
Abstract
We isolated cDNA of the mouse homologue of the src-suppressed C kinase substrate (SSeCKS) and analyzed the effects of lipopolysaccharide (LPS) injection on the tissue expression pattern of this protein. Northern blotting analysis showed that SSeCKS mRNA was expressed abundantly in the testis but at undetectable levels in other tissues of untreated control mice. Intraperitoneal administration of LPS strongly induced SSeCKS mRNA expression in the lung, heart, liver, spleen, kidney, lymph node, adrenal gland, and pituitary gland, as well as in the brain. In lung and spleen, the SSeCKS mRNA levels increased almost 10-fold at 1 hr after LPS injection and persisted at high levels until 4 hr. Both in situ hybridization and immunohistochemical studies revealed that LPS administration conspicuously elevated expression of SSeCKS mRNA and protein in vascular endothelial cells of several organs. Ectopic expression of SSeCKS caused loss of cytoplasmic F-actin fibers in the mouse endothelial cell line LEII. These results indicate that SSeCKS is one of the major LPS-responsive proteins and may participate in alteration of cytoskeletal architecture in endothelial cells during inflammation.
Keywords
LIPOPOLYSACCHARIDE (LPS), a component of the outer membrane of Gram-negative bacteria, is a potent inducer of systemic inflammation (Ulevitch and Tobias 1995). Endothelium is known to be a primary site in which LPS-induced inflammatory responses occur. In response to LPS and inflammatory cytokines induced by LPS, the endothelium alters vascular tone and permeability, absorbs infectious insults, and directs leukocytes into the areas of inflammation (Cines et al. 1998). These dynamic changes in endothelium are attributed to alterations in cytoskeletal architecture. Although several molecules, such as protein kinase C (PKC) and tyrosine kinases, are postulated to participate in the cytoskeletal reorganization of endothelial cells (Bannerman and Goldblum 1997; Defazio et al. 2000), the precise molecular mechanisms are not well understood.
Src-suppressed C kinase substrate (SSeCKS) was originally isolated from rat cell lines by Lin et al. (1995) and Chapline et al. (1996). Rat SSeCKS has a motif for binding PKC/protein kinase A (PKA) and is phosphorylated by PKC (Nauert et al. 1996). Several in vitro studies using cell lines have demonstrated that SSeCKS actually scaffolds PKC and PKA and is involved in the regulation of actin-based cytoskeletal dynamics and the cell cycle (Chapline et al. 1998; Gelman et al. 1998; Nelson and Gelman 1997). Previous in vivo studies demonstrated that SSeCKS is expressed in some restricted types of cells, such as spermatids at the late stage of spermatogenesis and renal mesangial cells, suggesting a role for SSeCKS in membrane-cytoskeletal remodeling and cell motility (Erlichman et al. 1999; Nelson et al. 1999; Gelman et al. 2000).
We have searched for inflammation-responsive genes in the mouse brain by differential display analysis and identified 11 of about 1500 genes whose mRNA levels were elevated by IP administration of LPS (Kitamura et al. 2000). In this study we sequenced one of the LPS-inducible cDNA clones, 131.5, and proved it to be a mouse homologue of SSeCKS. We also show that LPS administration dramatically induces SSeCKS mRNA and protein in vascular endothelium of the brain and several visceral organs, and that transfection with SSeCKS cDNA disrupts formation of F-actin fibers in cultured endothelial cells.
Materials and Methods
Experimental Animals and Treatments
Male C57BL/6 mice (7–8 weeks old; SLC, Shizuoka, Japan) and Wistar rats (9–10 weeks old; Hokkaido University, Sapporo, Japan) were housed in plastic cages at 24 ± 1C on a 12-hr light-dark cycle (lights on at 0700 hr–1900 hr) and given free access to laboratory chow and water. Some of them were injected IP with 1–3 mg/kg of LPS (E. coli 055:B5; Difco, Detroit, MI) or sterile PBS. The experimental procedure and care of the animals were in accordance with the guidelines of the Animal Care and Use Committee of Hokkaido University.
Cloning of Mouse SSeCKS cDNA
A mouse testis ΛTriplEX cDNA library (Clontech; Palo Alto, CA) was screened using [α32P]-dCTP-labeled cDNA of a primary isolated partial 131.5 fragment (corresponding to +4797 to +5551bp of mouse SSeCKS cDNA; accession number AB020886) and that of mouse SSeCKS (+ 146 to + 875 bp) obtained by reverse transcription-polymerase chain reaction (RT-PCR) from mouse testis RNA using the following primers: 5′-CCAAGCTCCCACAGAAGAATG-3′ and 5′-GCCCAACCGTGAGTGAAGAA-3′ based on the reported sequences of rat SSeCKS (accession number U23146). Four positive clones were obtained and the remaining nucleotides were amplified by PCR using the ΛTriplEX cDNA library as a template. Sequences of fragments were determined using an ABI PRISM 377 genetic analyzer (Perkin Elmer Applied Biosystems; Foster City, CA).
Northern Blotting
Tissue samples from the mice and rats were taken before and at 0.5–12 hr after LPS or PBS injection. Total RNA was extracted with TRIzol solution (Gibco BRL; Gaithersburg, MD), denatured at 65C, separated on a 1% agarose-formaldehyde gel, and transferred to and fixed on a nylon membrane (Amersham Pharmacia; Piscataway, NJ). A cDNA fragment of rat SSeCKS (+ 2459 to +3159 bp) was prepared by RT-PCR from total RNA extracted from rat testis using the following primers: 5′-ATGAGGACGACCCTAATGTC-3′ and 5′-CTCAACCTTCTCCAGTGCTT-3′. The cDNA probes of mouse SSeCKS (+4797 to +5551 bp) and rat SSeCKS (+ 2459 to +3159 bp) were labeled with [α32P]-dCTP. Nylon membranes were prehybridized with a buffer containing 50% formamide, 5 × SSPE, 0.1% SDS, 5 × Denhardt's solution, and 200 μg/ml salmon sperm DNA at 42C for 10 hr, and then hybridized at 42C for 12 hr in the prehybridization buffer supplemented with 1.0 × 106 cpm/ml of the labeled probes. The blots were washed with 2 × SSC, 0.1% SDS, and 0.1 × SSC, 0.1% SDS solutions, and were exposed to X-ray films for 1 or 2 weeks. The radioactivity was quantified using a BAS-1000 bioimage analyzer (Fuji Film; Tokyo, Japan). The membranes were also hybridized with cDNA of mouse glyceraldehyde-3-phosphate dehydrogenase (GAPDH) or mouse 18s ribosomal RNA (18s rRNA) as a reference.
In Situ Hybridization
Two non-overlapping sense and antisense oligonucleotides corresponding to mouse SSeCKS +4808 to +4852 bp and +4882 to +4926 bp were used as probes for in situ hybridization analysis. These oligonucleotides were labeled with [35S]-dATP using terminal deoxyribonucleotidyl transferase (Promega; Madison, WI). Tissues were freshly removed from mice at 2 hr after LPS or PBS injection and were frozen in liquid nitrogen. Cryostat sections 16 μm thick were prepared and mounted on glass slides pre-coated with 3-amino-propyltriethoxysilane. They were fixed with 4% paraformaldehyde for 10 min and acetylated for 10 min in 0.25% acetic anhydride in PBS. The sections were dehydrated through a graded series of ethanol and prehybridized for 1 hr in a buffer containing 50% formamide, 0.1 M Tris-HCl (pH 7.5), 4 × SSC, 0.02% Ficoll, 0.02% polyvinylpyrrolidone, 0.02% bovine serum albumin (BSA), 0.6 M NaCl, 0.25% SDS, 200 μg/ml yeast transfer RNA, 1 mM EDTA, and 10% dextran sulfate. Hybridization was performed at 42C for 10 hr in the prehybridization buffer supplemented with 1.0 × 107 cpm/ml of the 35S-labeled oligonucleotide probes. The sections were washed at room temperature for 20 min in 2 × SSC containing 0.1% sarkosyl and at 55C for 40 min in 0.1 × SSC containing 0.1% sarkosyl. The sections were either exposed to Hypefilm-βmax (Amersham Pharmacia Biotech) for 3 weeks or dipped in Kodak NTB2 nuclear track emulsion and exposed for 2 months. All sections were counterstained with hematoxylin.
Immunohistochemistry
Immunostaining of SSeCKS was carried out according to the avidin-biotin complex method with rabbit anti-rat SSeCKS serum (a kind gift from Dr. Irwin H. Gelman; Mount Sinai Medical Center, New York, NY). The crossreactivity of the serum against mouse SSeCKS was confirmed by Nelson et al. (1999) and Gelman et al. (2000). Western blotting analysis revealed two major bands (280 and 290 kD) as well as two minor bands (240 and 43 kD) in an extract from mouse testis (unpublished observations). At 0, 2, 4, 8, 12, or 16 hr after LPS or PBS injection, mice were anesthetized by a lethal dose of pentobarbital and perfused through the aorta with PBS, followed by 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4). Tissues were removed and immersed in the same fixative for an additional 6 hr. After dipping in 30% sucrose solution, the specimens were frozen in liquid nitrogen. Frozen sections 16 μm thick were prepared in a cryostat and incubated in 0.3% Triton X-100 for 30 min, followed by 0.3% H2O2 in absolute methanol for 10 min. After pretreatment with 5% normal goat serum for 30 min, the sections were incubated with the primary antibody (1:4000, rabbit anti-rat SSeCKS serum) overnight. The sections were then treated with a biotinylated goat anti-rabbit antibody (Histofine; Nichirei, Tokyo, Japan) followed by application of avidin-horseradish peroxidase complex (Vector Laboratories; Burlingame, CA), each for 1 hr. The antigen-antibody reaction was visualized by incubation in 0.05 M Tris-HCl buffer (pH 7.6) containing 0.01% 3,3′-diaminobenzidine and 0.002% H2O2. All sections were counterstained with hematoxylin.
For immunoelectron microscopy, the liver, brain, spleen, and lung were processed for a pre-embedding silver-intensified immunogold method. Paraformaldehyde-fixed cryostat sections were incubated in the SSeCKS antiserum diluted 1:2000, and subsequently reacted with goat anti-rabbit IgG covalently linked to 1.4-nm gold particles (Nanogold; Nanoprobes, Stony Brook, NY). After silver enhancement (HQ silver; Nanoprobes), sections were osmificated, dehydrated, and embedded in Epon 812 (Nisshin EM; Tokyo, Japan). Ultrathin sections were prepared and stained with an aqueous solution of 2% uranyl acetate for observation under an electron microscope (JEM-100SX; JEOL, Tokyo, Japan).
Ectopic Expression of SSeCKS mRNA in Cultured Endothelial Cells
The SSeCKS mRNA and its antisense expression plasmids for transfection were constructed by subcloning the full-length mouse SSeCKS cDNA (+1 to +5055 bp) into pDEST12.2 (Gibco) using the Gateway system (Gibco) according to the manufacturer's directions. Mouse endothelial cell line LEII was kindly provided by Dr. A. S. G. Curtis (University of Glasgow, UK) and was cultured in Dulbecco's modified Eagle's medium (Sigma; St Louis, MO) containing 10% fetal calf serum. LEII cells were transfected with a construct for sense or antisense SSeCKS, or with control pDEST12.2 using FuGENE reagent (Roche; Branchburg, NJ) for 3 days. Some cells were subsequently incubated with G418 neomycin (500 μg/ml) for 2 weeks. After washing with PBS, cells were seeded onto glass coverslips and cultured overnight. The cells were fixed with 4% paraformaldehyde in PBS for 10 min and permeabilized in 0.25% Triton X-100 for 5 min. They were then incubated with the anti-SSeCKS serum (1:250) in 1% BSA-containing PBS for 2 hr, blocked with 3% BSA in PBS for 1 hr, and finally incubated with both fluorescein isothiocyanate-labeled goat anti-rabbit IgG (1:500; Wako, Osaka, Japan) and phalloidin-rhodamine (1:80; Molecular Probes, Eugene, OR) for 1 hr in the dark. The coverslips were washed with PBS and observed with a confocal scanning fluorescence microscope (FLUOVIEW system; Olympus, Tokyo, Japan).
Statistics
All values were expressed as means ± SEM. Statistical comparison was done by analysis of variance, followed by Scheffe's F-test.
Results
cDNA Cloning of Mouse SSeCKS
We screened inflammation-responsive genes in the mouse brain by differential display analysis and found one cDNA clone, 131.5, that markedly increased after IP administration of LPS. The full-length 131.5 cDNA had an open reading frame of 5055 bp encoding a polypeptide of 1684 amino acids, which was 84.8% identical to rat SSeCKS (GenBank accession number U23146). The deduced polypeptides of 131.5 preserved four PKC phosphorylation motifs, four serine residues phosphorylated by PKC, one PKA-binding site, and one N-terminal myristylation signal as reported in rat SSeCKS (Lin et al. 1996; Chapline et al. 1998; Erlichman et al. 1999), and therefore was concluded to be mouse SSeCKS (Figure 1). Mouse SSeCKS was 88 amino acids longer than rat SSeCKS at the C-terminal region because of the insertion of a cytosine ribonucleotide in its mRNA at +4639 bp, resulting in a frame shift. Moreover, a putative zinc-finger domain near the C-terminus reported in rat SSeCKS was not identified in mouse SSeCKS. These data for mouse SSeCKS were registered in the database of GenBank/EMBL/DDBJ (accession number AB020886).
SSeCKS mRNA Expression After LPS Administration
The mRNA expression of SSeCKS in mouse tissues was investigated by Northern blotting analysis (Figure 2). In control mice injected with PBS, SSeCKS mRNA was at undetectable or quite low levels in all tissues examined except testis. Testis showed abundant SSeCKS mRNA expression, which was not influenced by LPS injection. However, remarkable induction of SSeCKS mRNA was found in brain, lung, heart, liver, spleen, kidney, lymph node, adrenal gland, and pituitary gland at 2 hr after IP injection of LPS. Similar induction of SSeCKS mRNA by LPS injection was also observed in rats (Figure 2). Figure 3 shows the time-dependent expression of SSeCKS mRNA after LPS injection in mouse lung and spleen. The SSeCKS mRNA level rose 10-fold in lung and spleen at 1–4 hr and 1–2 hr after LPS injection, respectively, and declined to lower levels at 12 hr.
In Situ Hybridization of SSeCKS mRNA
Cellular localization of SSeCKS mRNA was examined in PBS- and LPS-injected mice by in situ hybridization using 35S-labeled antisense oligonucleotides as probes. Selective expression of SSeCKS mRNA was found only in testis in PBS-injected mice, whereas at 2 hr after LPS injection intense signals of SSeCKS mRNA appeared in many tissues, including pituitary gland, lymph node, adrenal gland, lung, kidney, heart, liver, lung, brain, and spleen (Figure 4). Two non-overlapping antisense oligonucleotide probes gave essentially the same labeling patterns, whereas no specific hybridization signals were obtained with sense oligonucleotide probes (Figure 4).
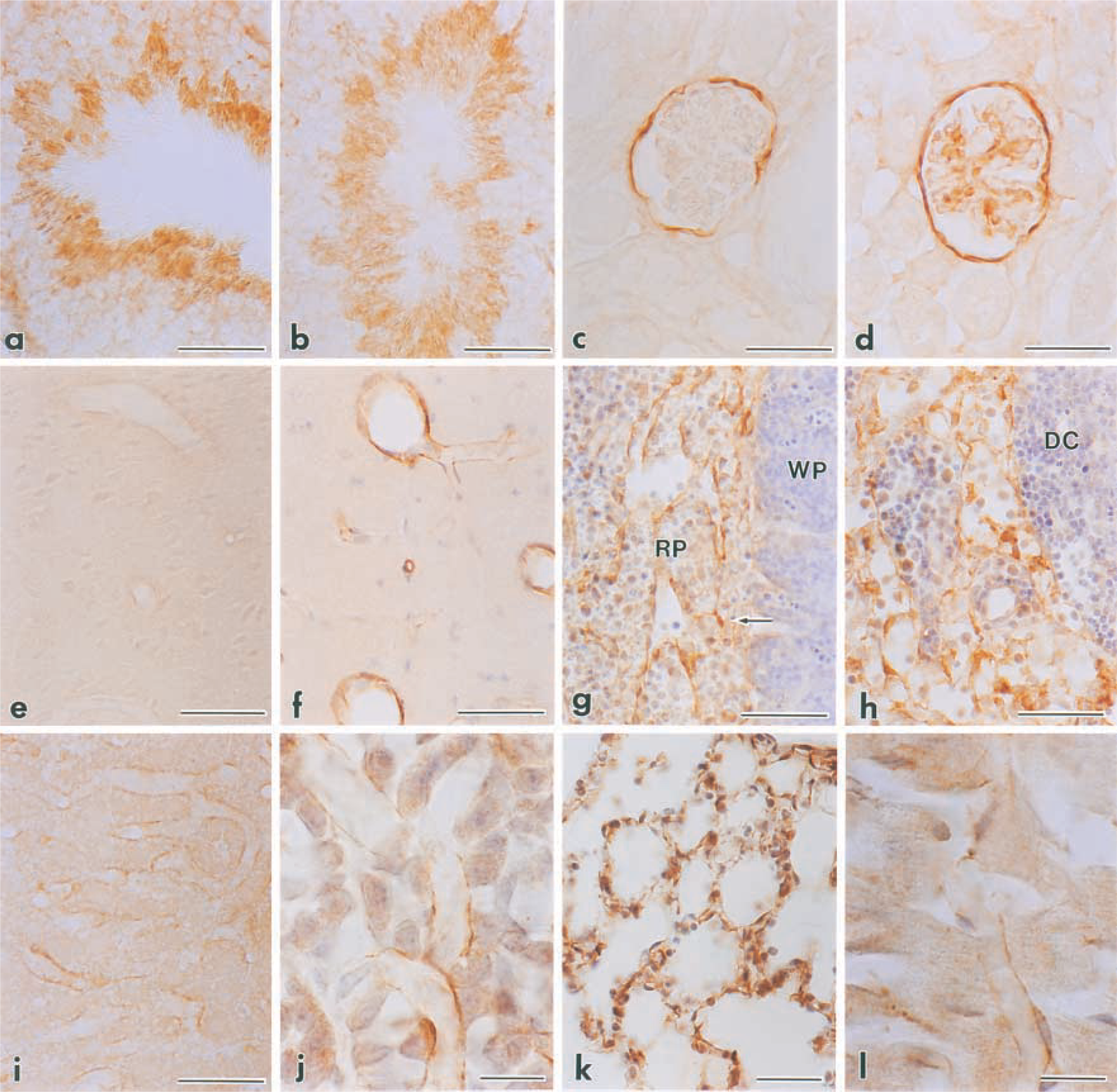
By light microscopy, SSeCKS mRNA in control mice was abundantly present at the luminal side of the seminiferous tubules in the testis (Figure 5a), although the expression level differed considerably among the individual tubules depending on the stage of spermatogenesis. LPS treatment did not affect the SSeCKS mRNA expression in the testis (Figure 5b). In the kidney in control mice, weak but significant signals of SSeCKS mRNA existed at the periphery of renal corpuscles, which corresponded to Bowman's capsules (Figure 5c). After LPS stimulation, signals in the kidney were not restricted to Bowman's capsules but appeared diffusely and intensely in the renal glomeruli (Figure 5d). Signals in other tissues did not exceed the background levels in the control, whereas they prominently increased at 2 hr after LPS injection (Figures 5e and 5f). In the brain, the SSeCKS mRNA signal exhibited many linear structures in the parenchyma and along the brain surface, corresponding to vascular endothelial cells and pia mater, respectively (Figure 5f). The signals in the spleen and lymph node were recognized selectively in the red pulp and medullary sinuses, respectively (Figures 5g and 5h). In the liver, intense signals of SSeCKS mRNA were localized in slender cells along the sinusoidal walls but not in hepatocytes (Figure 5i). The anterior pituitary showed a similar image: dispersed cells among endocrine cells were intensely labeled (Figure 5j). Diffuse signals were distributed throughout the lung, except for non-labeled bronchioles (Figure 5k), and in the adrenal cortex (Figure 5l) and heart (data not shown).

Amino acid sequences of mouse and rat SSeCKS. Identical residues in the two species are boxed. The arrowhead indicates the site of a frame shift. Asterisks represent residues phosphorylated by PKC. Four PKC phosphorylation sites, one PKA-binding site, and one N-terminal myristylation signal are also indicated.
Immunohistochemical Localization of SSeCKS Protein
For detailed identification of SSeCKS-expressing cells, immunohistochemical examinations were conducted using a rabbit antiserum against rat SSeCKS protein. In the testis in control mice, the SSeCKS immunoreactivity existed only in elongated spermatids at the late stage of spermatogenesis, and this staining pattern was not changed by LPS injection (Figures 6a and 6b). The immunoreactivity in the kidney was restricted to the epithelial cells lining Bowman's capsules in control mice, whereas it also appeared intensely in mesangial cells in LPS-injected mice (Figures 6c and 6d). In other organs, the SSeCKS immunoreactivity was undetectable in control mice, in which it became noticeable after LPS injection, being strongest at 8–12 hr. The LPS-induced immunoreactivity was localized in endothelial cells and reticular cells. For example, at 8 hr after LPS injection intense immunoreactivity was observed in all types of vessels (arterioles, capillaries, and venules) in the brain (Figures 6e and 6f) and in those of sinuses of the spleen (Figure 6g). LPS also induced SSeCKS immunoreactivity in reticular cells in the red pulp of the spleen and the medulla of the lymph node (Figures 6g and 6h). The immunoreactive reticular cells in the lymph node were distributed in the lumina of the medullary sinuses and lined the sinus walls as lymphatic endothelium (Figure 6h). The endothelial cells of capillaries and sinusoidal capillaries were predominantly immunoreactive in the liver (Figure 6i), adenohypophysis (Figure 6j), lung (Figure 6k), heart (Figure 6l), and adrenal cortex (data not shown). Thus, the cellular localization of SSeCKS protein was almost identical to that of its mRNA revealed by in situ hybridization.
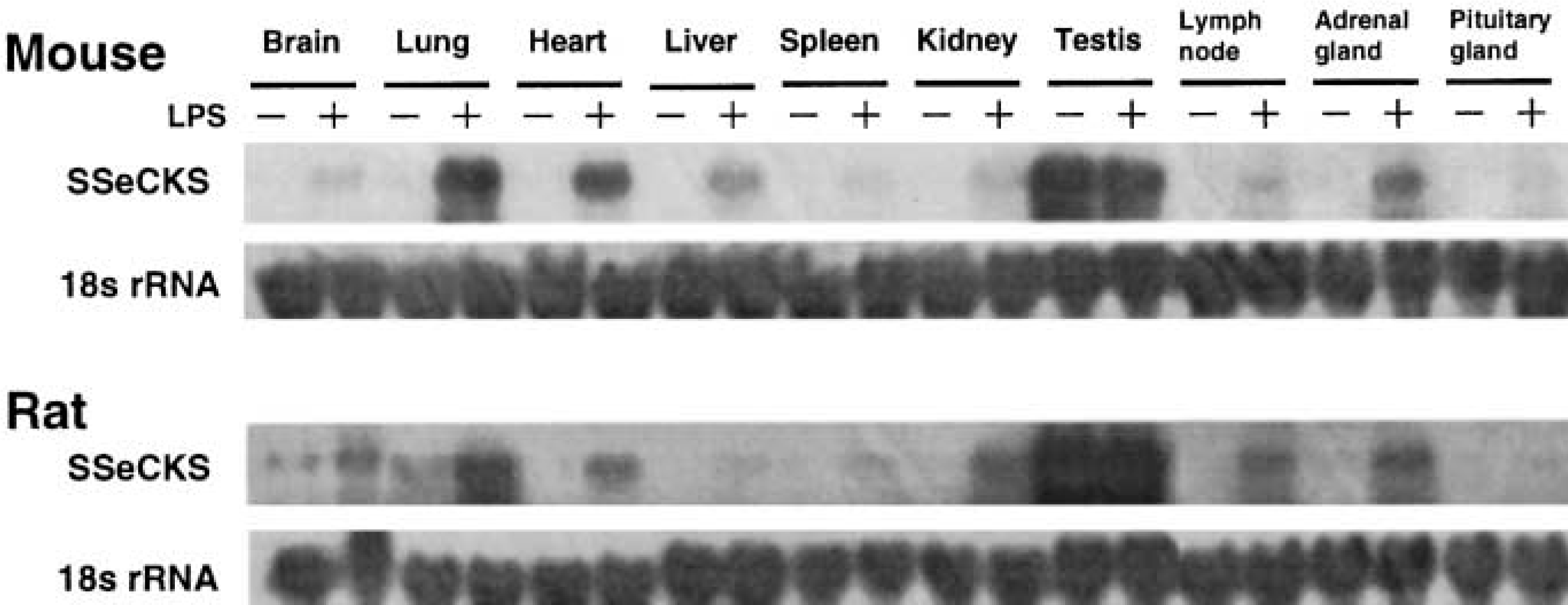
LPS-induced SSeCKS mRNA expression in the various tissues of the mouse and rat. Mice and rats were injected IP with LPS (mice 3 mg/kg; rats 1 mg/kg) or PBS. At 2 hr (mouse) and 3 hr (rat) after the injection, 40 μg of total RNA was isolated and subjected to Northern blotting analysis using SSeCKS and 18s rRNA probes. Results are representative of three independent experiments. For the lymph node, pituitary gland, and adrenal gland, tissues from two to 10 animals were pooled for RNA extraction.
To confirm the localization of SSeCKS in the endothelial cells, we performed immunoelectron microscopy using the silver-intensified immunogold method. As shown in Figure 7, SSeCKS-gold particles were detected mainly in the cytoplasmic region of sinusoidal endothelial cells in the liver. SSeCKS immunoreactivity was also confined to endothelial cells and was undetectable in pericytes, vascular smooth muscle cells, and parenchymal cells in the brain, spleen, and lung (data not shown).
F-actin Fibers in Endothelial Cells After Transfection with SSeCKS cDNA
It has been argued that SSeCKS regulates the cytoskeletal architecture in cultured fibroblasts and mesangial cells (Gelman et al. 1998; Nelson et al. 1999). To clarify the role of SSeCKS in endothelial cells, we examined the relationship between SSeCKS and F-actin fibers in the cultured endothelial cell line LEII. First we examined mRNA and protein levels of SSeCKS in LEII cells before and after LPS stimulation. LEII cells expressed considerable levels of SSeCKS mRNA and protein, which were not increased even after LPS stimulation (data not shown). Then we changed the intracellular SSeCKS levels of the cells by transfection of SSeCKS cDNA. Cells transiently transfected with SSeCKS cDNA were intensely stained with the anti-SSeCKS antibody. The immunoreactivity was localized predominantly in the perinuclear region and near the plasma membrane (Figure 8A). The cells ectopically expressing SSeCKS showed decreased F-actin fibers compared with non-transfected cells (Figure 8A). We also observed F-actin fibers in cells stably transfected with a sense or antisense construct for SSeCKS or a control plasmid (Figure 8B). Compared with control cells, cells expressing SSeCKS clearly caused F-actin fibers to diminish in the central area of the cell. In contrast, cells transfected with the antisense construct had more abundant F-actin structures in cytoplasm. Therefore, increased expression of SSeCKS disrupted organization of F-actin fibers in cultured endothelial cells.
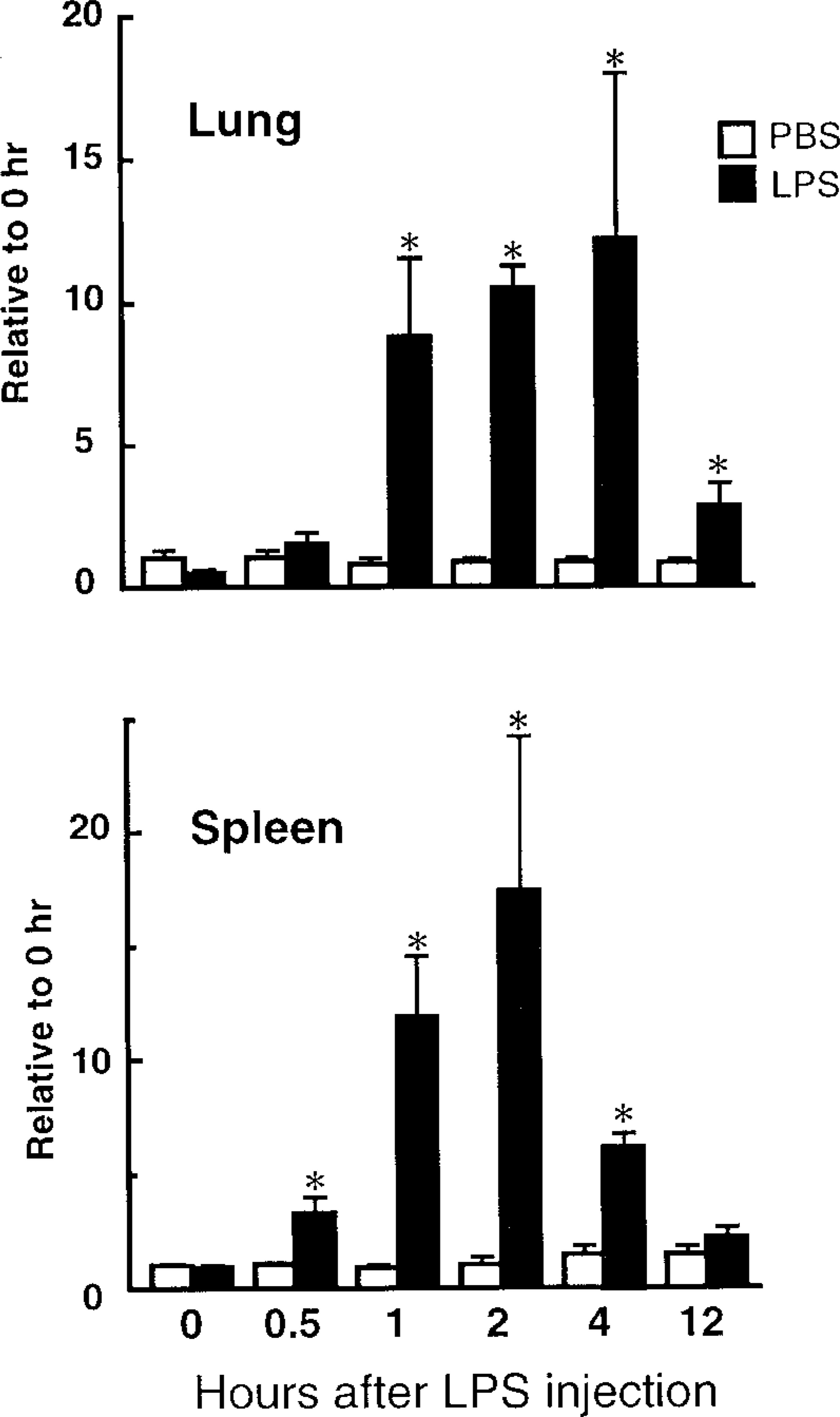
Time course of the LPS-induced SSeCKS mRNA expression in the mouse lung and spleen. Forty μg of total RNA was isolated at 0–12 hr after IP injection of LPS (3 mg/kg) or PBS. For quantitative analyses, the SSeCKS mRNA level was normalized by those of GAPDH mRNA, and expressed as relative to those of the 0-hr PBS control. Neither LPS nor PBS injection caused any significant alteration in the GAPDH mRNA level. Values are means ± SEM for three mice. ∗ p < 0.05 compared with the PBS-injected group.
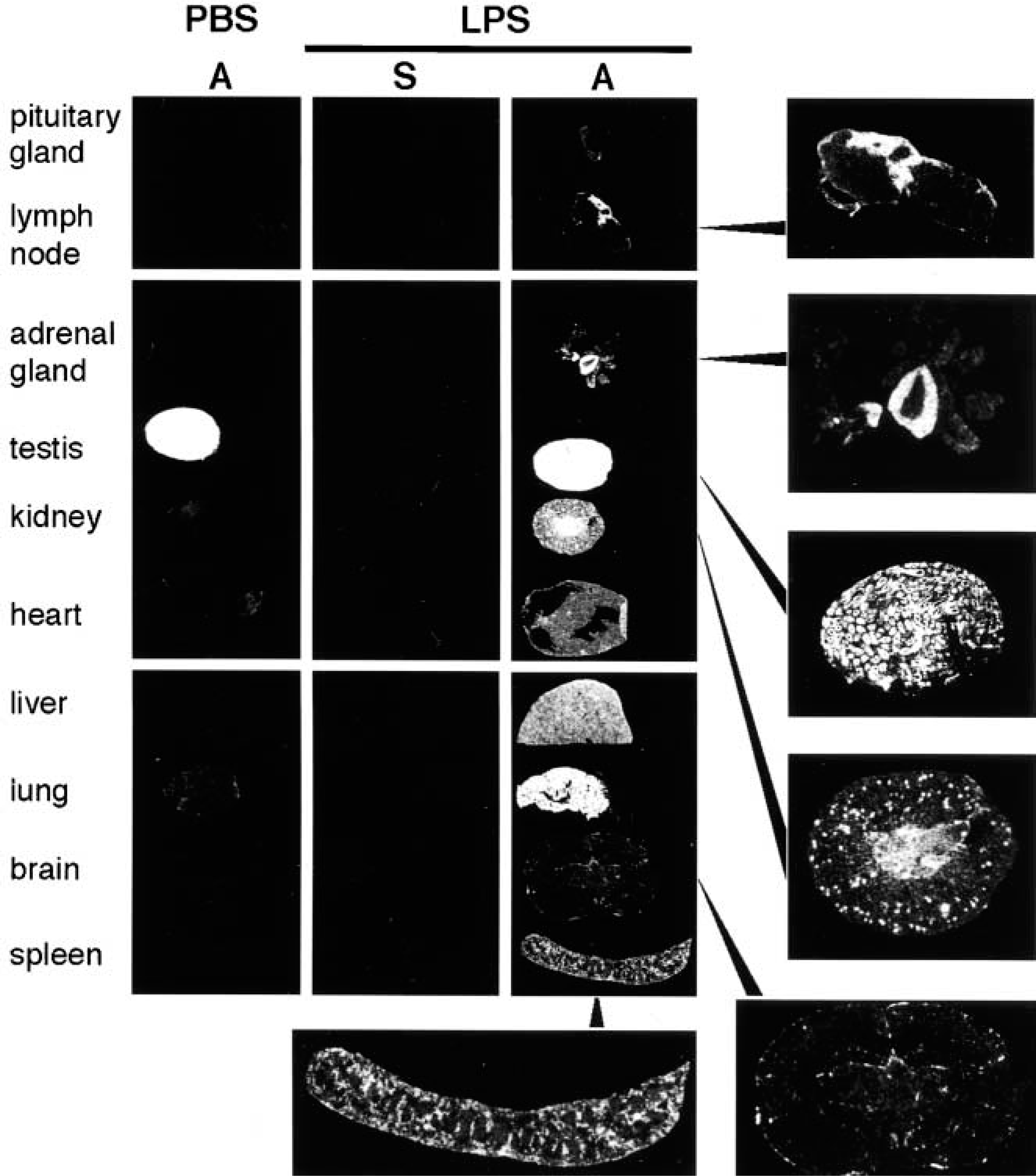
Macroscopic images of SSeCKS mRNA signals in the mouse tissues. At 2 hr after IP injection of LPS (3 mg/kg) or PBS, tissues were taken and frozen sections were processed for in situ hybridization. Sections were hybridized with either an antisense (A) or sense (S) oligonucleotide probe corresponding to +4808 to +4852 bp of mouse SSeCKS cDNA. Essentially the same results were obtained using another oligonucleotide probe set of mouse SSeCKS, +4882 to 4926 bp. Results are representative of three independent experiments.
Discussion
Using a differential display technique, we cloned mouse SSeCKS as one of the molecules whose expression was conspicuously induced in the brain and several visceral organs by IP administration of LPS. Similar to rat SSeCKS (Nauert et al. 1996), mouse SSeCKS possessed four PKC phosphorylation sites, a PKA-binding site, and an N-terminal myristylation site. In contrast, mouse SSeCKS lacked a putative zinc-finger domain found in rat SSeCKS. Moreover, it exhibited an 88-amino acid extension in the C-terminus because of one nucleotide insertion in its mRNA. These differences between the rodent species suggest that the zinc-finger domain and C-terminus are not necessary for the function of SSeCKS.
The most notable finding of the present study was that IP LPS injection induced expression of SSeCKS predominantly in vascular endothelial cells of several organs. The LPS-induced SSeCKS expression was demonstrated both at the mRNA level by Northern blotting and in situ hybridization analyses and at the protein level by immunohistochemical examination, i.e., LPS injection caused rapid (1–4 hr) and remarkable (~10-fold) increases in mRNA expression, followed by the obvious appearance of immunoreactivity for SSeCKS in 8–12 hr. It should be emphasized that the SSeCKS expression in these cells was undetectable in the normal condition without LPS injection. Previously, Gelman and colleagues (Nelson et al. 1999; Gelman et al. 2000) reported the relatively ubiquitous localization of SSeCKS immunoreactivity in a variety of cells and tissues of normal adult mice, including the neural tissues, smooth/cardiac muscles, intestinal/urinary/bronchiolar epithelia, spermatids/sperm, Sertoli cells, epithelial cells of Bowman's capsules, mesangial cells, fibroblasts, and macrophages. It was also reported that vascular endothelial cells in the kidney, heart, and lung were immunoreactive with intermediate levels of intensity (Gelman et al. 2000). In the present study we used the same antiserum as theirs but failed to detect any significant immunoreactivity in these cells of control mice, except for spermatids and Bowman's capsules. The discrepant results may have arisen from the different staining procedures employed. We stained cryostat sections using a highly diluted antiserum (1:4000), whereas they stained paraffin sections using the antiserum at low dilutions (1:250–500). In general, the antigenicity in immunostaining is better preserved in cryostat sections than in paraffin sections, and nonspecific binding is minimized by use of a highly diluted antiserum. Nevertheless, LPS specifically induced SSeCKS protein in endothelial cells and reticular cells, which was supported by the present Northern blotting and in situ hybridization analyses of SSeCKS mRNA.
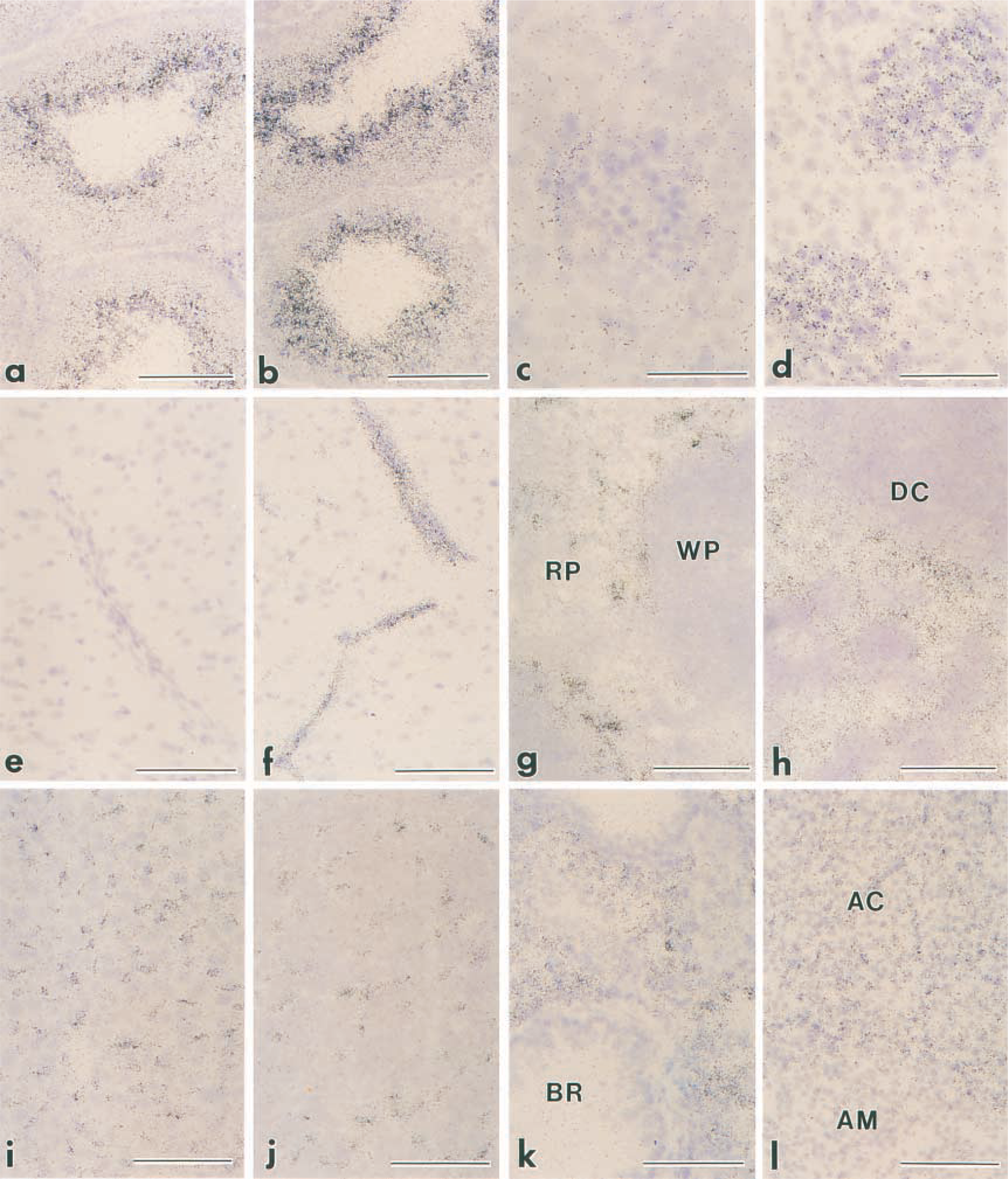
In situ hybridization of SSeCKS mRNA in the mouse tissues after LPS injection. Mice were injected with either PBS as a control (
Immunohistochemical localization of SSeCKS in the mouse tissues after LPS injection. Mice were injected with either PBS as a control (
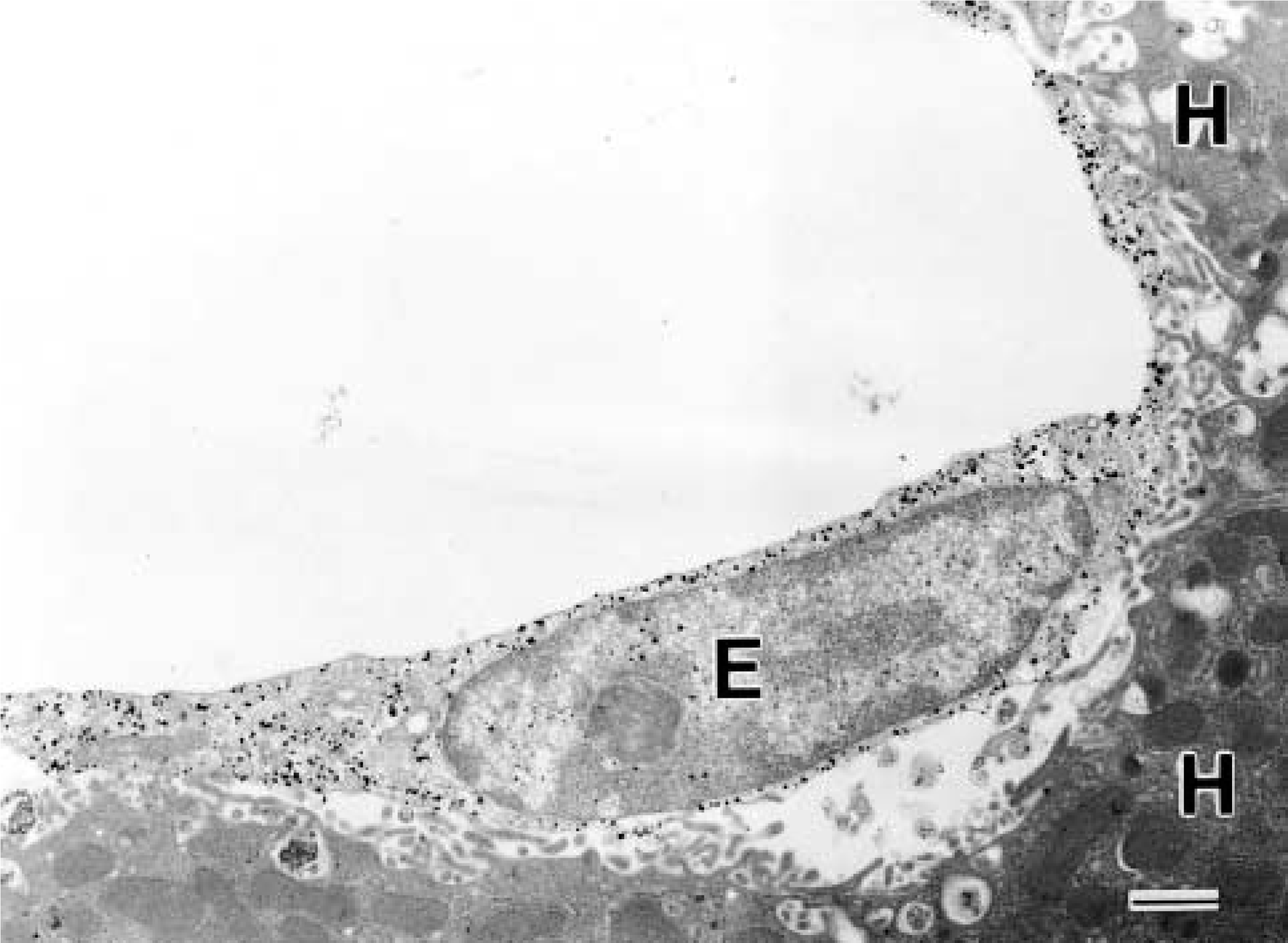
Immunoelectron microscopy for SSeCKS in the liver after LPS injection. Gold particles densely aggregate on an endothelial cell (E) but not on hepatocytes (H). Bar = 1 μm.
Cells expressing SSeCKS in response to LPS included reticular cells of the spleen and lymph node as well as endothelial cells of the adenohypophysis, lung, spleen, liver, and adrenal cortex. These cells are members of the reticuloendothelial system (RES) that take up exogenous substances such as lithium carmine, glycosaminoglycans, proteoglycans, collagen, lipoprotein, and denatured erythrocytes (Praaning van Dalen et al. 1987; Kawai et al. 1998; Smedsrod et al. 1990; Terada 1993). LPS is known to induce myristoylated alaninerich C kinase substrate (MARCKS), a PKC substrate sharing biochemical and structural characteristics with SSeCKS (Thelen et al. 1990; Seykora et al. 1991), which participates in phagocytosis by the actin-dependent translocation of phagocytic vesicles in macrophages (Allen and Aderem 1995). Previous in vitro studies have suggested that SSeCKS controls the actin-based cytoskeletal architecture during cell migration, cytokinesis, and tissue development under the regulation of PKC. For example, transient stimulation of cells by phorbol esters diminishes F-actin fibers coincident with the phosphorylation and translocation of SSeCKS to perinuclear sites (Lin et al. 1996). Ectopic expression of SSeCKS in cultured fibroblasts also induces a temporal loss of F-actin fibers and vinculin-associated adhesion plaques, and the formation of filopodia- and lamelliopodia-like extensions (Gel-man et al. 1998). The present study also confirmed a significant loss of F-actin fibers caused by ectopic expression of SSeCKS in the mouse cultured endothelial cell line LEII. These results, together with findings that an F-actin disassembler, cytochalasin D, stimulates endocytosis (Song et al. 1999), appear to indicate that SSeCKS regulates F-actin formation of endothelial and reticular cells in vivo and contributes substantially to uptake of exogenous particles and macromolecules from blood and lymph circulation during inflammation.
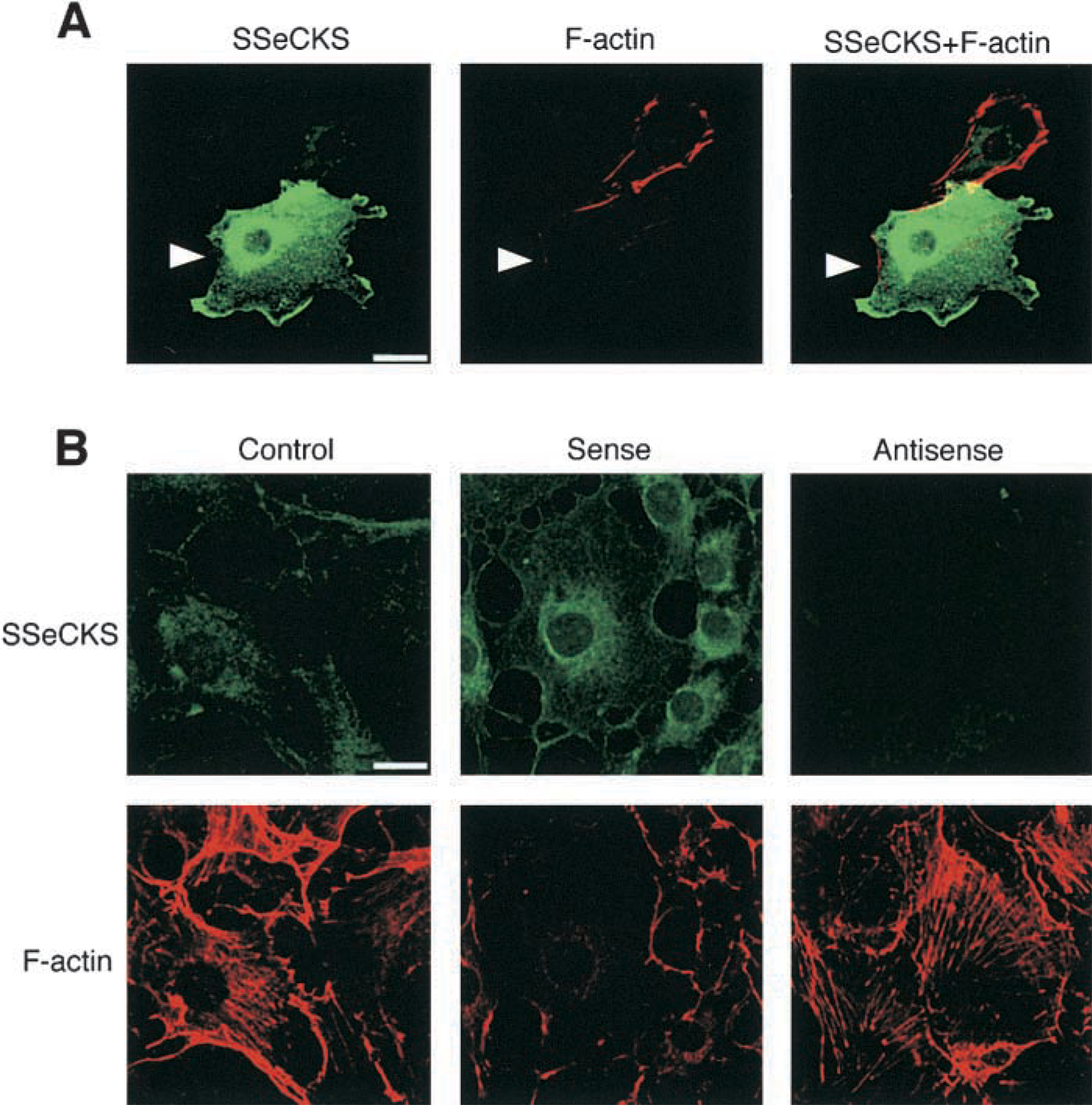
Effects of ectopic expression of SSeCKS on F-actin fibers. (
In the present study we failed to detect any increase of SSeCKS mRNA and protein in LEII cells, even after LPS stimulation. The paucity of SSeCKS induction by LPS may be attributable to unusual characteristics of this immortal and proliferative cell line. Alternatively, LPS itself might not induce SSeCKS, and endogenous humoral factors released from non-endothelial cells stimulated by LPS might participate in SSeCKS induction. One of the candidate factors is tumor necrosis factor (TNF), which is a vasoactive cytokine induced in macrophages in response to LPS (Camussi et al. 1991; Goldblum et al. 1993). In preliminary experiments we observed that TNF significantly increased SSeCKS mRNA expression in the mouse lung and also in LEII cells. Further studies are needed to clarify the role of TNF in SSeCKS induction in LPS-induced inflammation.
Rodent SSeCKS shows a significant structural homology with human gravin (Nauert et al. 1996), which was originally identified as an autoantigen of myasthenia gravis (Gordon et al. 1992). The N-terminal 1000 amino acids are 69% identical and the C-terminal 30 residues are ~80% similar (Nauert et al. 1996). Gravin is selectively expressed in adherent cultured cells, and associated with the cytoskeleton in intracellular localization, suggesting a role for gravin in cell motility and adhesion (Gordon et al. 1992). Gravin is also present in human umbilical vein endothelial cells and in restricted types of endothelia in the normal baboon, i.e., hepatic sinusoidal endothelial cells and epithelia of intestinal lacteals (Gordon et al. 1992; Grove et al. 1994). Moreover, a recent fluorescence in situ hybridization analysis suggested that gravin may be a human orthologue of SSeCKS (Xia et al. 2001). It would therefore be interesting to examine the expression of gravin in human subjects under inflammatory conditions.
Footnotes
Acknowledgements
Supported in part by grants-in-aid from the Ministry of Agriculture, Forestry and Fisheries (RCP-2000–4230) and from the Ministry of Education, Culture, Sports, Science and Technology of Japan (12760191).
We are grateful to Dr I. H. Gelman for his kind gift of the antiserum against rat SSeCKS. We are also grateful to Dr A. S. G. Curtis for providing LEII cells.