
Abstract
An improved thioflavin-S stain, Gallyas silver stain, and two immunostainings were quantitatively compared for demonstration of neurofibrillary tangles (NFTs) on the same sections. Sections of hippocampal formation from seven cases of Alzheimer's disease (AD) were immunofluorescently stained with a commercially available polyclonal NFT antibody or a PHF-1 monoclonal antibody, followed by an improved thioflavin-S stain, and finally by Gallyas silver staining. The thioflavin-S method was improved by using a combination quenching method that removes background autofluorescence without remarkable tissue damage and by post-treatment with concentrated phosphate buffer, which minimizes photobleaching. PHF-1 or NFT immunostaining is much less sensitive than the improved thioflavin-S staining and Gallyas silver staining, particularly in the transentorhinal region. Moreover PHF-1 immunoreactivity varied greatly among AD individuals. Thioflavin-S staining and Gallyas silver staining show almost the same sensitivity in NFT demonstration, but only the former depends on the secondary protein structure of NFTs. This study suggests that the improved thioflavin-S staining is a simple, sensitive, and consistent method for demonstration of neurofibrillary pathology.
Keywords
N
Comparisons of the various staining methods for NFT demonstration had been reported in several earlier studies (Lamy et al. 1989; Vallet et al. 1992; Cullen et al. 1996). The commonly reported drawbacks associated with the available methods include nonspecificity, stain variability, tissue damage, and amount of time required. In this study we introduce a modification of the thioflavin-S histochemical technique that addresses many of these practical concerns. The present study compares the modified thioflavin-S staining to conventional thioflavin-S staining, Gallyas silver staining, and anti-NFT or anti-tau immunofluo-rescence. Gallyas silver staining was chosen as a representative of the available silver staining techniques because it shows much less nonspecific staining on normal cellular elements and is generally considered a relatively specific stain for neurofibrillary pathology. Because the past methodological comparison studies had been performed on different sections, the present study may permit a more accurate comparison of sensitivity and selectivity by testing three staining methods on the same sections.
Materials and Methods
Tissues
Postmortem brain tissues were obtained from 7 AD patients autopsied at the University of Kentucky Alzheimer's Disease Research Center (ADRC). Age at death of AD patients was 79.2 ± 8.7 years (range 65–91) and that of controls was 79.9 ± 9.1 years (range 66–95). The mean postmortem intervals were 2.8 hr (range 2.7–3 hr) for AD brains and 2.9 hr (range 1.8–4.5 hr) for controls. All AD subjects met the clinical and neuropathological criteria for the diagnosis of AD. Control subjects were individuals without evidence of neurological disorders who had been followed longitudinally as part of the normal volunteer control group at the University of Kentucky ADRC. Paraffin-embedded blocks were prepared by sequential dehydration in graded ethanol and vacuum-infiltration in paraffin before embedding and serial sectioning to a thickness of 8 μm.
Antigen Retrieval and Quenching Autofluorescence
Deparaffinized brain sections were rehydrated in distilled water and placed in a stainless steel pressure cooker containing a boiling dilution (1:100) of Antigen Unmasking Solution (#H-3300; Vector Laboratories, Burlingame, CA) according to the manufacturer's protocol. After heating under pressure for 1 min, samples were removed, placed briefly in a room-temperature (RT) water bath, and washed in PBS pH 7.2, for 5 min.
Because the human brain sections showed considerable autofluorescence at FITC and Cy3 channels because of the abundance of highly fluorescent lipofuscin granules, a combination quenching procedure was used to reduce background autofluorescence (Sun et al. 2001). First, slides were incubated in 0.3% KMnO4 for 3–5 min, washed with water, then treated with a solution of 1% K2S2O5 and 1% oxalic acid until the brown color was removed from the tissue (typically 20–40 sec). After a second wash with water, 1% sodium borohydride (NaBH4), which was prepared 2 hr before use, was applied for 5 min. Samples were then washed three times with distilled water and returned to PBS.
Immunofluorescent Staining
After antigen retrieval and quenching treatment, the deparaffinized sections were subjected to conventional immunofluorescence staining using either rabbit anti-NFT polyclonal antibody (#AB1518; Chemicon, Temecula, CA) or mouse PHF-1 monoclonal antibody, which recognizes the dually phosphorylated Ser396 and Ser404 epitope of tau peptides (Otvos et al. 1994). The PHF-1 antibody was kindly provided by Dr. Peter Davies (Albert Einstein College of Medicine; Bronx, NY). Incubation with anti-NFT antibody (1:400) or PHF-1 antibody (1:60) took place overnight at 4C, followed by 1 hr at room temperature (RT). One control section was processed identically except that the primary antibody was omitted from the incubation buffer. After rinsing, sections were incubated in the presence of Cy3-conjugated goat, anti-rabbit secondary antibody (1:200) or goat anti-mouse antibody (1:100) (Sigma; St. Louis, MO) for 3 hr at RT. Sections were then thoroughly rinsed in PBS. For the same-section comparisons in this study, this step was immediately followed by the thioflavin-S staining procedure.
Improved Thioflavin-S Staining
Traditional thioflavin-S staining was previously modified by Guntern et al. (1992) by adding pretreatments with KMnO4 for 20 min followed by NaOH plus H2O2 treatment. However, NaOH treatment causes tissue damage in some cases, as reported by others (Cullen et al. 1996) and in our preliminary study. Photobleaching is also encountered in the modified method above (Vallet et al. 1992), possibly due to the quenching treatment or/and the low thioflavin-S concentration used. In this study we introduce two modifications to the thioflavin-S staining method based on our empirical observations of staining quality. First, the quenching method was improved, as described above, by reducing the duration of potassium permanganate oxidation from 20 min (Vallet et al. 1992) to 4 min, followed by 1% sodium borohydride. Treatments with NaOH, H2O2, and acetic acid were abolished. Second, sections were post-treated with a high concentration of phosphate buffer for 30 min at 4C, which was found to alleviate photobleaching. In addition, because of the high background associated with the intrinsic blue fluorescence from tissue proteins under the DAPI filter set, we used in this study the FITC filter set (ex 470 ± 20 nm, em >515 nm).
In our same-section comparison study of multiple staining methods, the thioflavin-S staining was preceded by the combination quenching steps (described above) and conventional immunofluorescent staining on deparaffinized sections. Sections were then stained with 0.05% thioflavin-S in 50% ethanol in the dark for 8 min. In preliminary optimization trials, dilutions of 0.01%, 0.05%, 0.1%, and 1% thioflavin-S solutions were tested for optimal staining quality at 8 min. A thioflavin-S concentration of 0.05% was found to be optimal and was used for the remainder of the study. This step was followed by differentiation in two changes of 80% ethanol for 10 sec each and three washes in large volumes of distilled water. Slides were then incubated in a high concentration of phosphate buffer (411 mM NaCl, 8.1 mM KCl, 30 mM Na2HPO4, 5.2 mM KH2PO4, pH 7.2) at 4C for 30 min or more, then briefly rinsed with distilled water and coverslipped using water. No mounting resin was used so as to facilitate re-use of the same sections for subsequent stains. Images at different subregions were captured with a digital camera, and NFTs were counted (see below) before any subsequent silver staining procedure.
Fluorescence Microscopy
The induced fluorescence, silver staining, and immunostaining were viewed using an Axioplan 2 imaging microscope (Zeiss; Oberkochen, Germany). For fluorescence, the filters used were as follows: rhodamine (ex 546 ± 6 nm, em > 570 nm); FITC (ex 470 ± 20 nm, em >515 nm); DAPI (ex 365 ± 6 nm, em >397 nm). Images were captured using an Axiocam digital camera interfaced with a computer containing Axiovision 3.0 software (Zeiss). Inputs from rhodamine and FITC channels were superimposed to facilitate comparison.
Gallyas Silver Staining
Gallyas silver staining (Gallyas 1971) was performed on the same sections as described previously. For purposes of same-section comparisons in this study, the Gallyas silver staining was performed after micrographs and NFT quantitation results had been obtained for the immunohistochemical and thioflavin-S stains.
NFT Density
Typical fields containing high tangle densities were chosen for comparison at three representative regions (CA1, subiculum/presubiculum, and transentorhinal cortex). The numbers of NFTs were counted within the frame of captured images (1340 × 1060 μm2) at ×200 magnification. Histological cues within a section were used to facilitate selection of identical fields so that the counting frame was set at the same positions for the comparison of the three methods. NFT density was calculated as the number of NFTs in the frame divided by the area of the frame. In the transentorhinal region, we followed the pattern of lamination described by Braak and Braak (1992).
Formic Acid Treatment
Formic acid destroys the secondary structure of proteins immediately without cleaving or removing the protein (Cammarata et al. 1990). Sections were treated with 99% formic acid for 2 min and washed thoroughly with PBS. This step was performed either before or after thioflavin-S staining to test the effects of secondary structure on induction or maintenance, respectively, of thioflavin-S-associated staining.
Statistics
NFT densities are expressed as mean ± SE. ANOVA was used to test the significance of difference in NFT densities detected by the three methods.
Results
Improvement of the Thioflavin-S Staining Method
We report a substantial improvement of the thioflavin-S technique that eliminates background cellular autofluorescence, minimizes photobleaching effects, and reduces tissue damage associated with NaOH and acetic acid pretreatment in deparaffinized sections. In the conventional thioflavin-S method, as modified by Vallet et al. (1992), the quenching pretreatment step, consisting of a 20-min KMnO4 incubation followed by NaOH plus H2O2, and finally acetic acid, sometimes causes tissue damage and partial detachment, as demonstrated by local variations in the focal plane (Figure 1A, arrows and arrowheads). In contrast, minimal tissue destruction is encountered in the newly modified procedure (Figure 1B), which involves a shortened 5-min KMnO4 oxidation followed by 1% NaBH4, as described in Materials and Methods. In addition, tangles can be clearly delineated with very little background. A second drawback of the former thioflavin-S staining method is rapid fading of fluorescence intensity, with extensive photobleaching encountered after 1 min of illumination in a fluorescent microscope (Figure 1C). Post-treatment with concentrated phosphate buffer at 4C substantially enhances photostability when tested under otherwise identical conditions (Figure 1D). The post-treatment step also appears to reduce the yellow content of the green signal visible under FITC filters. The use of a DAPI filter set, as recommended by previous protocols, results in a high blue background (Figure 1E) attributable to the intrinsic autofluorescence of tissue proteins. This background is effectively eliminated when viewed under the FITC filter (Figure 1F), with no change in the type or number of pathological entities visible. An optimization of thioflavin-S concentration has determined that 0.05–0.1% thioflavin-S produces an acceptable compromise between signal strength and specificity. The use of 1% thioflavin-S yields strong signal intensity but causes strong nonspecific staining of blood vessels (Figure 1G), whereas 0.01% thioflavin-S stains NFTs with very little nonspecificity but only minimally stains plaques. In the latter case, post-treatment is required to prevent photobleaching.
Comparison of Immunostaining, Thioflavin-S Staining, and Gallyas Silver Staining on the Same Sections
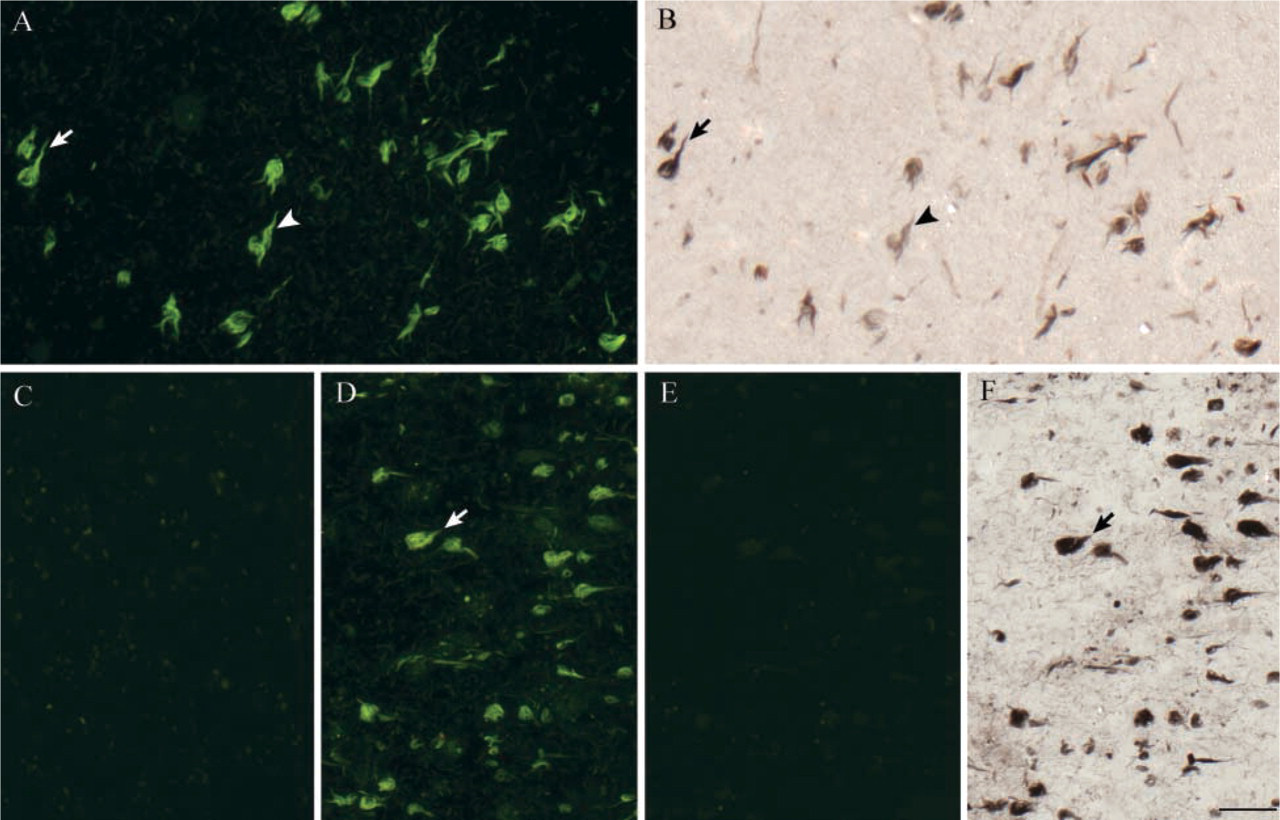
In this study we compared NFT immunostaining, thioflavin-S staining, and Gallyas silver staining in paraffin-embedded sections of various regions of the AD brain. Comparisons were made on the same sections to avoid possible misinterpretations caused by differences among sections and to permit detection of more subtle differences in specificity or sensitivity. Sequential application of anti-NFT immunohistochemistry, our improved thioflavin-S staining, and the Gallyas silver staining revealed co-detection of some tangles in hippocampal CA1 (Figures 2A1–2A4, arrows) and subiculum/presubiculum (Figures 2B1–2B4, arrows). However, the polyclonal anti-NFT antibody rarely detected NFTs in the pri-α layer of transentorhinal cortex (Figures 2C1–2C4), where most NFTs appear early in AD (Braak and Braak 1991), despite the application of antigen retrieval in all cases. On the contrary, this NFT antibody appears to stain plaques only slightly. The lower sensitivity of the anti-NFT immunohistochemical method is demonstrated by a large number of NFTs that were detected by thioflavin-S or Gallyas silver staining but not by NFT immunostaining (Figure 2, arrowheads).
The monoclonal PHF-1 antibody, which recognizes tau protein phosphorylated at serine residues 396 and 404, detected NFTs in both the hippocampus and transentorhinal cortex (Figure 3). However, greater variations in PHF-1 immunoreactivity were observed among AD individuals. Some AD cases showed very few PHF-1-positive NFTs but had a considerable number of NFTs detectable by thioflavin-S. This variation was also reflected in the quantitative comparisons as larger standard errors for PHF-1 staining compared with those for staining with polyclonal NFT antibody (see Table 1). Even in cases where abundant NFTs were stained with PHF-1 antibody (Figure 3), PHF-1 immunostaining still detected fewer NFTs than thioflavin-S.
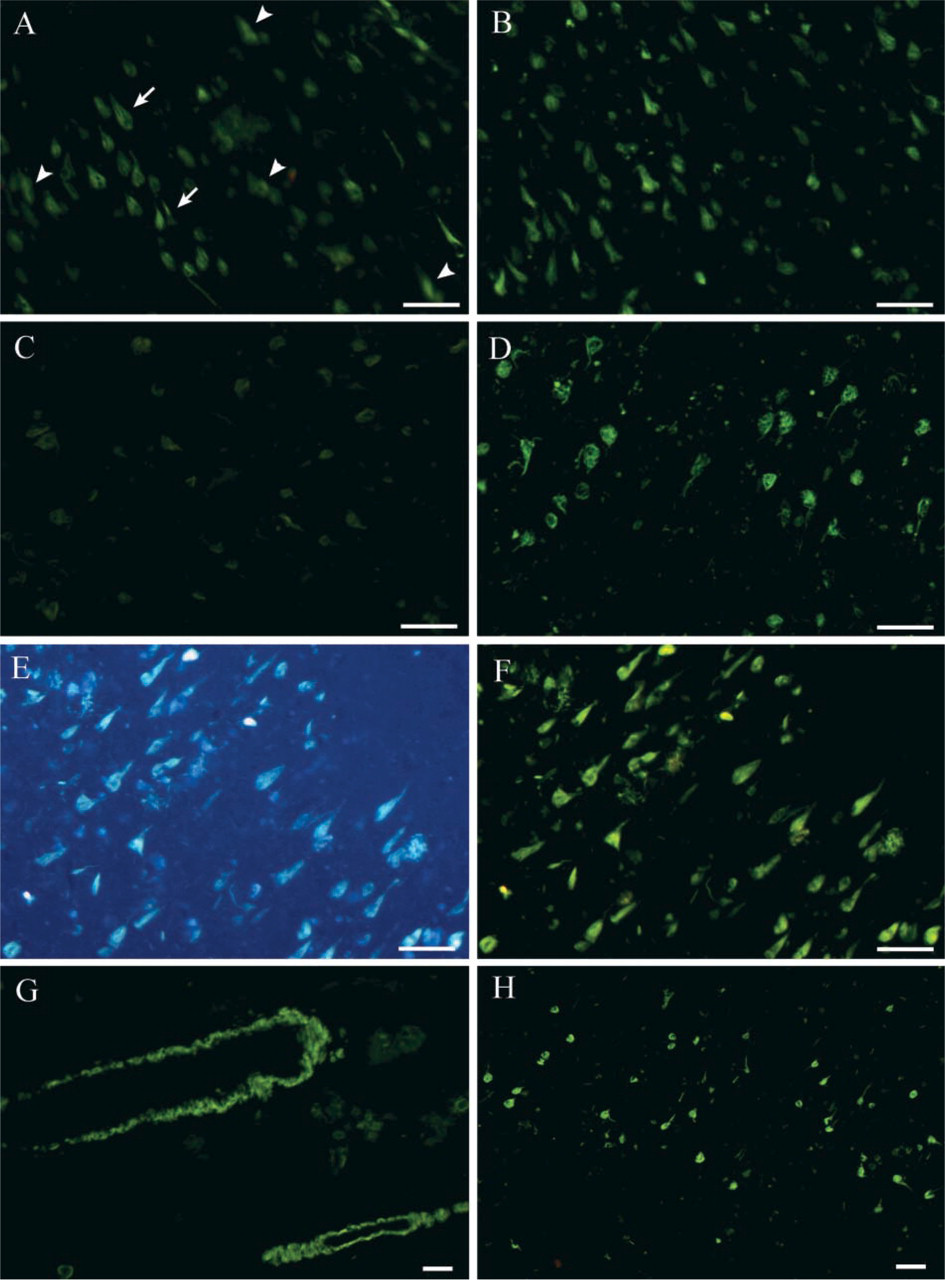
Comparison of the classical thioflavin-S method (left column) and the improved method (right column) on tissue damage (
Staining with Gallyas silver and thioflavin-S on the same fields revealed that these two methods detected the same NFTs (Figure 2C–2D). Some NFTs displayed only very faint staining in 2/14 sections examined (Figure 4B, arrowhead) and in their adjacent sections stained with Gallyas silver alone (data not shown). Gallyas silver staining appeared to detect more neuropil threads (Figure 4A and 4B) but only weakly stained amyloid plaques. In addition, our preliminary results using a recently developed intrinsic fluorescence induction method (Sun et al. 2001) showed that thioflavin-S and Gallyas silver staining methods failed to detect a subpopulation of plaque neurites (data not shown).
Quantification of NFTs showed that the thioflavin-S and Gallyas silver stains identified the same number of NFTs in the three regions examined. In the CA1 and subiculum regions, polyclonal NFT antibody or PHF-1 antibody recognized approximately half of the NFTs detected by the other two methods (Table 1). In the transentorhinal/entorhinal cortex, PHF-1 immunostaining detected more NFTs than the polyclonal NFT antibody (Table 1), although it still left out many NFTs that could be detected by thioflavin-S staining and Gallyas silver staining.
Dependence of Staining on Secondary Structure
Because the thioflavin-S and Gallyas silver staining showed comparable sensitivity in demonstrating NFTs (Table 1), these two methods might share a similarity in staining mechanisms. To address this question, we investigated whether β-pleated sheet secondary structure was necessary for both methods. When applied after treatment with formic acid, which disrupts the structure of β-pleated sheets without cleaving or removing the protein (Kitamoto et al. 1987), the thioflavin-S staining failed to detect NFTs (Figure 4C). Furthermore, addition of formic acid to thioflavin-S-stained sections immediately removed any fluorescence present (Figure 4D and 4E). However, in formic acid-treated sections, the Gallyas silver staining was still able to stain NFTs (Figure 4F). These results indicate that the two methods stain NFTs through different mechanisms.
Discussion
In this study we further improved the thioflavin-S staining method by introducing several technical modifications that result in overall reduction of tissue damage, photobleaching, and background autofluorescence, with no loss of sensitivity or selectivity. Although the modification of the thioflavin-S technique by Vallet et al. (1992) greatly improved the signal-to-background ratio compared to the classical thioflavin-S method, it introduced some problems such as tissue damage, partial detachment, and photobleaching, as found by others (Cullen et al. 1996) and by our preliminary study. Our proposed combination quenching method, using NaBH4 instead of NaOH, increases the efficacy of background quenching and alleviates tissue damage. Treatment with a concentrated phosphate buffer significantly slows photobleaching, possibly by stabilizing critical secondary structural elements. Comparison of this improved thioflavin-S method with other common conventional staining methods on the same sections demonstrated that PHF-1 or NFT immunostaining is much less sensitive in detecting NFTs than the improved thioflavin-S staining and Gallyas silver staining, particularly in the transentorhinal cortex. The similarity in sensitivity between the thioflavin-S and Gallyas silver methods is consistent with previous studies (Vallet et al. 1992; Cullen et al. 1996).
The results of this study show that NFT or PHF-1 immunostaining is much less sensitive than thioflavin-S or Gallyas silver staining in detecting NFTs. The polyclonal anti-NFT antibody used in the present study was created by immunizing against a set of NFTs that were extracted in boiling SDS and purified by sucrose gradient centrifugation (Gorevic et al. 1986). One study had identified at least two morphologically distinct fractions of PHF that can be separated on a sucrose density gradient (Ksiezak-Reding et al. 1994). Morphologically and immunochemically, NFTs are known to be highly heterogeneous (Schmidt et al. 1988; Uboga and Price 2000). Intracellular neurofibrillary tangles (iNFTs) have distinct epitopes from extracellular neurofibrillary tangles (eNFTs, or ghost tangles), which are the insoluble cytoskeletal debris remaining when
Comparison of representative NFT immunostaining, thioflavin-S staining, and Gallyas silver staining on the same sections from AD hippocampal formation. NFT immunofluorescent staining ( Comparison of PHF-1 immunostaining (red) and thioflavin-S staining (green) on the same sections of AD brains at CA1 (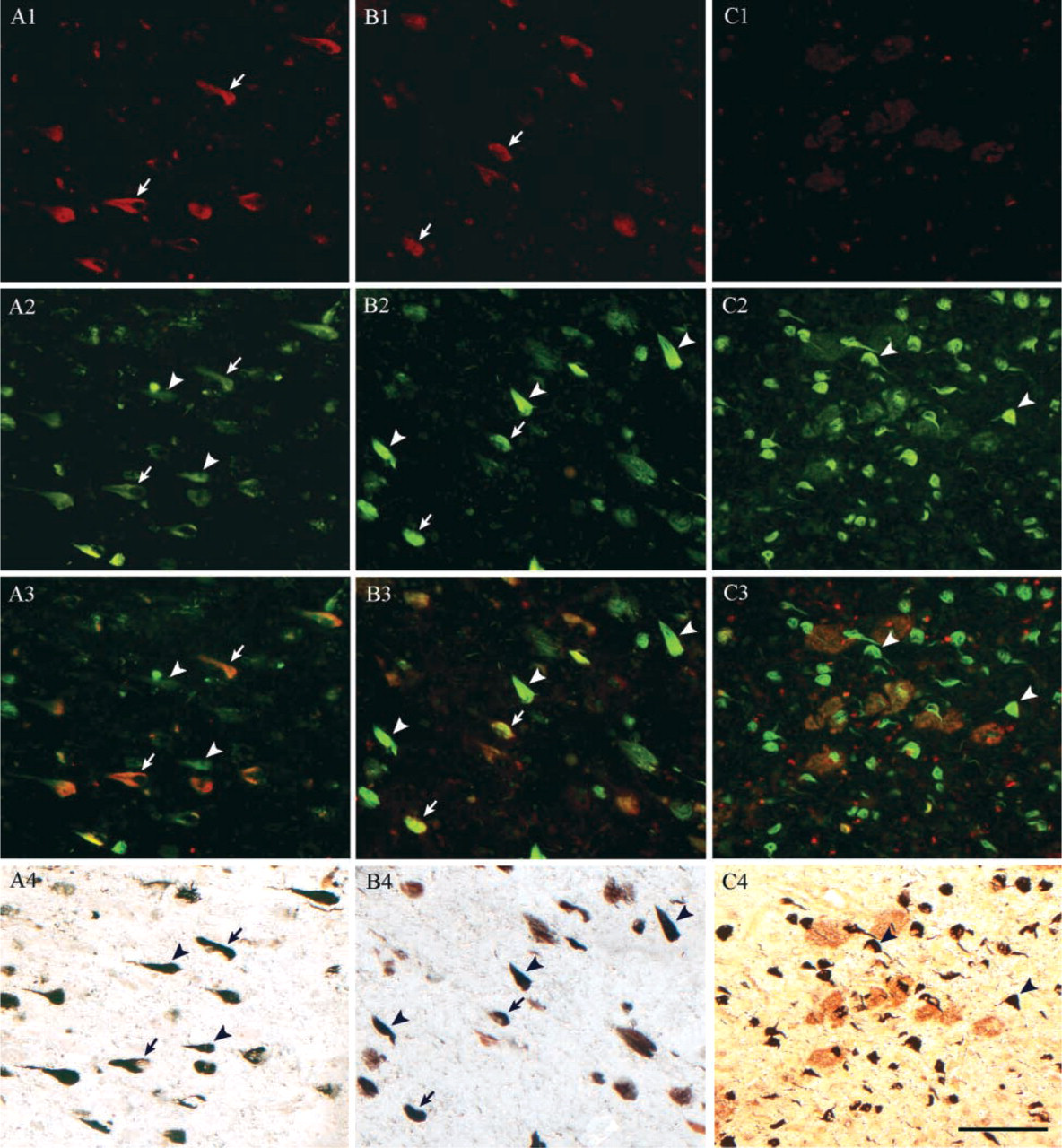
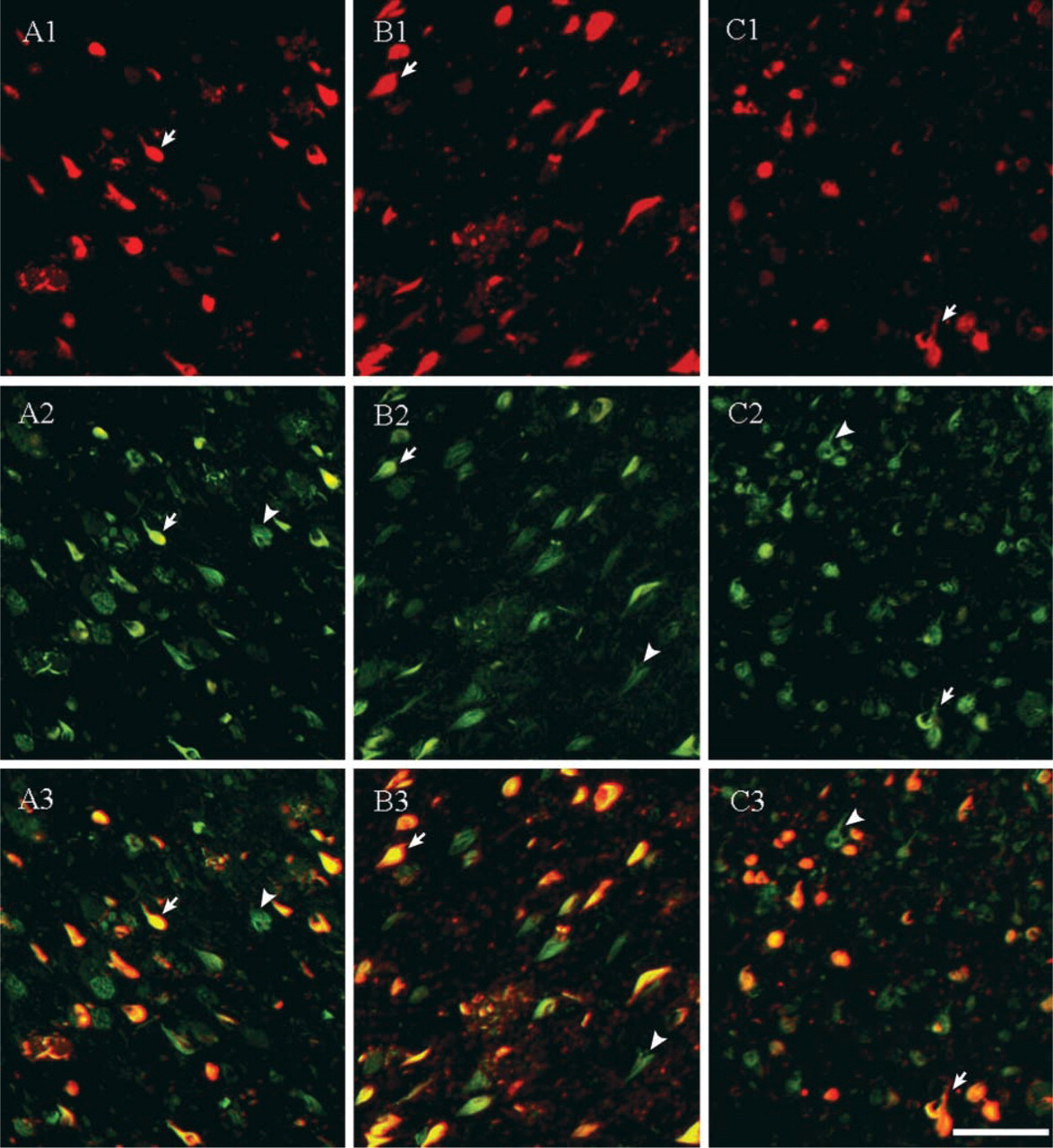
It should also be noted that the PHF-1 antibody used in the present study detects a specific phosphorylated epitope at serine residues 396 and 404, near the C-terminus, of the tau protein. Although the pathways and key phosphorylation events leading to tau hyper-phosphorylation in vivo are poorly understood, in vitro studies show that hyperphosphorylation of tau is part of the mechanism of self-assembly into tangles of PHFs and straight filaments (Alonso et al. 2001). However, iNFTs containing full-length tau do not demonstrate immunoreactivity to some phosphorylation-specific antibodies (Bondareff et al. 1995), which may account for the absence of PHF-1 immunoreactivity in a number of tangles observed in the present study. Despite the relative specificity of this antibody in recognizing late-stage tangles over other forms of pathology, the activity of endogenous phosphatases and proteases may affect sensitivity and could account for part of the individual variability found in this study. The PHF-1 antibody has also been used in earlier studies (Gomez-Isla et al. 1997; Uboga and Price 2000), including a study in which neuronal loss was quantitatively demonstrated to exceed PHF-1 immunoreactivity in AD brains, leading to the conclusion that NFTs may not account for all neuronal loss in AD (Gomez-Isla et al. 1997). However, the results of the present study suggest that caution should be exercised in utilizing PHF-1 immunoreactivity as a quantitative indicator of NFT abundance, particularly in the transentorhinal/entorhinal region, because some NFTs are undetected by PHF-1 immunohistochemistry.
Numerical density of NFTs (mean ± SE) in the AD hippocampus and transentorhinal cortex as detected by immunostaining, an improved thioflavin-S staining, and Gallyas silver staining methods on the same sectionsa,b
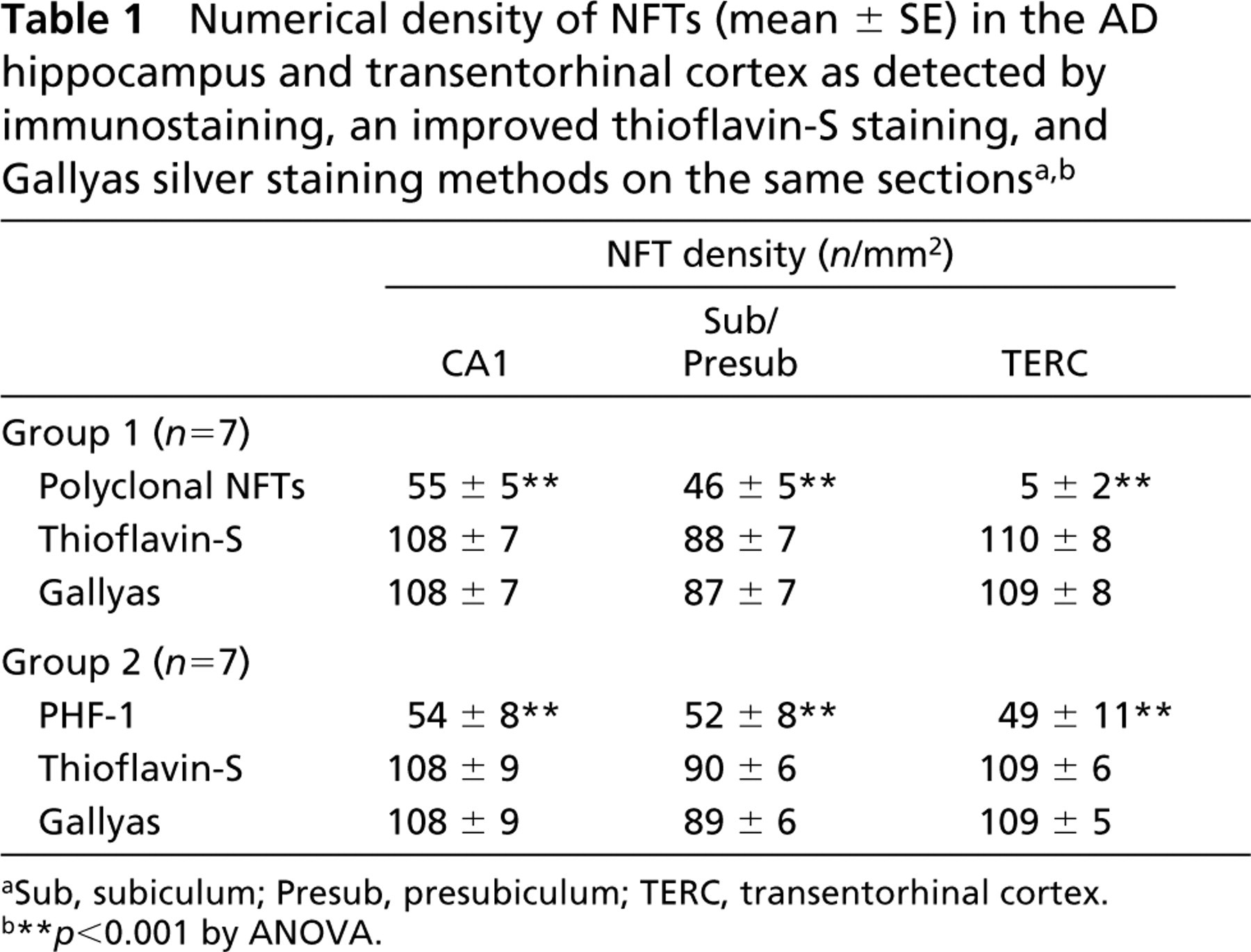
Sub, subiculum; Presub, presubiculum; TERC, transentorhinal cortex.
∗∗ p<0.001 by ANOVA.
Our study showed clear region-specific differences in NFT and PHF-1 immunoreactivity, with transentorhinal and entorhinal cortical areas yielding the fewest immunopositive tangles. Based on the known neuroanatomic distribution of NFTs as a function of AD progression (Braak and Braak 1991), it is likely that these differences reflect changes in available epitopes as the tangle evolves during the course of neuronal degeneration. It has been inferred from neuropathological studies that the transentorhinal/entorhinal regions are the sites of appearance of the first NFTs in AD and that these changes spread in a predictable non-random manner across the hippocampus and neocortex (Braak and Braak 1991; Gertz et al. 1998; Delacourte et al. 1999). The loss of epitopes in the transentorhinal/entorhinal areas may be due to proteolytic processing of eNFTs over time, because eNFTs are only weakly reactive with antibodies to carboxy- or amino-terminal epitopes. Our results are also consistent with the observation that Layer II of the transentorhinal cortex and the subiculum contains a large proportion of eNFTs, especially in severe cases of long duration (Dickson et al. 1992).
Different effects of formic acid treatment on thioflavin-S staining or Gallyas silver staining. Although thioflavin-S staining (
The greater sensitivity of the thioflavin-S and Gallyas silver staining methods compared to anti-tau immunohistochemistry warrants further inquiry into the mechanisms of these stains. The related benzothiazole dye thioflavin-T (ThT), a classical amyloid stain for senile plaques containing β-amyloid peptide in AD brain, binds rapidly and specifically to anti-parallel β-sheet fibrils. The fibrillar β-sheet-bound dye species undergoes a characteristic 120-nm red shift of its excitation spectrum that may be selectively excited at 450 nm, resulting in a fluorescence signal at 482 nm (LeVine H 1997). Thioflavin-S is likely to share a similar mechanism. In our in situ emission scan study, thioflavin-S-stained tissues, when post-treated with concentrated phosphate buffer, displayed a sharp emission peak at 525 nm when excited around 480 nm (data not shown). The emission spectrum of thioflavin-S staining might be wide or might have two peaks, because thioflavin-S staining is visible at both DAPI and FITC channels. Consistent with a binding interaction with fibrillar β-sheet elements, treatment with formic acid immediately destroys thioflavin-S staining. Although formic acid treatment can covalently modify the protein, it usually takes hours (t1/2 = 3.5 hr for β-amyloid) (Klunk et al. 1994). The low background staining indicates that a large array of β-sheet structure is required for binding because thioflavin-S does not appear to stain native proteins that have only a few relatively isolated β-strands. Previously proposed models hypothesize that a minimum of five adjacent β-sheet strands is necessary for Congo red (Klunk et al. 1989), and a similar requirement may apply to thioflavin-S. The intense staining of AD pathology by thioflavin-S raises the question of whether it could identify pathological structures in other neurodegenerative diseases.
The mechanisms of Gallyas silver staining of NFTs have not been fully elucidated. It does not bear an absolute requirement for β-sheet structures because it can stain NFTs in formic acid-pretreated sections. One possibility is that Gallyas silver staining may be related to the loss of the ability of NFTs to reduce silver ions in a manner similar to that of degenerative axons (Gallyas et al. 1980; Gallyas 1982).
In terms of applicability to routine neuropathological diagnosis, the general affinities of thioflavin-S for diverse AD pathologies can be viewed as a technical advantage because it can stain both plaque and tangle pathology with a single and simple technique. Usually, difficulty in long-term preservation of thioflavin-S results limits its wide application in routine neuropathological diagnosis of AD, although the ability to digitally store photomicrographs may obviate this drawback. Our modifications to the procedure, which enhance tissue quality and fluorescence photostability, may make the method more consistent and therefore more amenable to routine clinical use. The Gallyas silver stain is another sensitive method for NFT demonstration but, like other silver stains, is often associated with higher variability than thioflavin-S. One drawback is that, unlike fluorescence-based methods, it lacks the capacity for multiple labeling. Our results with immunostaining suggest that tau immunoreactivity using the two antibodies examined may not be useful for quantitative staining of tangles, although they can be useful in differentiating specific NFT subpopulations. Finally, our general approach to performing multiple stains on same sections may be useful in a number of research situations where individual tangles may need to be identified or categorized based on their affinities for certain antibodies.
Footnotes
Acknowledgements
Supported by NIH grant 3R01 NS39345-01 (to GB).
Human brain sections used in this study were kindly provided by the University of Kentucky Alzheimer's Disease Research Center, headed by Dr William R. Markesbery. Ann Tudor cut the sections for this study.