Periarticular cartilage remains unmineralized, and it is the loss of periarticular cartilage due to new bone formation that leads to joint diseases such as osteoarthritis (Goldring 1999). Therefore, AJ18, which is also expressed in cartilage cells present at the periarticular region, may play a role in inhibition of the terminal differentiation of these chondrocytes under the control of BMP-7. That AJ18 expression is lost in hypertrophic chondrocytes, in which Runx2 expression is induced (Kim et al. 1999), further suggests that AJ18 and Runx2 have antagonistic activities, as evident in the AJ18 suppression of Runx2 transcriptional activity (Jheon et al. 2001). Thus, whereas the overexpression of Runx2 in pre-hypertrophic chondrocytes induces differentiation to hypertrophic chondrocytes (Takeda et al. 2001), the overexpression of AJ18 in pre-hypertrophic chondrocytes may inhibit hypertrophic chondrocyte differentiation. Therefore, it would be interesting to determine the phenotypic effects of AJ18 overexpression in a chondrogenic cell line such as CFK2 (Bernier and Goltzman 1993), along with its effects on Runx2 and Ihh expression.
In summary, these studies show that the expression profile of AJ18 in embryonic tissues is consistent with the concept that AJ18 acts downstream of BMP-7 during tissue morphogenesis.
Footnotes
Acknowledgements
Supported by a grant (MOP) from the Canadian Institutes of Health Research (CIHR). Andrew Jheon was the recipient of an Ontario Graduate Student Science and Technology (OGSST) scholarship.