
Abstract
Adenosine exerts multiple receptor-mediated effects in the heart, including a negative chronotropic effect on the sinoatrial node. The aim of this study was to investigate the distribution of the equilibrative nucleoside transporter rENT1 in rat sinoatrial node and atrial muscle. Immunocytochemistry and/or immunoblotting revealed abundant expression of this protein in plasma membranes of sinoatrial node and in atrial and ventricular cells. Because rENT1-mediated transport is likely to regulate the local concentrations of adenosine in the sinoatrial node and other parts of the heart, it represents a potential pharmacological target that might be exploited to ameliorate ischemic damage during heart surgery.
U
Nucleoside transporters play an important regulatory role in the adenosine-mediated regulation of many physiological processes, including coronary blood flow and myocardial O2 supply-demand balance. In the heart, adenosine regulates pacemaking and contractility through binding to cell surface purinergic receptors. In the pacemaker of the heart (the sinoatrial or SA node), adenosine has a negative chronotropic effect in a number of different species, including humans (West and Belardinelli 1985). Extracellular adenosine and adenosine analogues exert their effects on cells of the SA node by activating adenosine receptors (A1 receptors). Activation of A1 receptors is believed to increase K+ conductance, leading to an inhibition of automaticity in the SA node (West and Belardinelli 1985). Adenosine also directly inhibits the hyperpolarization-activated current, If, in the SA node (Zaza et al. 1994). In addition to these roles in normal physiology, there is now considerable evidence that adenosine exerts beneficial cardioprotective effects in the ischemic/reperfused myocardium (de Jong et al. 2000). The relative rates of adenosine uptake and release from cardiac cells play an important part in such regulatory processes by determining the concentration of this nucleoside available for binding to cell surface receptors. Transporters of the es type are pharmacological targets for the coronary vasodilators dipyridamole and dilazep in many mammalian species (Sundaram et al. 1998). These agents prevent influx of adenosine across the cell membrane, thereby potentiating the interaction of extracellular adenosine with adenosine receptors (Belardinelli et al. 1989).
Previous studies have provided evidence for the existence of an es-type NBMPR-sensitive nucleoside transport process in isolated rat cardiac myocytes (e.g., Geisbuhler et al. 1987). To further understand the physiological roles of adenosine transport in the heart, we have used an immunological approach to investigate the distribution of the es-type transporter rENT1 in the rat SA node and its surrounding atrial muscle.
Materials and Methods
The antibodies used in this study were anti-rENT1226–291 and anti-connexin (Cx43). The anti-rENT1 polyclonal antibody was raised in rabbit against a glutathione-S-transferase fusion protein bearing the central cytoplasmic loop region of rENT1 (residues 226–291; Yao et al. 1997), prepared using the vector pGEX-KT (Hakes and Dixon 1992). This antibody (designated anti-rENT1) was affinity-purified by passage of the antiserum through a column of cellulose binding domain (CBD) fusion protein coupled to Sepharose CL-4B. The anti-Cx43 monoclonal antibody (Chemicon; Harrow, UK) was raised in mouse against rat cardiac Cx43 using a synthetic peptide corresponding to residues 252–270.
Immunocytochemical experiments were carried out on four rats, using tissue sections through the SA node and its surrounding atrial muscle. Rats of either sex weighing 0.2–0.3 kg were sacrified by stunning and cervical dislocation. Dissection and preparation for cryosectioning of the SA node and surrounding atrial muscle were carried out as previously described (Dobrzynski et al. 2000). Sections (20 μm) were mounted on poly-L-lysine-coated glass slides (BDH; Poole, UK) and stored at — 80C until used for immunocytochemistry. Before immunocytochemistry, the tissue sections were fixed with cold methanol (a process that permeabilizes the plasma membrane) and washed three times with 0.01 M PBS (Sigma; Poole, UK). The tissue sections were then blocked with 10% normal donkey serum (NDS) in PBS for 1 hr and were simultaneously double-labeled overnight at 37C with a mixture of anti-rENT1 antibody at 10 μg/ml and anti-Cx43 antibody at 1 μg/ml in PBS containing 6.15 mM NaN3 and 2% bovine serum albumin. On the following day, the slides were washed three times over a 30-min period in PBS and incubated with anti-rabbit and anti-mouse secondary antibodies conjugated to the fluorescent markers FITC and TRITC, respectively (Chemicon) for 1 hr at room temperature. Secondary antibodies were diluted 1:100 in PBS that also contained 1% BSA and 1.5% NDS. After washing three times in PBS over a 30-min period, coverslips were mounted on the slides with 60% glycerol in PBS, sealed with nailpolish, and stored in the dark at 4C for subsequent viewing in a Leica TCS SP laser scanning confocal microscope equipped with argon, krypton, and helium-neon lasers, which allowed excitation at 488 and 568 nm wavelengths for the detection of FITC and TRITC, respectively.
The specificity of the anti-rENT1 antibody was determined by immunoblotting of the intercaval (i.e., SA node-containing) region and atrial and ventricular tissue samples from seven rat hearts. Tissue samples were crushed under liquid N2 and then re-suspended in sample buffer (containing 1% SDS and 300 mM sucrose plus 10 mM EDTA, 0.1 mM iodoacetamide, 0.1 mM benzathonium chloride, and 0.57 mM phenylsulfonyl fluoride as protease inhibitors). After centrifugation at 10,000 × g for 3–4 min, samples (25 μg) of membrane myocardial proteins in the supernantant were separated by electrophoresis on 10% SDS polyacrylamide gels and then transferred to nitrocellulose membranes by semi-dry blotting. After blocking overnight at 4C in 10% dried skimmed milk powder, blots were incubated for 1 hr with 1.25 μg/ml anti-rENT1 antibody in PBS containing 0.05% Tween-20. The bound antibody was detected with a horseradish peroxidase-conjugated anti-rabbit secondary antibody (1:30,000; Dako, Poole, UK). Immunoreactivity was visualized by using a peroxidase-based chemilumines-cent substrate kit (Amersham Life Sciences; Poole, UK).
Results
To assess the distribution of rENT1 in the heart, sections of the rat SA node and its surrounding atrial muscle were simultaneously probed with rabbit anti-rENT1 and mouse Cx43 antibodies. The anti-Cx43 antibody was used as a negative morphological marker for the SA node, as previously described (Dobrzynski et al. 2000). The bound primary antibodies were detected with FITC and TRITC- labeled secondary antibodies, respectively. Figure 1A shows strong plasma membrane-associated immunostaining of rENT1 (green fluorescence) in cells located within the SA node. The absence of immunostaining of Cx43 (red fluorescence) of the majority of the cells that stained strongly for rENT1 is evident in Figure 1B, which shows the same section as in Figure 1A. Figure 1F, in which images shown in Figures 1A and 1B have been merged, confirms the lack of cellular co-localization of rENT1 and Cx43 labeling. Therefore, the majority of the rENT1-positive cells visible in Figure 1A are SA node cells. Similar results were obtained using tissues from four rats.
Immunostaining revealed that rENT1 was also abundant in the atrial muscle of the crista terminalis adjoining the SA node (Figure 1C. As in the case of SA node cells, immunostaining was located predominantly at the plasma membrane, and rENT1 labeling intensity within the crista terminalis (Figure 1C was comparable with that seen in the SA node (Figure 1A. The atrial cells of the crista terminalis stained strongly for Cx43 (Figure 1D, as expected from previous studies (e.g., Dobrzynski et al. 2000). However, the merged image of the double-labeled section shown in Figure 1E indicates that although rENT1 and Cx43 were clearly present in the same cells, labeling was apparently not co-localized at the subcellular level. Strong immunostaining of blood vessel smooth muscle cells was also apparent in the crista terminalis sections (arrows in Figure 1E).
The specificity of the rENT1 staining seen in sections of the SA node and crista terminalis was evident from the lack of staining obtained if the primary antibody was omitted or if it was pre-absorbed with the CBD fusion protein bearing the central cytoplasmic region (residues 226–291) of rENT1 (data not shown). Additional evidence for the specificity of the anti-rENT1 antibody was provided by immunoblotting of membrane preparations from rat heart; tissue was taken from the intercaval region, where the SA node is located, atrium, and ventricle. In each case the antibody stained primarily a single band of apparent molecular mass approximately 60 kD (Figure 2). Similar results were obtained from seven rats. Although the size of rENT1, predicted from its amino acid sequence, is 50 kD, photoaffinity labeling studies have revealed that the es-type transporter from various rat tissues migrates as an N-glycosylated species with apparent molecular mass of 62 kD (Yao et al. 1997). Minor bands of lower and higher molecular masses seen in Figure 2 probably represent degradation products and oligomers of the rENT1 polypeptide, respectively. No bands were evident on the blots if the primary antibody was pre-absorbed with the CBD fusion protein bearing the central cytoplasmic region (residues 226–291) of rENT1 (data not shown).
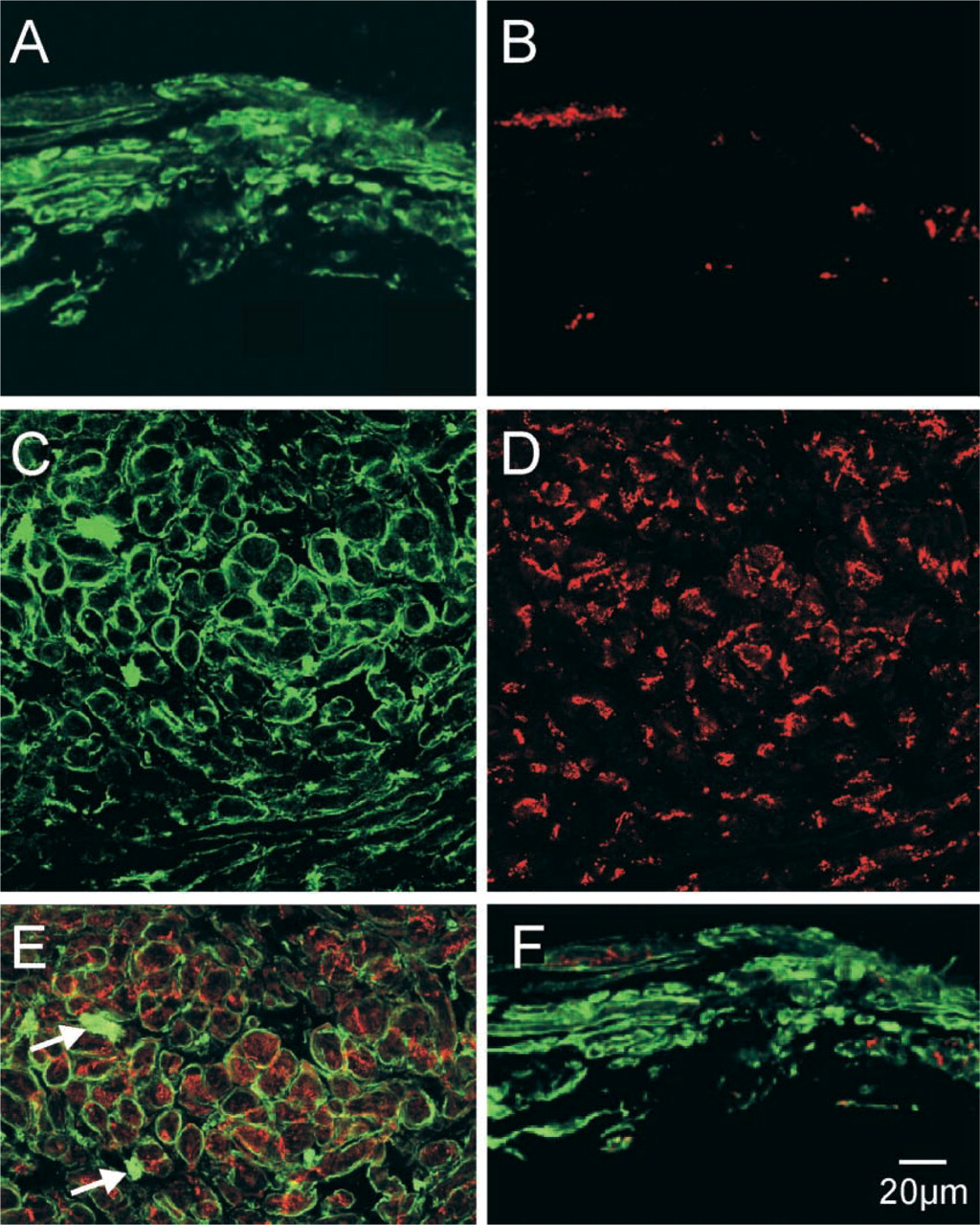
Detection of rENT1 and Cx43 proteins by immunohistochemistry and confocal microscopy in the rat heart. (
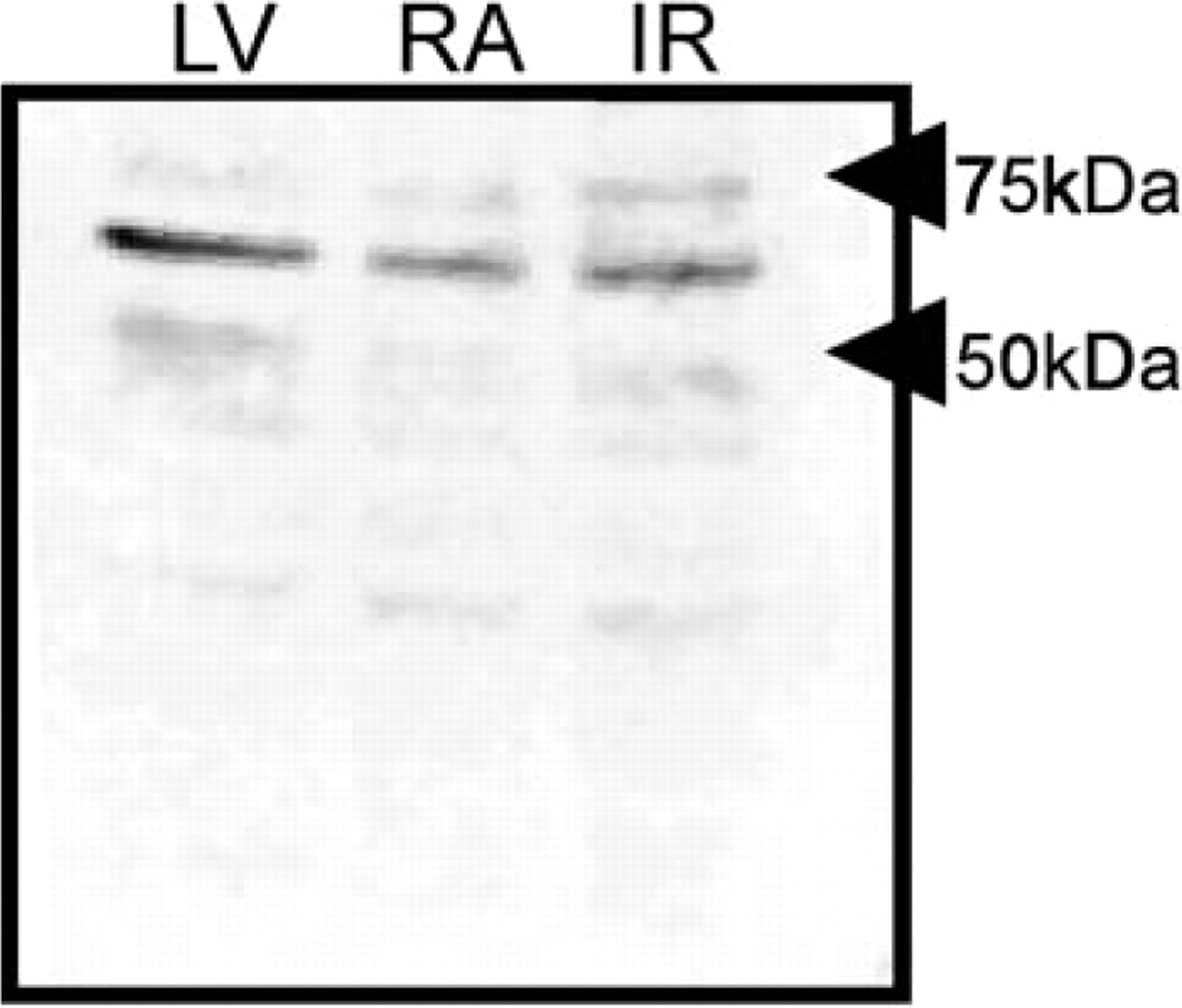
Immunoblots of tissue samples from different regions of rat heart probed with the anti-rENT1 antibody. A major band of approximately 60 kD was detected in all tissues investigated. IR, intercaval region; LV, left ventricle; RA, right atrium.
Discussion
Cardiac Automaticity
Because nucleoside transporters exert effects on the local concentration of adenosine in tissues, the presence of the es-type transporter rENT1 in the rat SA node is likely to be of physiological importance. Pharmacological evidence for this importance has been provided in guinea pig SA and AV nodes, where dipyridamole, a potent inhibitor of es-type transporters in this species, was found to potentiate the chronotropic and dromotropic effects of adenosine, respectively (Meester et al. 1998). The effects of dipyridamole could be accounted for entirely by its inhibition of adenosine uptake (Meester et al. 1998). Although the transporters involved were not identified at the molecular level in the study of Meester et al. (1998), dipyridamole was significantly more potent in enhancing the effects of adenosine on the SA node than on the AV node, suggesting that different transporter types might be involved at these two locations. In this connection, it is interesting that mRNA encoding the ei-type transporter hENT2, which is known to have a lower affinity for dipyridamole than the es-type transporter hENT1 (Ward et al. 2000), has also been detected in human heart (Griffiths et al. 1997). Whatever the identity of the relevant transporter(s) in guinea pig, the dipyridamole effect on SA node cells mimics the sinus bradycardia that occurs when there is a decrease in blood supply to the SA node and a subsequent accumulation of adenosine. Although this phenomenon cannot be mimicked pharmacologically in the rat with dipyridamole, because equilibrative nucleoside transporters of rat are resistant to inhibition by dipyridamole (Yao et al. 1997), similar experiments could be performed using NBMPR, which does potently inhibit rENT1. Moreover, it would interesting to examine the cardiac expression of hENT1, the closely related human homologue of rENT1. If hENT1 is expressed in the human SA node, it might represent a future target for pharmacological agents.
Preconditioning and Ischemia
Adenosine has been suggested to have a cardioprotective role in myocardial ischemia. Myocardial ischemia can be defined as “an imbalance between the amount of oxygen and substrates supplied to the heart and the amount needed to perform normal function” (Verdouw et al. 1998). Murry et al. (1986) first described the phenomenon of “ischemic preconditioning” whereby exposure of the heart to short periods of (non-lethal) ischemia and reperfusion protected the heart from injury during a subsequent prolonged period of ischemia. One of the major factors responsible for the beneficial effects of preconditioning is adenosine, which in the rat heart appears to exert its cardioprotective action by binding both to A1 and A3 purinergic receptors (de Jong et al. 2000). Several recent studies have shown that administration of adenosine to patients before coronary artery bypass surgery or angioplasty reduces myocardial injury (e.g., Lee et al. 1995). Because nucleoside transport can profoundly influence local concentrations of adenosine, pharmacological manipulation of transporters in humans may provide a safe and effective alternative strategy for exploiting the preconditioning abilities of adenosine and thereby attenuating ischemic injury in patients undergoing heart surgery. Transport inhibitors such as draflazine have already been proposed for such strategies (Rongen et al. 1995). Identification of the specific targets of such drugs and their locations in the heart, as described in the present study, should greatly facilitate implementation of this approach.
Footnotes
Acknowledgements
Supported by the British Heart Foundation, the MRC, the Wellcome Trust, the Canadian Institutes of Health Research, and the Alberta Cancer Board. Fatima Abidi was supported by an ORS award. Carol Cass is Canada Research Chair in Oncology.