
Abstract
We examined the cellular distribution of glutathione transferase A4 (GSTA4) in various human tissues by indirect immunoperoxidase using a specific polyclonal antibody raised in rabbit. This enzyme was localized in hepatocytes, bile duct cells, and vascular endothelial cells in liver, upper layers of keratinocytes and sebaceous and sweat glands in skin, proximal convoluted tubules in kidney, epithelial cells of mucosa and muscle cells in colon, muscle cells in heart, and neurons in brain. Staining was increased in pathological situations such as cirrhosis, UV-irradiated skin, and myocardial infarction and was strongly decreased in hepatocellular carcinoma. These results strongly support the view of a close correlation between cellular GSTA4 localization and the formation of reactive oxygen species in the tissues investigated.
G
One of the major GST classes, the alpha class expressed in most organs, comprises at least four genes encoding hGSTA1, A2, A3, and A4, as well as several pseudogenes. The most recently discovered, hGSTA4, is believed to be involved in tissue and cell defense against oxidative stress through its high activity for 4-hydroxynonenal and related endogenous electrophiles. In contrast to other GST isoenzymes, the cellular localization of GSTA4 has not been investigated in human tissues. Only studies performed on a few rat or mouse tissues have led to the conclusion that GSTA4 is expressed in epidermis and sebaceous glands of dermis (Hiratsuka et al. 1999), in smooth muscle cells of the vascular media of the aorta (Misra et al. 1995), and in astrocytes (Johnson et al. 1993).
To further characterize hGSTA4, we have analyzed its cell distribution in several human tissues using the immunohistochemical (IHC) technique. For this purpose, we first raised a polyclonal antibody against a recombinant peptide derived from an mGSTA4 peptide sequence, which specifically reacted with GSTA4. Our results show that, compared to other GSTs, GSTA4 has a unique cellular distribution in the different human tissues studied, and support the view of its potent role in the elimination of lipid peroxidation products.
Materials and Methods
Materials
The QIAexpressionist kit was used to obtain high-level expression and purification of 6xHis-tagged peptide GSTA4 (Qiagen; Courtaboeuf, France). The in vitro transcription and translation kit was purchased from Promega (Madison, WI). The immunopure immobilized protein A used for immunoprecipitation experiments was from Pierce (Rockford, IL). The secondary antibody was peroxidase-conjugated, affinity-purified goat anti-rabbit IgG (BioRad; Hercules, CA).
Tissue Samples
Human liver, skin, colon, kidney, and heart samples were obtained from the department of surgery, Hopital Pontchaillou (Rennes, France). Brain samples were obtained from the neuropathology department [Hopital Pitié-Salpetriere, Paris (Pr. Hauw)]. Access to biopsy material was in agreement with French laws and satisfied the requirements of the local Ethics Committee. After macroscopic examination, tissue fragments were fixed in 4% formalin for 12–24 hr and embedded in paraffin for routine histopathological diagnosis. A portion of fresh liver was also snap-frozen in isopentane, cooled in liquid nitrogen, and stored at — 80C until protein extraction. Fresh frozen normal and UV-B-exposed (4.13 mV/cm2; 0.503 J/cm2; 5 min) skin and normal colon tissues were obtained from Biopredic International, (Rennes, France).
Production of Recombinant Protein mGSTA4, Antibody, and Immunoprecipitation
The mouse GSTA4 peptide (amino acids 106–175) was subcloned into pQE10 vector (Qiagen) in frame with the 6-histidine residue tag. The construct was transferred into E. coli M15 (pREP4) host cells in standard Luria Broth medium supplemented with 100 μg/ml ampicillin and 25 μg/ml kanamycin at 37C. When optical density purity reached 0.7, protein expression was induced by adding isopropyl thio-β-
The antiserum was purified by binding of GSTA4 antibody to the recombinant mGSTA4 peptide immobilized on nitrocellulose membrane according to protocols described by Harlow and Lane (1988). Antibody fixed on the nitrocellulose membrane was eluted with 100 mM glycine (pH 2.5), immediately neutralized with 1 M Tris-HCl (pH 8, 10v/1v), and frozen for further use. Concentration of antibody was 0.3 mg/ml.
The purified antibody was tested for its specificity by its ability to immunoprecipitate murine and human GSTA4, but not other GSTs. mGSTA4, hGSTA4, hGSTA1, hGSTP1, and hGSTM1 cDNA were generated by reverse transcription polymerase chain reaction (RT-PCR) using total RNA isolated from mouse or human liver. Sequencing of the GST cDNA products obtained demonstrated identity to the corresponding GST sequences previously published (DeJong et al. 1988; Dixon et al. 1989; Hubatsch et al. 1998; Tu and Qian 1986; Zimniak et al. 1992). Each cDNA was cloned into the pCl-neo mammalian expression vector (Promega). In vitro transcription and translation (IVTT) of these GST isoforms were performed using the translation-coupled reticulocyte lysate system (Promega) and [35S]-methionine (Amersham; Arlington Heights, IL) as described by the manufacturer. The labeled IVTT products were diluted 1:50 with lysis buffer (Tris 50 mM, pH 8, NaCl 50 mM, EDTA 5 mM, MgCl2 15 mM, NP40 0.5%, and extemporaneously 1 mM DTT, 0.1 mM PMFS, 10 μg/ml leupeptin, and 10 μg/ml aprotinin) and incubated with the antibody (1.5 μg) for 2 hr at 4C. Immune complexes were bound to protein A-Sepharose beads, washed three times in lysis buffer, then recovered and denatured with sample buffer (Laemmli 1970). Proteins were resolved in 12.5% SDS-PAGE. Gels were fixed in ethanol 70%, acetic acid 25%, and H2O 5%, and dried [35S]-methionine-labeled proteins were revealed by autoradiography. The antibody specificity was also tested by Western blotting using the GST IVTT products.
Protein Extraction and Western Blotting Analysis
Biopsy fragments were homogenized in Western blotting lysis buffer (Hepes 50 mM, pH 7.5, NaCl 150 mM, EDTA 1 mM, EGTA 2.5 mM, 0.1% Tween-20, 10% glycerol, 10 mM β-glycerophosphate, 1 mM NaF, 0.1 mM orthovanadate sodium, 0.1 mM PMFS, 10 μg/ml leupeptin, and 10 μg/ml aprotinin). Proteins (100–150 μg/well) were fractionated by SDS-PAGE (12.5% gel), and transferred to a PVDF membrane. GSTA4 was detected by immunoblotting with the GSTA4-specific polyclonal antibody.
Immunohistochemistry
The indirect immunoperoxidase method was used to localize GSTA4 in paraffin-embedded sections. Sections were deparaffinized and blocked for nonspecific binding of antibody with PBS containing 3% bovine serum albumin for 30 min and then covered with a solution of anti-rabbit GSTA4 antiserum (1:100) for 1 hr at room temperature. After three rinses with PBS, the sections were incubated for 2 hr in a solution containing goat anti-rabbit IgG conjugated to peroxidase (1:200). The sections were rinsed, stained with diaminobenzidine, and counterstained with hematoxylin, except for liver and brain sections, which were developed with aminoethylcarbazole.
Control reactions included incubation with purified immunoglobulins of the preimmune rabbit serum (diluted at 1:100 in PBS) and then with peroxidase-anti-rabbit immunoglobulins and direct reaction with the staining reagent.
Results
Production and Characterization of a Mouse GSTA4 Antibody
Homology between two members of the alpha class is high. To obtain a specific antibody against GSTA4, a region of the mGSTA4 sequence (Zimniak et al. 1992) presenting a high homology with hGSTA4 (Desmots et al. 1998; Hubatsch et al. 1998), but the lowest homology with other GST members was selected. The most appropriate domain was found to lie between amino acids 106 and 175 using BLAST search software. The recombinant 6x-His mGSTA4 fusion protein was produced in E. coli, purified under denatured conditions using nickel beads, and injected into rabbits.
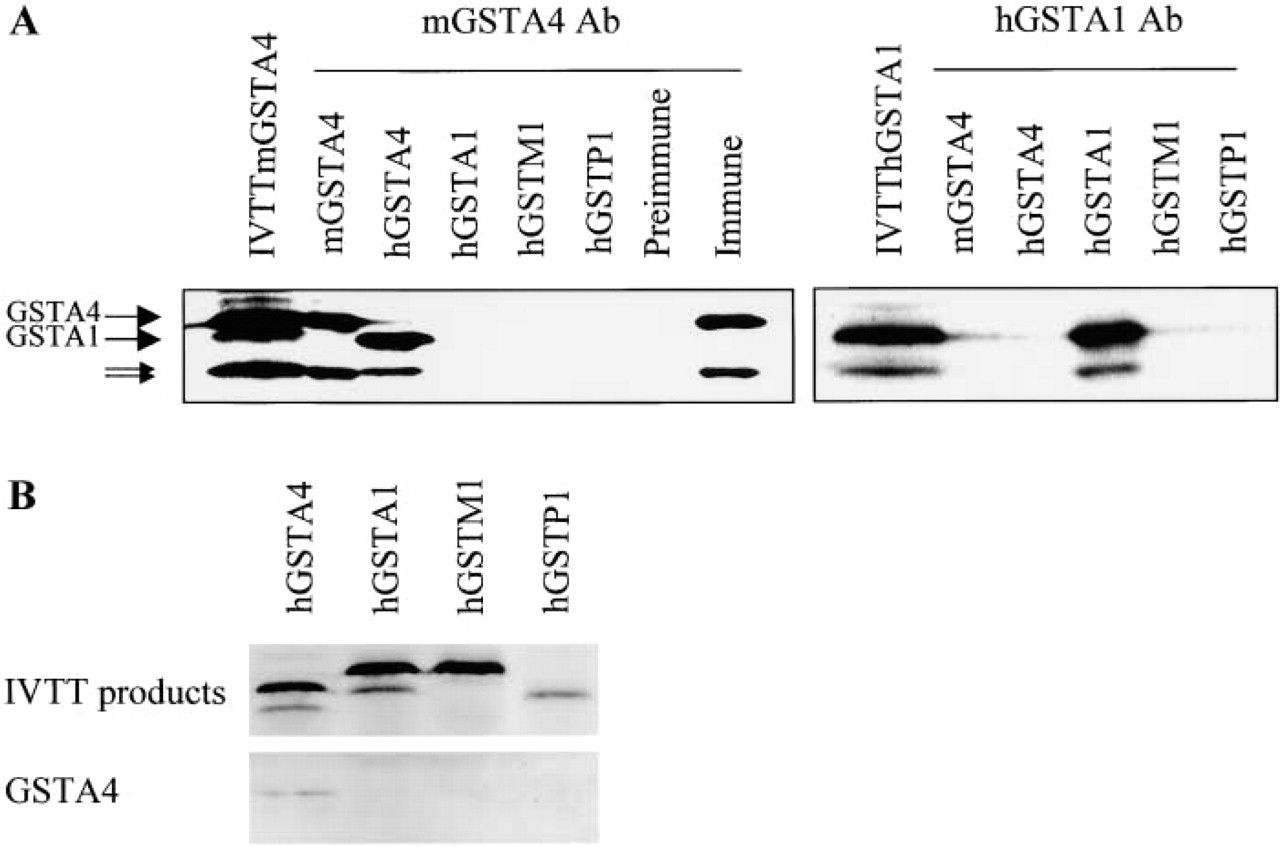
(
The purified anti-GSTA4 antibody was demonstrated to specifically immunoprecipitate GSTA4 and did not crossreact with other GST isoforms, i.e., GSTA1, M1, and P1 of either mouse or human origin (Figure 1A). This antiserum immunoprecipitated both m- and hGSTA4 IVTT products. Used as a control, preimmune serum of the same rabbit did not immunoprecipitate the mGSTA4 IVTT products. Similarly, by Western blotting, the anti-GSTA4 antibody recognized IVTT products of mGSTA4 (data not shown) and hGSTA4, but not those corresponding to human GSTA1, M1, and P1 (Figure 1B). A single band was also detected in whole-cell extracts of mouse, human, and rat liver with the mGSTA4 antibody (Figure 2). Reactivity of mouse and human GSTA4 was found to be markedly different.
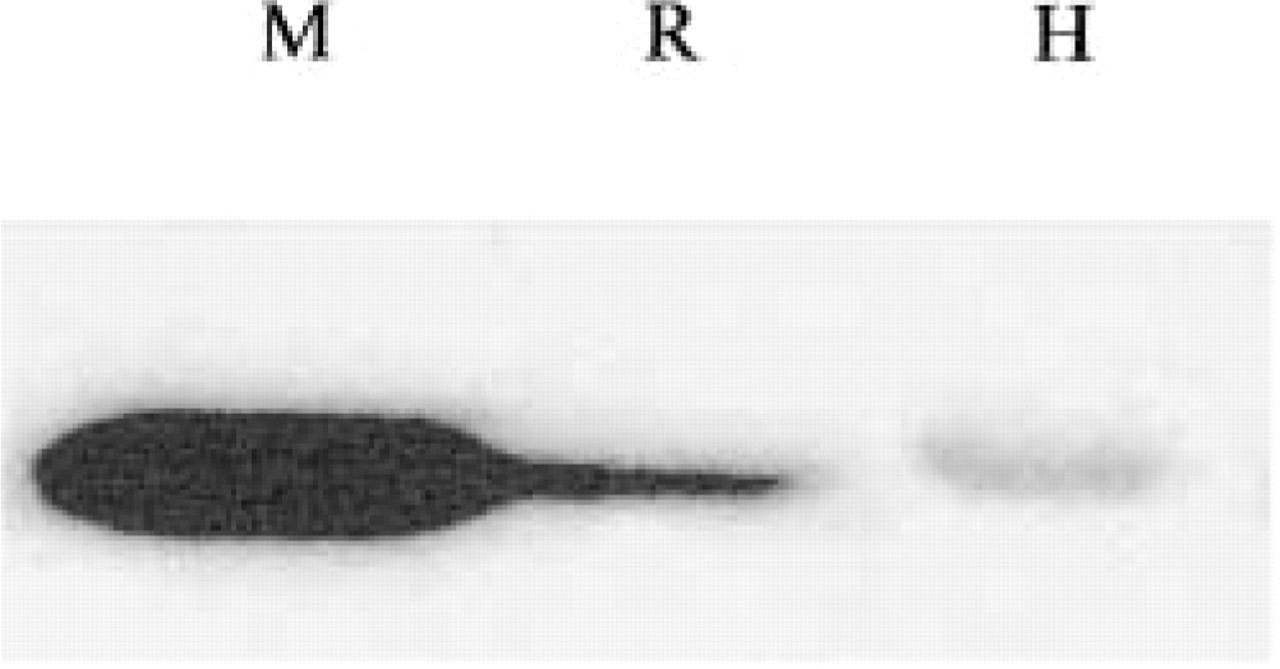
Western Blotting analysis of GSTA4 expression in mouse (M), rat (R), and human (H) liver.
Western Blotting Analysis
GSTA4 expression in liver, skin, and colon was demonstrated by Western blotting (Figure 3). Some variations were observed between normal and pathological conditions. Our data indicate that GSTA4 was expressed in normal tissues and pathological conditions such as fibrosis, whereas its expression was strongly decreased in hepatocellular carcinoma. GSTA4 expression in skin was markedly enhanced (1.8-fold) when the skin was exposed to UV-B light. GSTA4 expression was also demonstrated in two normal colon samples.
Immunohistochemical Study of GSTA4 Expression
Sections from liver, skin, kidney, colon, heart, and brain tissues were examined. Fifteen liver samples, including eight normal tissues, two cirrhosis, three hemochromatosis, and two hepatocellular carcinoma were analyzed. In normal liver GSTA4 was detected in most hepatocytes, exhibiting a heterogeneous intralobular distribution. Staining was usually more intense in periportal than in centrilobular parenchymal cells and was restricted to the cytoplasm (Figure 4A). Moreover, inter-individual variations were observed in the intensity of hepatocyte immunostaining for GSTA4. Bile duct cells and vascular endothelial cells were also positive. In pathological tissues, except for tumor cells that exhibited low staining (Figure 4B), GSTA4 appeared to be expressed at variable levels and no preferential intralobular distribution could be visualized. Most cirrhotic hepatocytes were strongly stained.
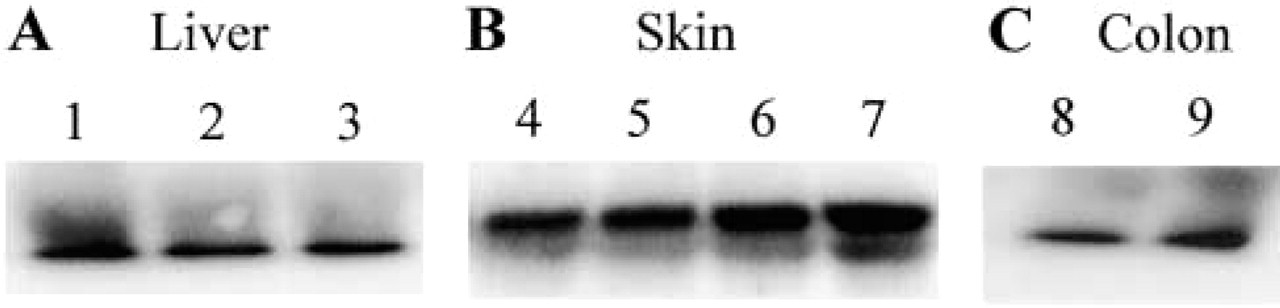
Western blotting analysis of GSTA4 in human samples. (
Four normal skin samples and two of basocellular carcinoma were observed. In normal tissues, GSTA4 staining was predominantly found in the upper layer of keratinocytes (Figure 4C) and the outer sheath of the hair follicles in the epidermis, as well as in sebaceous and sweat glands and vessels of the dermal compartment (Figure 4D). GSTA4 was also evident in basocellular carcinoma cells, being more intense at the periphery of the two tumors studied, both originating from light-exposed skin.
In the six normal kidney samples studied, proximal convoluted tubules in the renal cortex, glomeruli, and thick loops of Henle were labeled with GSTA4 antibodies, whereas distal convoluted tubules were unstained (Figure 4E).
In normal colon tissue, strong staining was visualized in columnar and crypt epithelial cells. The four samples were similarly labeled. In addition, muscle cells of muscular mucosa and vessels of submucosa were also stained (Figure 4F).
One normal heart and two myocardial infarction specimens were analyzed. Normal muscle cells of the myocardium were strongly and selectively stained with the GSTA4 antibody (Figure 4G) and still more intensely labeled in the two myocardial infarction tissues.
Three normal brain and two Alzheimer disease (AD) tissues were examined. The perinuclear area of neurons as well as vessels were well stained (Figure 4H) and no obvious difference in the intensity of staining was observed among the different samples.
Discussion
A polyclonal antibody specifically reacting with hGSTA4 and not with other GST alpha members was raised in rabbit. A major difference in reactivity between the mouse and human GSTA4 was observed. This could be related to the percentage of homology in amino acids between mGSTA4 and hGSTA4 or rGSTA4, i.e., 65% and 86%, respectively, and/or to the different quantities of GSTA4 in the mouse and human liver samples tested.
The specific GSTA4 antiserum was used to examine cellular distribution of this GST isozyme in six different human tissues, i.e., liver, skin, kidney, colon, heart, and brain. In normal liver, GSTA4 was found in the cytoplasm of hepatocytes, bile duct cells, and vessels. GSTA4 staining was frequently more intense in periportal than in centrilobular normal hepatocytes. By contrast, cytochromes P450 are usually more expressed in centrilobular parenchymal cells (Ratanasavanh et al. 1991), and other GST alpha enzymes do not exhibit heterogeneous intralobular location (Hiley et al. 1988). The preferential periportal expression of GSTA4 could be related to higher peroxidase activity in this lobular area, in agreement with the role of this isozyme in detoxification of lipid peroxidation products such as hydroxyalkenals (Kera et al. 1987). The hepatic periportal zone is recognized to be implicated in the initial breakdown of fatty acids and to show a higher rate of fatty acid oxidation, which is associated with generation of reactive oxygen species (ROS) (Gebhardt 1992). The intracytoplasmic GSTA4 staining appears to be punctate, supporting the observations of Bhagwat et al. (1998) that this enzyme is partly located in mitochondria. By using confocal microscopy, we have confirmed a cytosolic and mitochondrial localization of GSTA4 in mouse hepatocytes (unpublished results). Moreover, inter-individual variations were observed in the intensity of hepatocyte staining, suggesting differences in the level of GSTA4 expression, as usually observed for xenobiotic metabolizing enzymes.
The distribution of GSTA4 in other normal human tissues also appeared to be related to its role in the degradation of lipid peroxidation products. Thus, in normal skin GSTA4 was mainly expressed in the superficial layers of the epidermis and in the glands of the dermis. This preferential localization of GSTA4 in the upper keratinocyte layers is probably related to their involvement in elimination of lipid peroxidation products after induction of oxidative stress by exogenous factors. This interpretation is supported by the findings that GSTA4 expression was induced in rodent (Hiratsuka et al. 1999) and human skin (this study) after UV irradiation.
The detection of GSTA4 in convoluted proximal tubules of the renal cortex and in columnar and crypt epithelial cells of the colon is also compatible with the occurrence of an oxidative stress induced by chemicals in these cells. Their localization in their respective tissues places these cells in contact with various chemicals. Interestingly, we recently found that GSTA4 was increased in proximal convoluted tubules after iron overload (unpublished data), which is known to be associated with active formation of ROS (Harley et al. 1993; Houglum et al. 1997). The localization of GSTA4 in myocardial muscle cells and in neurons is also in agreement with a production of lipid peroxidation products in these target cell types for toxic compounds.
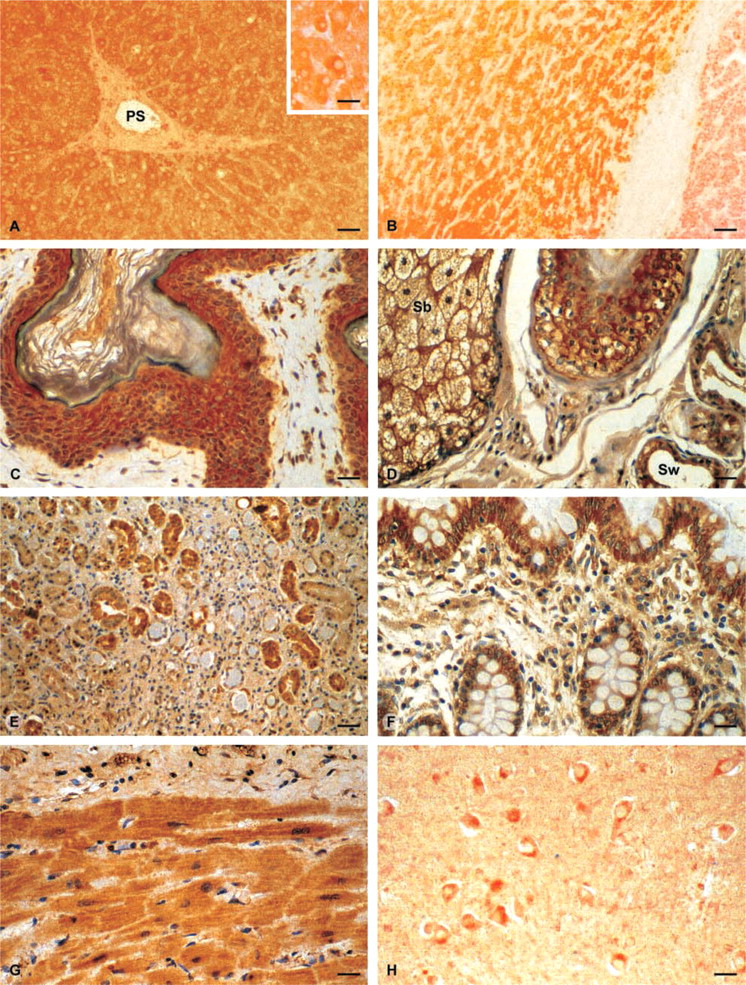
Distribution of GSTA4 in different human tissues. (
GSTA4 distribution only partly overlaps with that of other alpha class GSTs and mu and pi GSTs (Table 1). For example, in the skin, GSTA1, the major alpha class member enzyme, is not expressed, whereas GST9.9, another alpha class enzyme, is located only in sweat glands (Raza et al. 1991; Moral et al. 1997). GSTPi, the major isozyme in the skin, is found in the stratum basalis and, to a lesser extent, in superficial keratinocyte layers and sweat glands. This unique localization of GSTA4 enzyme may be explained by its major role in the elimination of lipid peroxidation products. The increased GSTA4 staining in pathological situations, such as hepatic fibrosis, cirrhosis, hemochromatosis, and myocardial infarction, also supports this conclusion. In these diseases, inflammation and cell alterations result in an increased production of ROS. By contrast, GSTA4 expression was strongly diminished in primary hepatocarcinoma. This finding agrees with recent reports showing that activities of antioxidant enzymes such as superoxide dismutase are reduced in both human hepatocellular carcinoma and in the N1S1-transplanted rat hepatoma, and favors the conclusion that these tumor cells produced fewer ROS (Liaw et al. 1997; Sung et al. 1999). Similarly, the high GSTA4 expression in the superficial cell layers of basocellular carcinoma is compatible with the production of free radicals in these cells, which remained exposed to UV (Chan and Yu 2000).
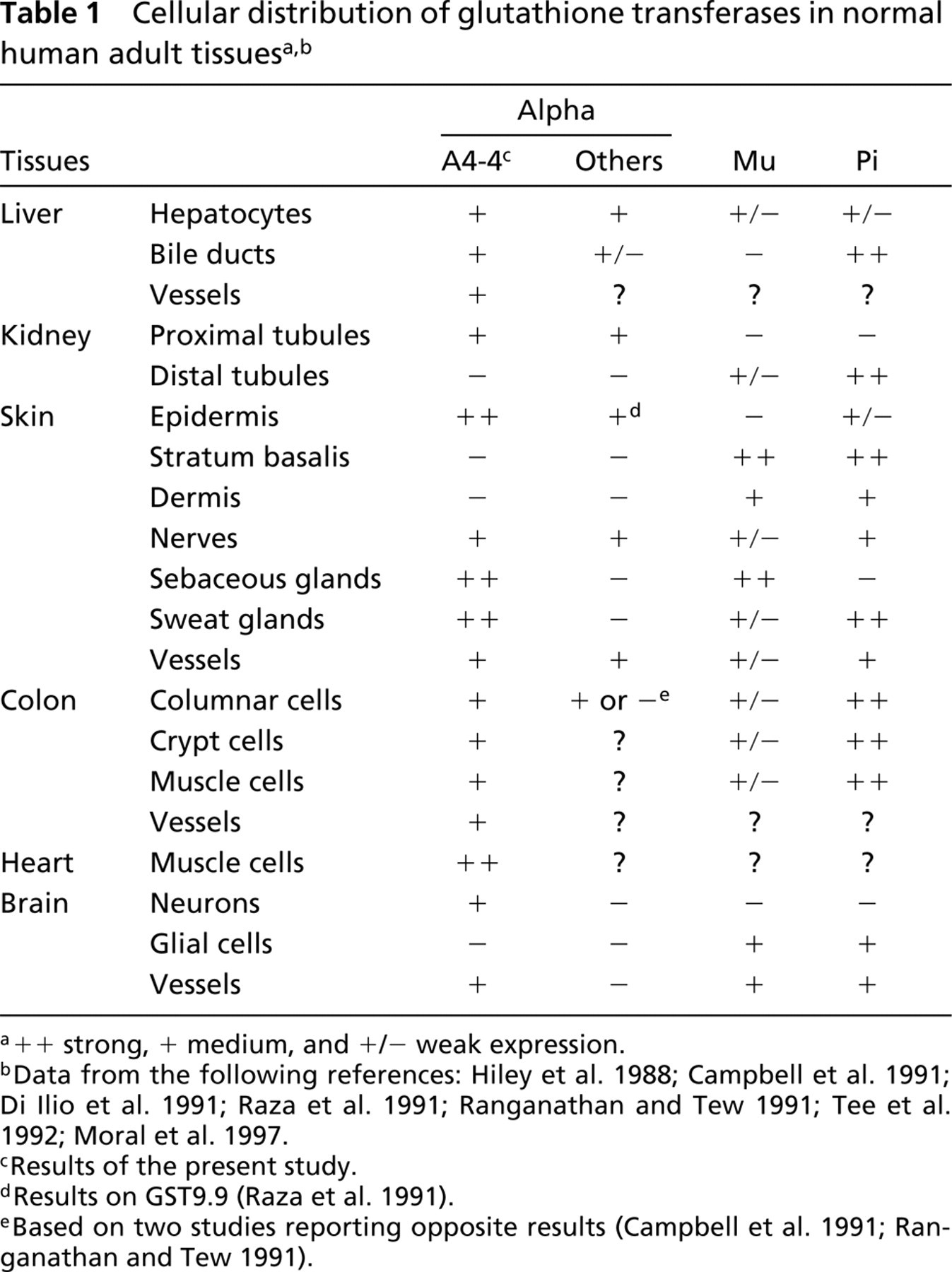
++ strong, + medium, and +/− weak expression.
Data from the following references: Hiley et al. 1988; Campbell et al. 1991; Di Ilio et al. 1991; Raza et al. 1991; Ranganathan and Tew 1991; Tee et al. 1992; Moral et al. 1997.
Results of the present study.
Results on GST9.9 (Raza et al. 1991).
Based on two studies reporting opposite results (Campbell et al. 1991; Ranganathan and Tew 1991).
In conclusion, using a specific antibody against GSTA4, we determined for the first time the cellular localization of this enzyme in various human tissues. Our data are in favor of a close correlation between GSTA4 distribution and the production of ROS in the tissues studied.
Footnotes
Acknowledgments
Supported by the Institut National de la Santé et de la Recherche Médicale and the Association France Alzheimer.
We thank Orlando Musso for his helpful collaboration.