
Abstract
During the initiation of an immune response, antigen-presenting cells employ MHC class II antigens as key molecules to present small peptides to CD4-positive lymphocytes. The invariant chain (Ii; CD74) plays a critical role in this process by influencing the expression and peptide loading of the MHC class II molecules. Therefore, coordinate expression of these molecules is believed to play an important role in antigen presentation. This study explores the expression of these molecules in fetal tissues. Formalin-fixed, paraffin-embedded multi-organ tissue blocks from aborted fetuses (age range 7–22 weeks) were immunostained for Ii/CD74 and MHC class II antigens using commercially available monoclonal antibodies for Ii/CD74 (LN2) and MHC class II antigens (LN3), respectively. Coordinate staining for Ii/CD74 and MHC class II antigens was seen in the skin, proximal renal tubules, tips of small intestinal mucosa, and cells of the reticuloendothelial system, including the spleen and thymus. Expression of Ii/CD74, but not of MHC class II antigens, was seen in pulmonary alveolar epithelium in all cases and in testicular Leydig cells (11 of 11 testes examined). The distribution and intensity of staining did not change significantly with age. In conclusion, this study describes distribution of Ii/CD74 and MHC class II antigens in human fetal tissues. Coordinate expression of Ii/CD74 and MHC class II antigens was identified in most fetal tissues, but there were also notable exceptions. In all cases this took the form of expression of Ii/CD74 in the absence of MHC class II expression. Discordance was particularly striking in pulmonary alveolar epithelium and testicular Leydig cells. This suggests that the Ii/CD74 molecule has functional roles in addition to its role in antigen presentation.
M
After synthesis, MHC II proteins undergo folding in the endoplasmic reticulum before transport to the endosomal vesicles, where binding to the antigenic peptides takes place (cf. Cresswell 1994; Jensen et al. 1999b). The stabilization of the newly synthesized MHC II heterodimers is achieved through initial assembly with a chaperone protein called the invariant chain (Ii), a non-polymorphic Type II integral membrane glycopeptide molecule (Jensen et al. 1999a,b). Through the Leukocyte Typing workshops, Ii has been designated as CD74 (Moller et al. 1995).
Ii/CD74 is synthesized intracellularly in parallel with MHC class II synthesis, although the genes that encode them are located on different chromosomes. Ii/CD74 contains an unstructured region, the class II-associated Ii peptide (CLIP) that acts as a surrogate peptide and interacts directly with the peptide-binding site of the MHC II molecule (Ghosh et al. 1995). Ii/CD74 is also believed to serve additional functions in the MHC II processing pathway, such as targeting the MHC proteins to the endosomal compartment (Ceman and Sant 1995). In the absence of Ii/CD74, MHC II molecules may associate with the other proteins present in the endoplasmic reticulum, which could result in the presentation of self-antigens (Roche and Cresswell 1990; Teyton et al. 1990; Busch et al. 1996). Ii/CD74 is very sensitive to proteases and rapidly undergoes a series of proteolytic cleavages in the hydrolytic environment in the endosomal pathway (Blum and Creswell, 1988; Riese et al. 1996; Riese and Chapman 2000). The CLIP segment is protected from the proteases because it is topologically embedded in the MHC peptide-binding site (Ghosh et al. 1995). Peptide exchange reactions characterized by the dissociation of CLIP and the binding of heterogeneous population of peptides follow, before presentation of the peptide on the cell surface.
The complex interdependence of MHC class II molecules and Ii/CD74 antigens throughout processing is further reflected in their concordant regulation and expression in cell lines and in most adult tissues (Collins et al. 1984; Quaranta et al. 1984; Volc-Platzer et al. 1984; Levine et al. 1985; Symington et al. 1985; Momburg et al. 1986; Brown et al. 1991; Chang and Flavell 1995; Henne et al. 1995; Saleem et al. 1997). One of us has previously demonstrated coordinate expression of invariant chain and MHC class II antigens in adult lungs (n = 136), with positive immunoreactivity in both normal pulmonary epithelium and non-small-cell lung carcinomas (Hua et al. 1998).
In spite of their implications for development of immunological competence, expression of these antigens in human development has not been systematically studied. Results from such studies may add to understanding of the development of the antigen-presentation machinery in fetal tissues and could suggest additional functions for Ii or MHC II proteins by, e.g., revealing discordant expression patterns. Therefore, the concordant expression of the two molecules is a reflection of their underlying interdependent involvement in antigen presentation, whereas discordant expression in some cell types would point to additional independent functions of one or the other of the molecules. We undertook a study of the expression of these two molecules in tissues of fetuses of various ages ranging from 7 to 22 weeks estimated gestational age.
Materials and Methods
Human fetal tissues were obtained, after Institutional Review Board approval, from 23 autopsied fetuses (age range 7–22 weeks estimated gestational age), with the majority of the cases being from the second trimester. These fetuses were the subjects of surgical pathology review and special care had been taken with the diagnostic tissue blocks to include representative tissues from several organs from each fetus, sampled by a 3-mm-diameter skin punch. Organs sampled included the liver, kidney, pancreas, heart, skeletal muscle, lung, esophagus, stomach, small and large intestines, bone/cartilage, skin, testis, ovary, thymus, spleen, adrenal, and thyroid. The brain and spinal cord from these fetuses had been harvested for another study, and therefore samples from these organs could not be obtained. Multiple tissue samples (10–15) from an individual were placed into one cassette and processed according to standard protocol to generate paraffin blocks.
More than one of the following was used to estimate the fetal age: maternal ultrasound, last menstrual date, and fetal measurements such as crown-rump length, foot length, and fetal weight.
Monoclonal antibodies to Ii/CD74 (LN2) or MHC class II antigens (LN3) were purchased from Dako (Carpinteria, CA). The immunohistochemical assay used the avidinbiotin-peroxidase (ABC) method on serial sections. Briefly, 4-μm-thick paraffin sections, after deparaffinization and blocking of endogenous peroxidase activity, were subjected to microwave antigen retrieval for 10 minutes in a citrate buffer, pH 6.0. After this the sections were incubated with anti-LN2 or anti-LN3 for 12 hr. After subsequent incubations with biotinylated anti-mouse Ig and avidin-labeled peroxidase, reactions were visualized using diaminobenzidine (DAB) as the substrate, followed by counterstaining with hematoxylin. Normal tonsils were used as positive controls. The specificity of staining was ascertained by (a) staining in the absence of primary antibody (negative control) and (b) staining with a number of additional antibodies with different specificity, including anti-α-1-microglobulin (Lögdberg et al. 2000), Das-1 (Badve et al. 2000), anti-CD34, anti-RBP, and others (unpublished data).
Semiquantitative evaluation of staining intensity (0, absent; +, weak; ++, medium; or +++, strong) was performed by independent reading of the slides by all three authors, followed by collective evaluation in a multiheaded microscope.
Results
General Tissue Distribution of LN2 and LN3
In our material overall, staining for LN2 was more widespread than staining for LN3, confined not only to cells of the macrophage lineage but also apparent in epithelial cells. In fetal whole mount (EGA 7 weeks), both LN2 and LN3 staining was present in scattered cells in all the organs examined. Stained cells were either within hematolymphopoietic organs or existed as scattered single cells within the interstitium of organ parenchyma and soft tissue, and persisted throughout the fetal period studied. Representative examples of these tissues, at ∼ Week 21 of EGA, are presented in Figures 1–3. As might be expected, cytoplasmic staining was seen in APCs and in epithelial cells of endodermal and mesodermal origin. The pattern of expression of either antigen did not vary significantly with fetal age in most organs, with a few exceptions, notably in the kidney (Table 1). Staining patterns by organ system are described below.
Cardiovascular System. Scattered cells positive for LN2 and LN3 were detected within the myocardium. These were judged most likely to be tissue histocytes. There was no significant staining of the cardiac myocytes, endocardium or pericardium, or of the endothelia of small blood vessels or capillaries in the heart.
Respiratory System. The bronchial epithelium showed almost no reactivity to either antibody at 7 weeks of gestation. Expression did occur, however, in fetuses aged 14 weeks and older, and followed a similar pattern to that described for the pulmonary interstitium (see below), with non-epithelial cells reactive for both antibodies. Staining was also detected in some tracheal and laryngeal epithelial cells. These positive-staining epithelial cells were morphologically similar to those that did not express LN2 or LN3. There were greater numbers of cells staining for LN2 than LN3 at all ages. Lung parenchyma was poorly represented in the sections from the 7-week fetus.
Strong staining of the scattered single non-epithelial cells in the pulmonary interstitium with both LN2 and LN3 was seen at all ages (Figure 1B and 1D). These cells, presumed mostly to be tissue macrophages, increased in number with age. Pleural epithelial cells and subpleural fibroblasts did not stain with either antibody, nor did the endothelium of the pulmonary blood vessels show expression of target proteins.
LN2 Reactivity. In older fetuses, the lung acinar (alveolar) epithelium stained for LN2 in part or whole acini by the age of 12–14 weeks (Figure 1B). There was no or only minimal staining of the smaller bronchioles/alveolar ducts.
LN3 Reactivity. LN3 reactivity was restricted to the non-epithelial cells in the pulmonary interstitium.
Gastrointestinal System
Intestines. LN2 and LN3 positive cells, predominantly in the lamina propria and submucosa, were present in the intestinal stroma as early as at 7 weeks of gestation. With increasing age there was a marked increase in positive-staining cells in the lamina propria, particularly in the small and large intestines (Figure 2A and 2C). These cells were located immediately beneath the basement membrane of the surface epithelium, giving a wreath-like appearance on cross-section. Rare scattered LN2- and LN3-positive cells were observed within the muscularis propria and in the region of myenteric plexuses.
LN2 Staining. Weak expression of LN2 was seen in the gastric mucosa and focally in small intestinal surface epithelium in the older fetuses. There was no staining of the endothelial cells in most locales with either antibody. Expression of LN2 but not LN3 was noted in the mesenteric and adventitial vessels of the small bowel (Figure 2A and 2C).
LN3 Staining. No staining for LN3 of the surface epithelium or glands was noted at any age.
Liver and Pancreas. Staining of Kupffer cells was seen at 7 weeks of gestation by both LN2 and LN3. There was no staining of the hepatocytes or biliary tree at any age. In the pancreas, rare scattered stromal cells that were reactive with LN2 and LN3 were noted from very early stages. The number of these cells increased with age. No staining of the pancreatic duct lining cells was seen. Endothelial reactivity within few of the pancreatic blood vessels was noted in the late second-trimester fetuses with LN2 and in some cases also with LN3.
LN2 Staining. At older ages and with the development of biliary radicals, clusters of LN2-positive cells were seen in the portal tracts (Figure 3A). LN2 expression was noted in both islet and parenchymal cells of the fetal pancreas. This took the form of a dot-like supranuclear positivity, with the rest of the cytoplasm being devoid of any staining (Figure 2B). A diffuse cytoplasmic staining of the acinar cells was seen.
LN3 Staining. Although LN3 reactivity was more or less restricted to the scattered stromal cells described above, in an occasional case weak diffuse staining of the islet cells was seen.
Genitourinary System
Urinary Tract. At 7 weeks of gestation, scattered cells positive for both LN2 and LN3 were identified in the renal interstitium and in the glomerular mesangium. At all ages, more LN2- than LN3-positive interstitial cells were observed. The number of cells reacting with either antibody increased with age and followed the pattern of Ii/CD74 expression followed by MHC II expression. The ureters and urinary bladder were not represented in the samples examined.
LN2 Reactivity. By 10–14 weeks of gestation, additional staining for LN2 was noted predominantly in the proximal convoluted tubules (Figure 1A), small tubules in the deep medulla, and endothelia of small blood vessels. Tissues from some of the older fetuses that included collecting ducts were reactive with LN2 and only infrequently with LN3.
LN3 Reactivity. The medullary tubules did not react with LN3 at any age.
Genital Tract. No significant staining of the ovaries with either antigen was seen. However, the generally poor quality of this material precluded further evaluation of these tissues.
LN2 Reactivity. In the testes, a dual pattern of staining was revealed at all ages. One population of cells stained strongly (3+ intensity), was interstitial in location, and had the morphology of tissue macrophages. The second population consisted of Leydig cells, which stained with variable intensity. In the seminiferous tubules no staining was apparent in either the germ cells or the Sertoli cells. By comparison, the rete testis and the epididymis showed strong expression of LN2 (Figure 1E and 1F).
LN 3 Reactivity. No staining was seen of Leydig cells, rete testis, or epididymis (Figure 1G and 1H). As expected, the interstitial macrophages stained positive for LN3.
Endocrine System. There was no staining of fetal thyroid or parathyroid glands, with the exception of interstitial cells staining with both LN2 and LN3. The adrenal gland showed prominent diffuse cytoplasmic staining (LN2>LN3) of the zona fasciculata and scattered interstitial cells. The medulla showed no significant staining in the cases examined.
Reticuloendothelial System. Staining for both LN2 and LN3 was seen in cells belonging to the macrophage lineage and within the thymus. Intense staining of almost all the cells was noted in the thymic medulla. In the thymic cortex (Figure 3C), positive-staining cells were more scattered and were intermixed with unstained lymphoid cells. High-intensity staining also was seen in the spleen (Figure 3D) and lymphoid tissues.
Skin and Musculoskeletal Systems. LN2 and LN3 reactivity was present in scattered cells within the dermis as early as 7 weeks of gestational age (Figure 3B). With maturation of the fetus, stained cells increased in number and in staining intensity. Staining within the epidermis was not detected at 7 weeks. Basally located dendritic cells were detected at around 10–12 weeks of age by virtue of their reactivity with both the antibodies. No significant staining was seen in other epidermal cells or in adnexal structures. Scattered LN2- and/or LN3-positive cells were detected in the subcutaneous tissues and muscle at all ages. No staining was apparent in cartilage.
Discussion
LN2 and LN3 are monoclonal antibodies to Ii/CD74 and a non-polymorphic portion of the HLA-DR α-chain (human MHC class II antigen), respectively. Using these antibodies and immunohistochemical methods, we characterized the expression of MHC II and Ii/CD74 antigens in fetal tissues from estimated gestational ages of 7–22 weeks. We found expression of these antigens not only in APCs, but also in fetal epithelial cells. This suggests that induction, either by inflammatory or neoplastic processes, is not necessary for MHC expression in epithelial cells. The expression of MHC II and Ii/CD74 was concordant in most of the tissues studied. However, in some organs (lung, kidney, testis, and pancreas), we noted discordant expression (see Table 2). In these latter tissues, Ii/CD74 was expressed in non-immune cells in the absence of MHC II expression. In some tissues, such as the kidney, more interstitial cells expressed Ii/CD74 than MHC II at all ages.
On examination of serial sections, it appeared that all cells expressing MHC II also express Ii/CD74 (data not shown). It has been previously suggested that expression of at least one of the human MHC class II isotypes (DR, DP, DQ) might be essential for the full constitutive expression of Ii/CD74. Henne et al. (1995) have shown that Ii/CD 74 can be expressed in cell lines in the absence of class II products. Our finding of greater numbers of macrophages, epithelial cells, and endothelial cells expressing Ii/CD74 in the absence of MHC II (HLA-DR) supports these in vitro results. One implication of this is the possibility that mechanisms for trafficking Ii/CD74 to the cell surface exist in development that are independent of MHC II transport.
Saleem et al. (1997) demonstrated discordant expression of MHC II and Ii/CD74 in the rat kidney. In this model, however, MHC II expression was observed in the absence of Ii/CD74 expression. These authors suggest that the MHC II-positive phenotype of the interstitial cells of the kidney might serve a key role in peripheral T-cell tolerance. These observations are at variance with those of Momburg and Moller (1988) who systemically analyzed the tissue distribution of MHC II and Ii/CD74 in the rat without noting MHC class II expression in the absence of Ii/CD74 expression. Similarly, we did not observe the MHC class II-only phenotype in any of the human fetal tissues studied, although the intensity of expression of MHC class II was greater than that of Ii/CD74 in renal endothelial cells (Figure 1A and 1C). It is possible that differences in the preservation of the epitopes recognized by different antibodies may underlie such differences in the detectable patterns of expression. Again, in our experience, the expression of MHC II was always associated with Ii/CD74 expression, which ontogenically preceded it. A caveat that needs to be borne in mind is that none of the studies used dual staining methodologies for identification of MHC II and Ii/CD74 expression.
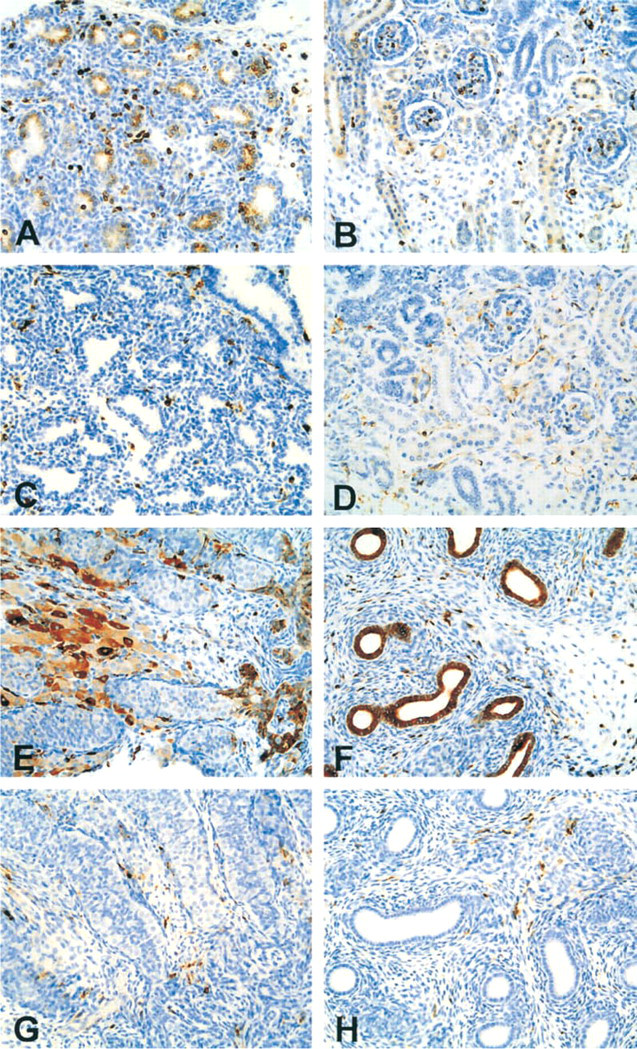
LN2 and LN3 expression in human fetus (∼21 weeks). Low-magnification photographs of LN-2 (
Age-related changes in expression of MHC class II (LN3) and invariant chain (LN2)a
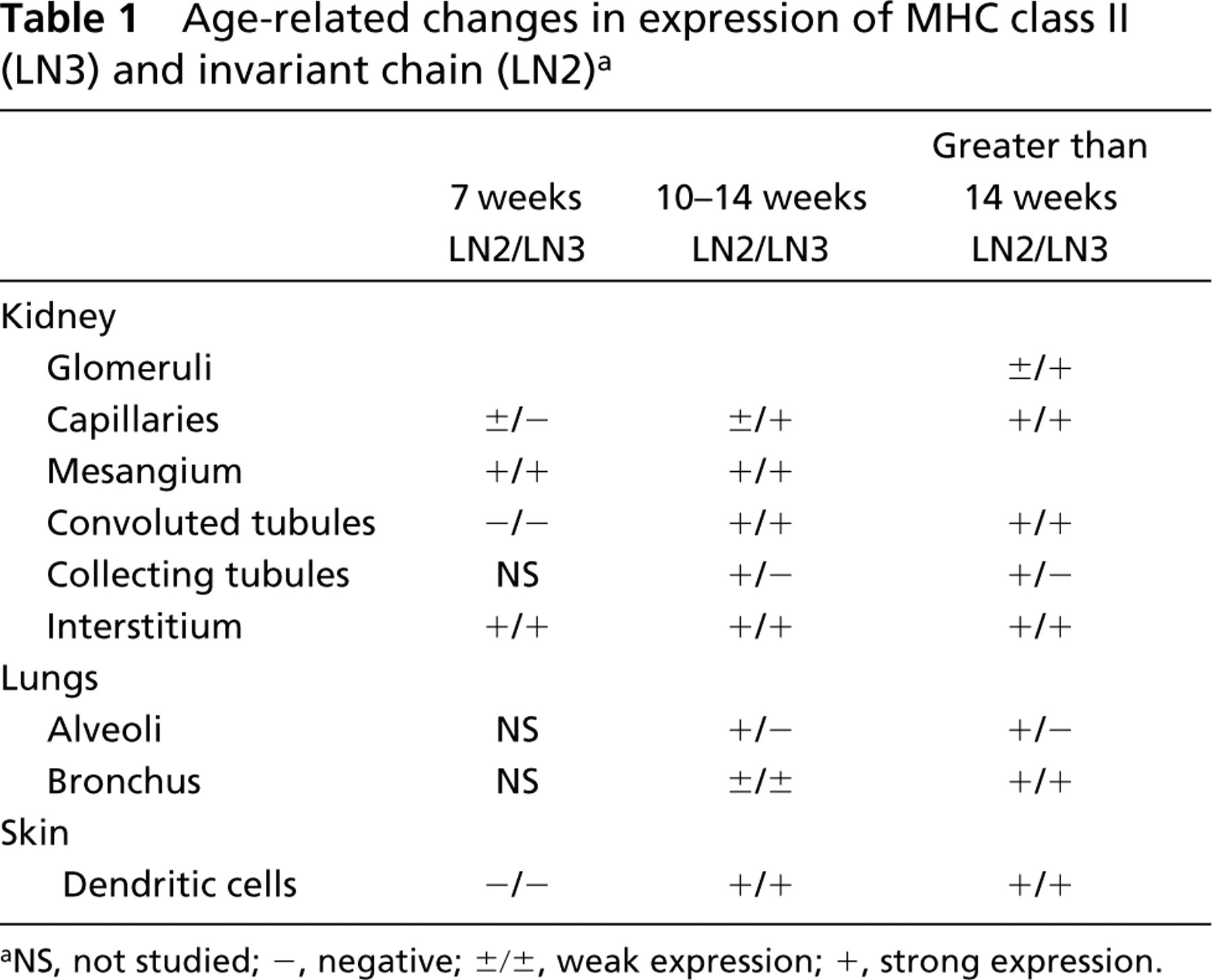
NS, not studied; -, negative; ±/±, weak expression; +, strong expression.
The pattern of expression of Ii/CD74 within the pulmonary epithelium was also interesting and to our knowledge has not been previously described. Ii/CD74 expression was observed in cells lining the developing pulmonary acini (Figure 1b) but not in the bronchioles and air ducts. This pattern of expression suggests a role for Ii/CD74 in the development and maturation of the alveolar epithelium. Expression of MHC II and Ii/CD74 is seen in normal pulmonary epithelium and in non-small-cell lung carcinomas (Ioachim et al. 1996; Hua et al. 1998) and is extremely rare in small-cell carcinomas of the lung. In the non-small-cell tumors, adenocarcinomas more than squamous cell carcinomas were likely to express MHC II and Ii/CD74 (Hua et al. 1998). A correlation with grade in these tumors could not be demonstrated (Hua et al. 1998).
Site of discordant expression of MHC class II (LN3) and invariant chain (LN2)

In adult pulmonary epithelium, previous studies by one of us (Hua et al. 1998) revealed concordant expression of Ii/CD74 and MHC II. In contrast, in the fetal material studied here, Ii/CD74 but not MHC II expression is apparent in pulmonary epithelium. In fact, this pattern of expression was seen in lung tissues of fetuses at all ages examined. The concordant expression of Ii/CD74 and MHC II in the interstitial APCs served as an internal control in this organ. Peters et al. (1990) noted a similar absence of expression of MHC II antigens in fetuses less than 21 weeks of age. In fetuses older than 21 weeks, these investigators found a significant correlation between lung inflammation and MHC II expression. Other factors, in addition to inflammation, that might possibly influence MHC II expression in lung include maturation and exposure to environmental antigens as a result of respiration on lungs. To differentiate the impact of these factors, lung tissues from third-trimester fetuses and stillbirths should be compared with those from autopsies in premature infants. In preliminary studies (to be reported elsewhere), we observed a trend for concordant expression of Ii/CD74 and MHC II in pulmonary epithelium during the late third trimester.
Expression of MHC class II was also noted in rare scattered cells of the bronchial epithelium. These cells, akin to those described by Peters et al. (1990) and Dammrich et al. (1990), were morphologically similar to ones that did not express MHC class II. Staining for Ii/CD74 was much more abundant than MHC class II in the bronchial epithelium. Expression was also noted in the tracheal and laryngeal surface epithelium and glands, indicating a constitutive expression of MHC II and Ii/CD74 in epithelial cells.
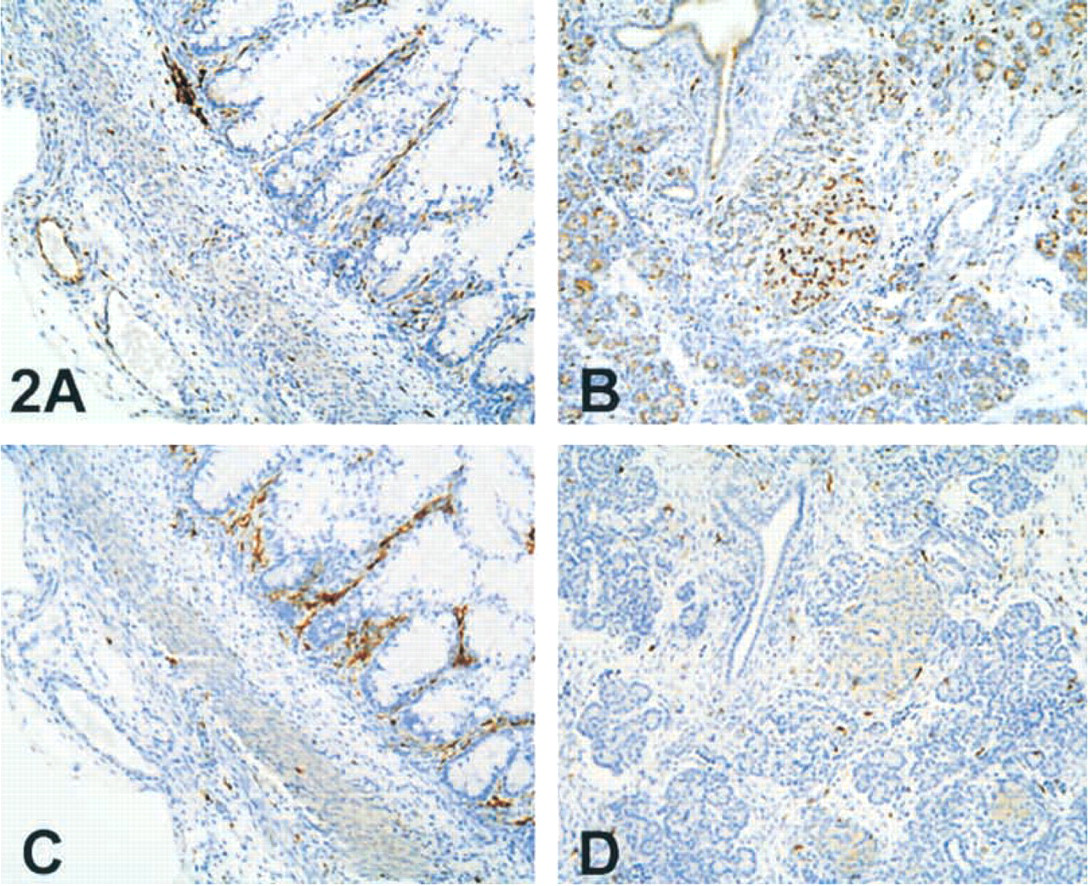
LN2 and LN3 expression in human fetus (∼21 weeks). Low-magnification photographs of intestine (
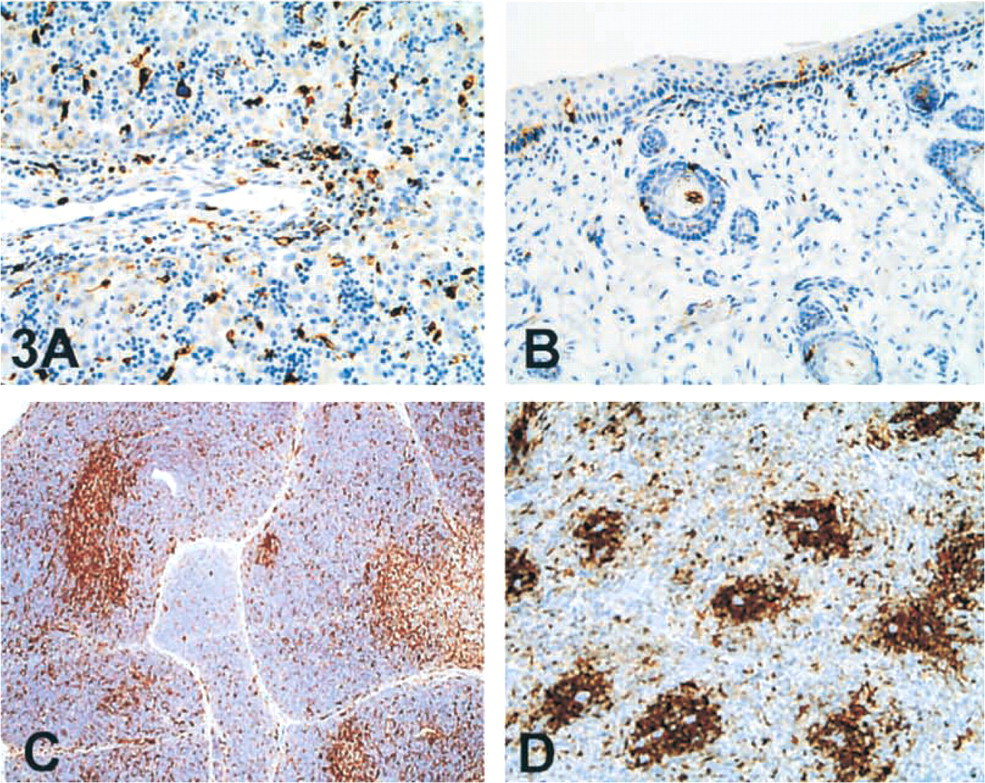
LN2 and LN-3 expression in human fetus (∼21 weeks). Medium magnification photographs of liver (
Within the intestines, expression of Ii/CD74 and MHC II was seen predominantly in the APCs in the lamina propria (Figure 2A and 2C). Surface epithelial cells of the small intestines were only weakly reactive for Ii/CD74, and no staining for MHC II was seen in these cells. This is in contrast to the findings of Vidal et al. (1993), who observed that the mouse enterocytes were MHC II +, Ii/CD74-. In human fetuses, Mac-Donald et al. (1988), and Oliver et al. (1988) have found faint and inconsistent expression of MHC II and Ii/CD74 at the tips of the intestinal villi at as early as 18 weeks of gestation. It has been suggested that prostaglandins, which were used to induce abortions, might have induced this expression (Rognum et al. 1992). Induction of MHC II and Ii/CD74 expression in epithelial cells is known to occur after local production or injection of interferon-γ (Kvale et al. 1988) in inflammation, autoimmune diseases, and allograft rejection. In a study of intestinal tissues from premature infants, Rognum et al (1992) have shown that expression of MHC class II antigens (HLA-DR) was present only 1 week after birth, around the same time as IgA-secreting cells emerge. They attribute these findings to postnatal environmental factors. However, this group did not study expression pattern of invariant chain. Similar studies in mice have shown marked increases in Ii/CD74 (Ii31, Ii41) and MHC class II (I-A β) mRNA after weaning (Sanderson et al. 1992,1993). It therefore appears that although expression of MHC II and Ii/CD74 is absent in the fetus, it can be induced both postnatally as well as intranatally.
In the fetal intestine, staining of the endothelial cells for Ii/CD74 but not for MHC II was noted (Figure 2A and 2C). In contrast, Momburg et al. (1988) found that HLA-DR+ endothelial cells in normal adult intestines were Ii/CD74-, and that in inflamed intestine (Crohn's disease) HLA-DR expression in venous/venular and capillary endothelium greatly exceeded Ii/CD74 expression. These investigators also found a considerable degree of inappropriate expression of MHC class II antigens and Ii/CD74 in epithelial cells, vascular endothelial cells, and Schwann cells of the enteric nerve plexus, and this expression correlated with the local inflammatory cell density. It should be noted that, within the colon, altered expression of MHC and Ii/CD74 has been noted in the vicinity of tumors (Degener et al. 1988). A significant number of tumors also exhibit non-coordinate expression with greater amounts of Ii/CD74 than MHC II (Degener et al. 1988; Moller et al. 1991; Jiang et al. 1999). Jiang et al. (1999) have demonstrated an association of increased Ii/CD74 expression and poor-grade and intratumoral lymphocytes. On the basis of this finding they suggest that excess Ii/CD74 expression might render the tumor less immunogenic by inhibiting antigen presentation. This putative link between cancer and immunogenicity, however, was not corroborated by Moller et al. (1991), who failed to find a correlation of MHC II and Ii/CD74 expression with recurrence or survival in colon carcinoma.
As for endocrine organs, our findings are similar to those reported by Oliver et al. (1988) in a previous fetal study. Like us, they reported the presence of scattered MHC II and Ii/CD74 positivity within pancreatic interstitial and parenchymal cells. Likewise, our findings in the thyroid and adrenal glands are similar to those previously described by these workers. In both studies, the endothelial cells in the adrenal were negative for MHC II and Ii/CD74, in direct contrast to the pattern of expression described in endothelial cells from adult tissues (Alejandro et al. 1982).
Our finding of discordant expression of MHC class II and Ii/CD74 in Leydig cells of the testis and in epididymis (Figures 1E-1H) has not been previously reported. In the adult, Haas et al. (1988) described the absence of MHC class I and II antigens in developing testicular germ cells, including spermatozoa MHC class II antigens were not found on any cells within the seminiferous tubules. However, the class II antigen was identified on dendrite-like cells between the seminiferous tubules and on vessel endothelium. Consistent with our results, Haas et al. (1988) did not find MHC antigen expression in Sertoli cells and germ cells. Wilkins et al. (1992) found that MHC class II and I were expressed by most stromal cells but not by seminiferous tubules in testes from pediatric subjects. Monoclonal antibodies against MHC class I antigens have been shown to stimulate testosterone production in mouse Leydig cells with the same potency as LH (Solano et al. 1988). These authors suggest that bound hormone triggers an association of the MHC class I antigen with the LH/hCG receptor, resulting in activation of the target cell. It is hypothesized (Hedger et al. 1990) that an “immunosuppressive” mechanism, principally mediated by non-steroidal factors secreted by the Leydig cells, exists in the gonads to prevent activation of the immune system by germ cell antigens and growth factors associated with germ cell proliferation and differentiation. This mechanism probably acts in parallel with normal antigen-specific tolerance mechanisms operating at the gonadal level. Because immune responses to germ cells are believed to be a significant causative factor in infertility, the role of Leydig cells in local immune responses needs to be further defined.
Expression of Ii/CD74 in the Leydig cells of the testis and in the cells lining the epididymis was an unexpected and previously unreported finding. The only other study that studied expression of Ii/CD74 (Ouellette et al. 1991) did not find mRNA for Ii/CD74 in adult mouse testes using cDNA probes.
In conclusion, although MHC II antigens and Ii/CD74 are generally concordantly expressed on cells of the monocyte/RES lineage in most tissues in fetal as well as adult life, reflecting their interdependent role in antigen presentation, several examples of tissue demonstrating discordant expression were seen in this study of fetal material. The lack of concordance was particularly prominent in fetal lung epithelium, epididymis, and on testicular Leydig cells, but also was noted on cell types in other tissues, such as renal tubule cells, pancreatic islet cells, bronchial and intestinal surface epithelium, and mesenteric vascular endothelium. This non-concordance always took the form of Ii/CD74 expression in the absence of MHC class II expression. It is unlikely that non-classical MHC class II antigens are playing a role as alternative partners to Ii/CD74 in some early fetal tissues, because it has been shown, at least in the bowel, that HLA-DR are the first of the class II antigens to be expressed (Ghosh et al. 1986; Momburg et al. 1988). This suggests functional roles for the Ii/CD74 molecule other than those established for antigen presentation in the context of MHC class II antigens. Possibly Ii/CD74 may act as chaperone in the expression of other molecules than MHC class II antigens during fetal life.
Footnotes
Acknowledgements
We wish to thank Linda Lögdberg for helping with the preparation of the manuscript.