
Abstract
Liposomal antigens are potent adjuvants of humoral and cell-mediated immunity. Although this property requires as an essential condition a physical association between the antigen and the phospholipid vehicle, the nature of the association, i.e., encapsulation or surface linkage, markedly influences the outcome of the elicited response. Available evidence suggests that macrophages are involved in this fine tuning of the immune response in a manner that is not yet clearly established. It is postulated that this might be related to their capacity to interact differently with surface-linked and encapsulated formulations. Using conalbumin as a model antigen, we address the question by analyzing the movements of encapsulated and surface-linked antigen as well as those of MHC-II molecules in macrophages in a pulse-chase immunoelectron microscopic study carried out over a 24-hr period. The antigen was followed using a polyclonal serum specifically raised against fragmented conalbumin (fCA) that allows the detection of processed antigen and of some MHC-peptide complexes. The results indicate that, in macrophages, the two liposomal formulations affect macrophage morphology in distinct ways and circulate through the various subcellular compartments with different kinetics. On the basis of the overall results, we conclude that surface-linked antigen gains access less readily to the endogenous presentation pathway than encapsulated antigen but can favor a more sustained activation of the immune system through its production of exosome-like structures and its more thorough utilization of the MHC-II pathway.
D
These observations on the functional duality of surface-linked and encapsulated antigens suggest that liposomal formulations have access to specific activation pathways from the earliest steps of their penetration of the immune network. This further suggests that professional antigen-presenting cells (APCs), including dendritic cells (DCs), B-cells, and macrophages, could be differently involved in the overall process through their capacity to differentially take up, process, and present liposomal antigens. Although the capacity of all three types of APCs to interact with liposomal antigens has been documented (Dal Monte and Szoka 1989; Nair et al. 1992; Alving 1993), macrophages are still considered to be the main cell type involved. In support of this are the observations that macrophages are the main cell targets of liposomes in vivo and that their elimination almost totally abrogates li-posomal adjuvanticity (Su and Van Rooijen 1989; Nair et al. 1995). Although this conclusion has been derived mainly from studies carried out with encapsulated material, in the rare instances where the mode of antigen association was analyzed, the importance of macrophages was also recognized (Dal Monte and Szoka 1989; Su and Van Rooijen 1989). However, the role of macrophages can hardly be considered exclusive because macrophages have a limited capacity to stimulate naive T-helper cells (Steinman 1991; Swain et al. 1996) and because their elimination does not similarly affect the response to encapsulated and surface-linked antigens (Su and Van Rooijen 1989; Guan et al. 1998). Available evidence more strongly suggests that macrophages are involved in the fine tuning of the immune response to liposomal antigens through their specific treatment of the different formulations. By allowing a different routing of encapsulated and surface-linked antigens after internalization, macrophages might favor specific cytokine influences and privileged contribution from other APCs. Preliminary data indicate that this might in fact be the case (Fortin et al. 1996). Here we present a more complete pulse-chase immunoelectron microscopic study that analyzes the fate of both types of liposomal antigens and the movements of MHC Class II molecules in macrophages over a 24-hr period. As a detection tool, we used a polyclonal serum specifically raised against conalbumin fragments (fCA) which allows the identification of processed antigens and of some MHC-pep-tide complexes.
Materials and Methods
Animals
Male and female BALB/c mice (haplotype H-2d) and New Zealand rabbits were obtained from Charles River Canada (St Constant, QC, Canada). Mice were used at 8–12 weeks of age.
Liposomes
Liposomes made of dimyristoyl phosphatidylcholine, cholesterol, and dipalmitoyl phosphatidylethanolamine (DPPE) in a molar ratio of 63:31:6 were prepared by extrusion as previously described (Fortin et al. 1996), using polycarbonate filters with pore size of 0.2 μm. Lipids, solubilized in chloro-form:methanol (9:1), were rotoevaporated to dryness under a nitrogen atmosphere. The lipid film was then resuspended in 10 mM HEPES, 0.9% NaCl, pH 7.4 and incubated for 2 hr at 37C before extrusion. Conalbumin was either encapsulated in the course of liposome formation or surface-linked by a disulfide linkage to pre-formed liposomes containing DPPE initially modified with the heterobifunctional reagent 3-(2-pyridyldithio) propionic acid N-hydroxysuccinimide ester (SPDP) according to the method described by Leserman et al. (1984). Liposome-associated antigen was separated from free conalbumin by two ultracentrifugation steps (30 min, 145,000 × g) carried out at 10C. The efficiency of association was evaluated by γ-counting using radioiodinated protein.
Rabbit Polyclonal Anti-fCA Antibodies
Antibodies to fCA obtained by the CNBr method were raised in rabbits. Each rabbit received three doses of 100 μg of fCA emulsified in complete (first dose) or incomplete (second and third doses) Freund's adjuvant and injected SC at 4-week intervals. The immune serum was purified by affinity chromatography on CNBr-activated Sepharose using fCA as ligands. The antiserum was tested by ELISA and Western blotting against native CA and fCA as well as against unrelated proteins such as ovalbumin, bovine serum albumin, and lysozyme. By ELISA, it was shown to be highly specific for fCA, although in Western blots the native form was also recognized. The capacity of the antiserum to detect MHC-II-peptide complexes was demonstrated by double-labeling confocal immunofluorescence microscopy as well as by Western blotting of anti-Iad immunoprecipitates of whole-cell lysates according to the method of Davidson et al. (1991).
Macrophage Preparation and Sensitization
Macrophages were elicited by IP injection of 1 ml of 20% proteose peptone (Becton-Dickinson Microbiology Systems; Cockeysville, MD). Three days later the peritoneal exudate was recovered in cold RPMI 1640, washed three times, and plated in 35-mm Petri dishes at a density of 5.2 × 106 cells/ml (1.3 ml/dish) in RPMI supplemented with 0.05 mM mercaptoethanol and 20 U/ml rIFN-γ (Pharmingen; Mississauga, ON, Canada). The cells were left to adhere for 24 h at 37C in a humidified atmosphere enriched with 5% CO2. Adherent cells were pulsed for 15 min with 10 μg of either encapsulated or surface-linked CA (50 μg protein/μmol phospholipid) in 1 ml of RPMI. After the pulse, the cells were washed and chased at 37C for various times in RPMI (1 ml/dish).
High-resolution Immunocytochemistry
At the end of the chase period, Petri dishes were put on ice. Cells were washed twice with ice-cold Dulbecco's PBS, pH 7.4, and then fixed for 2 hr at room temperature with 1% glutaraldehyde in 0.1 M cacodylate buffer (pH 7.2) containing 1% sucrose. After three washes with 0.1 M cacodylate buffer (pH 7.2) containing 4.5% sucrose, cell monolayers were left overnight at 4C in the same buffer. Fixed cells were recovered from Petri dishes with a cell scraper and centrifuged for 8 min at 7000 × g in the sucrose-cacodylate buffer. The pellets were resuspended in 200 μl of 2% liquid agar in cacodylate buffer and immediately centrifuged for 5 min at 16,000 × g. The pellets were then cut into 1-mm3 pieces, postfixed for 1 hr in the dark with 1% tannic acid in 0.05 M cacodylate buffer, pH 7.0, partially dehydrated in ethanol, and infiltrated with LR White resin (“hard grade”'; Pelco, Redding, CA).
Thin sections (90 nm) were mounted on 300-mesh unsupported nickel grids and processed for immunogold labeling. Cell sections were etched for 30 min with 0.1% aqueous sodium m-periodate. Free aldehydes were blocked for 15 min with 0.1 M glycine in 0.05 M Tris, 0.15 M NaCl, pH 8 (TBS). Free sites and Fc receptors were blocked by 1% fish gelatin, 0.01% nonimmune decomplemented mouse serum in TBS for 15 min. Sections were double-labeled on separate sides to detect processed fCA and mouse MHC-II molecules (Iad). Mouse monoclonal anti-Iad (Pharmingen) and rabbit anti-fCA were used at a concentration of 40 μg/ml in TBS containing 0.1% OVA-0.05% fish gelatin. Gold-labeled goat anti-mouse IgG and goat anti-rabbit IgG were then used for detection with particles of 15 and 10 nm, respectively (British Biocell International; Cardiff, UK). Sections were finally contrasted with 4% aqueous uranyl acetate followed by Reynold's lead citrate and examined with a Philips PW 6006/00 transmission electron microscope. For each time point, 16 cells cut at similar cell levels and exhibiting minimal background staining were randomly selected. Gold particles present in different compartments of the selected cells were counted on micrographs.
To assess the specificity of the labeling reaction, the following immunocytochemical controls were carried out: labeling of macrophages incubated in the absence of liposomal antigens; treatment of sections with nonimmune serum or with buffer before incubation with labeled secondary antibody; and extinction of the reaction by preincubation of the specific antibodies with fCA.
Results
Immunocytochemical Analysis
For the purpose of immunoelectron microscopic analysis, the following subcellular compartments were considered: the cytosol, the Golgi, the plasma membrane, and different internal vesicular structures subdivided into three groups on the basis of morphological criteria. Vesicles containing little or no electron-dense material and mainly recovered at the cell periphery were called clear vesicles (CVs). They included membrane rufflings as well as membrane invaginations such as macropinosomes and early endosomes. Vesicles containing dispersed electron-dense material were named granular vesicles (GVs). These vesicles, usually larger than CVs, were more regularly circular and found deeper in the cytosol. They were assumed to represent more mature endosomal compartments, such as late endosomes and early lysosomes. Vesicles filled with electron-dense material and recovered mainly in the vicinity of the nucleus were named dense vesicles (DVs). These vesicles were considered to be lysosomes.
Effect of Liposomal Antigens on Macrophage Morphology
Encapsulated antigen and surface-linked antigen were both shown to influence significantly, although differently, the morphology of macrophages throughout the 24-hr chase period (Figures 1 and 2). After sensitization with surface-linked antigen, many CVs and GVs of similar dimensions initially formed at one pole of the cell, some perinuclear DVs being visible and the intervesicular spaces being filled with a relatively abundant cytosol (Figure 1A). This pattern evolved rapidly towards the networking of CVs, the enlargment as well as the decreased granulosity of GVs, and the almost total disparition of DVs (Figures 1B and 1C).
During the same chase period, the morphological aspect of macrophages sensitized with encapsulated antigen remained relatively stable (Figures 1D-1F). At all times however, membrane ruffling was much less intense than observed with surface-linked antigen, the cytosolic compartment was more developed, and the endocytotic pathway mainly dominated by large GVs of low granulosity, CVs being only occasionally seen.
The differences between the two liposomal formulations became more striking during the last phase of the chase spanning the period of 3–24 hr. In response to surface-linked antigen (Figures 2A-2C), the cytosolic compartment almost completely disappeared, CVs and GVs occupying most of the cellular space. Whereas CVs were mainly organized as an interconnected convoluted network, GVs containing only a few remnants of granular material were present as large, circular, individualized structures. With time, the vesicular structures segregated in specific cellular areas, GVs occupying a central region completely surrounded by the compact, convoluted CV network (Figure 2C).
With encapsulated antigen, a CV network of significant importance became apparent only after 3 hours of chase (Figure 2D) but, compared to that observed in response to surface-linked antigen, it was more loosely organized. Large GVs were almost as abundant as CVs and were recovered more deeply into the cell than observed at earlier times. With time (Figures 2E and 2F), GVs become more granular and the CV network, although still loosely organized, more convoluted. Throughout the chase period, the cytosolic compartment remained well developed.
Intracellular Trafficking of Encapsulated Antigen
After sensitization of macrophages with the encapsulated formulation, CA was detected from the earliest time of the chase in all the vesicular structures examined and, in all these compartments, it was recovered either free in the lumen or close to the vesicular membrane.
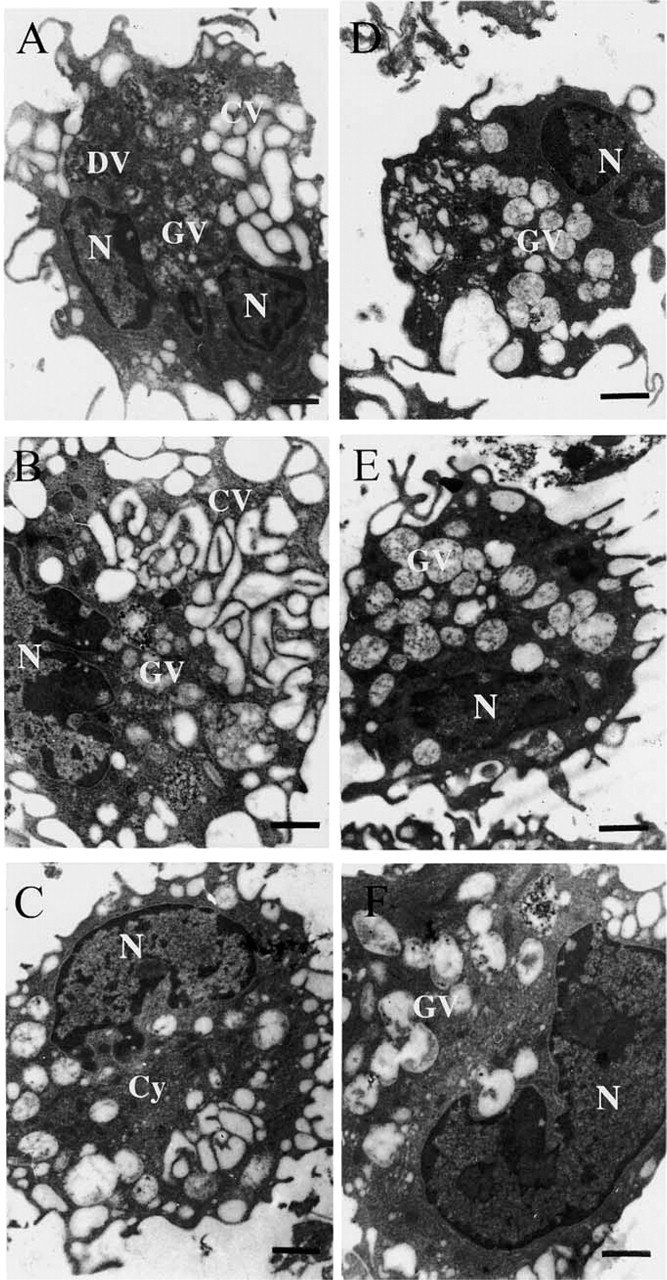
Effect of liposomal antigens on macrophage morphology. Macrophages pulsed for 15 min with surface-linked (
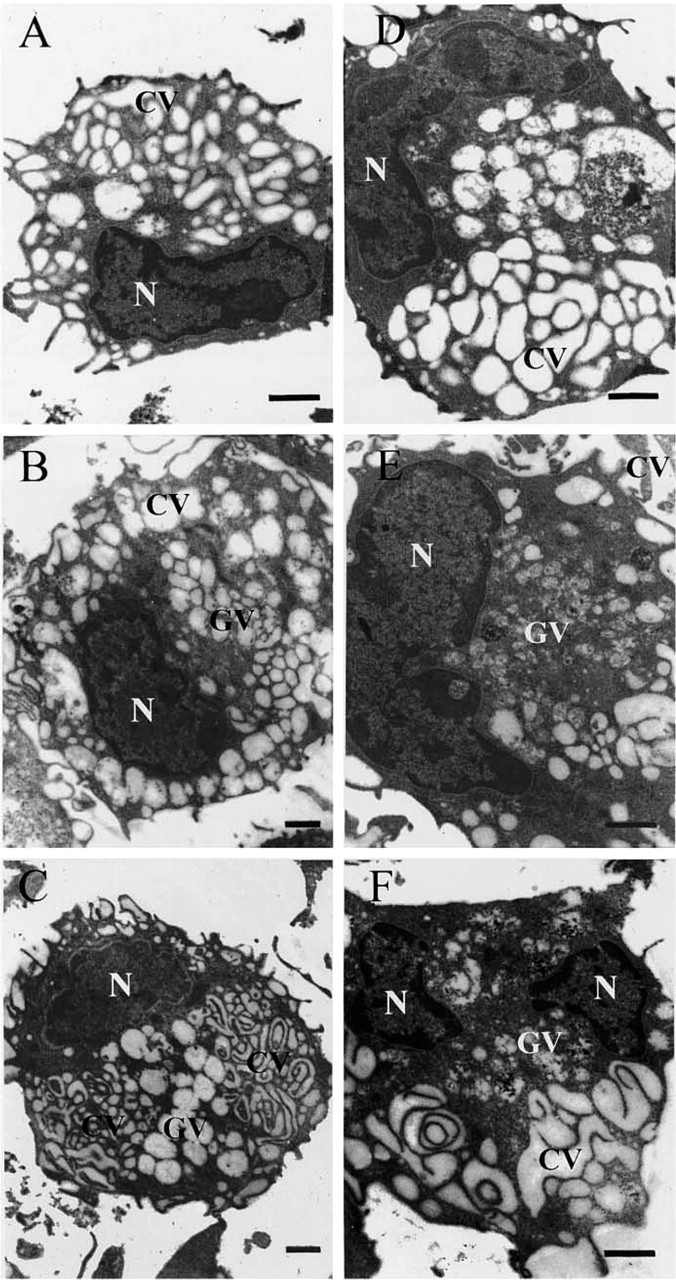
Effect of liposomal antigens on macrophage morphology. Macrophages pulsed for 15 min with surface-linked (
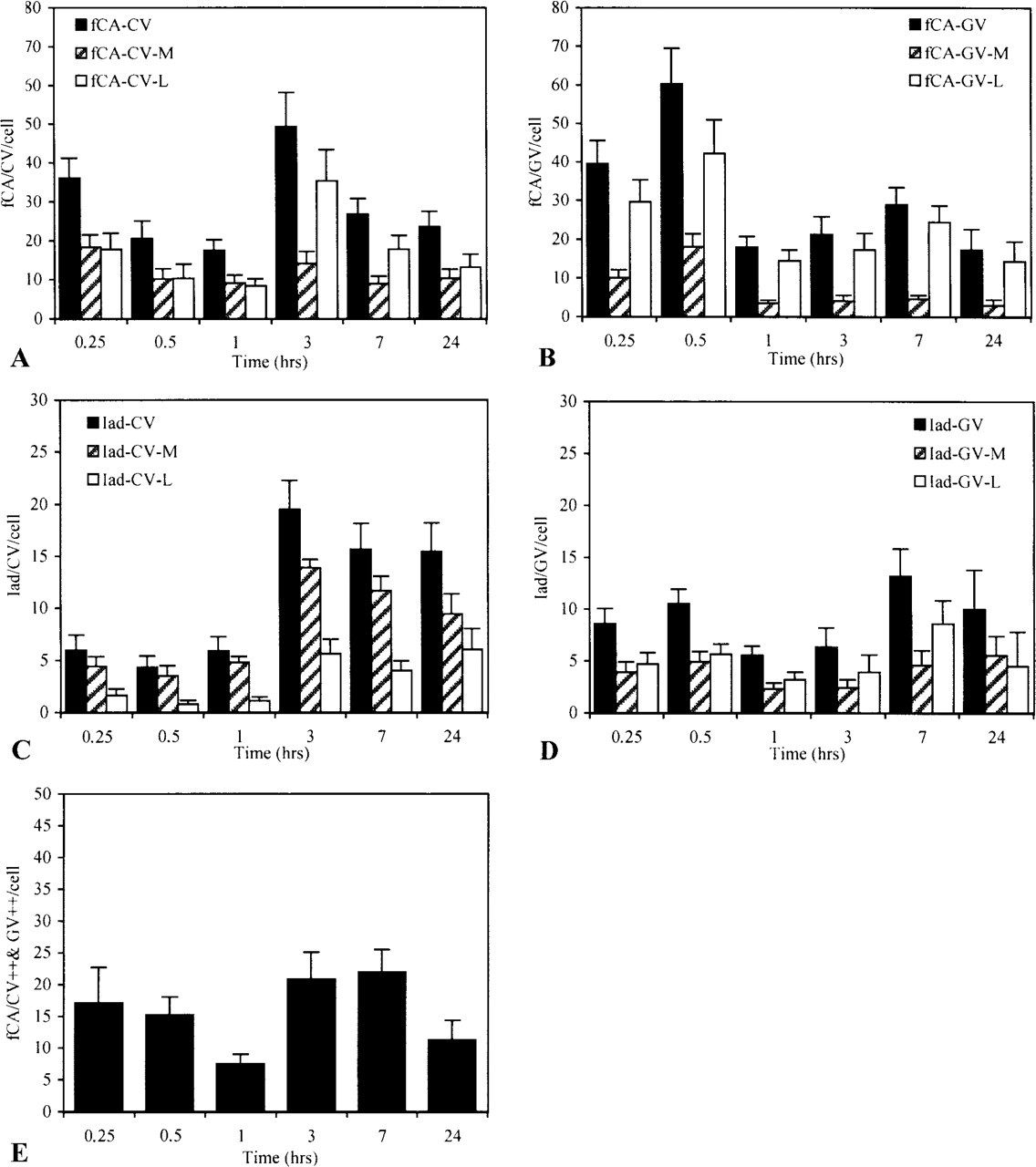
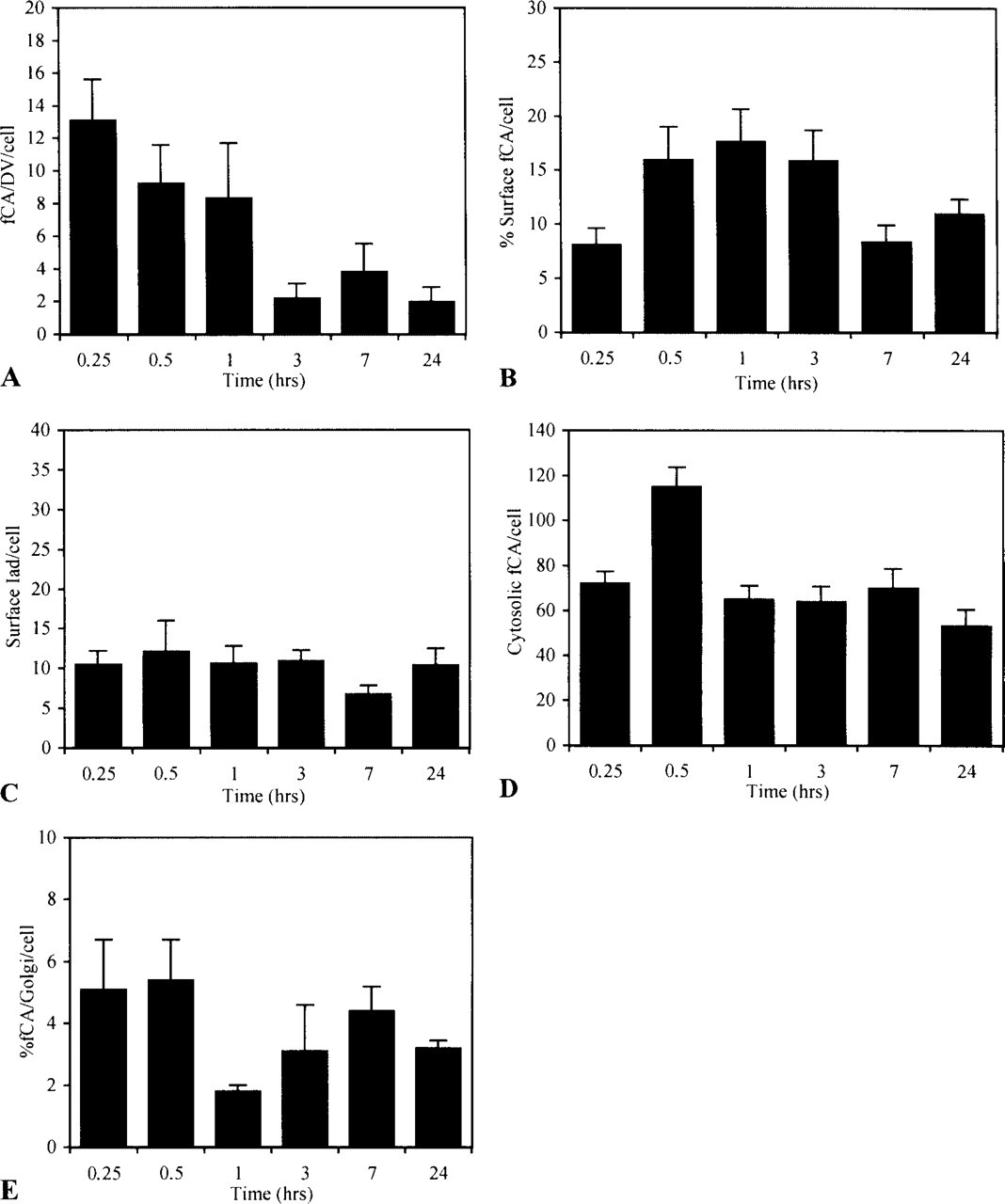
Intracellular trafficking of encapsulated antigen and Iad molecules. Macrophages pulsed for 15 min with encapsulated antigen were chased for various periods of time in RPMI. At each time, cells were fixed and analyzed for the presence of conalbumin fragments (fCA) and Iad molecules in CVs and in GVs. For each time point, 16 cells were analyzed. (
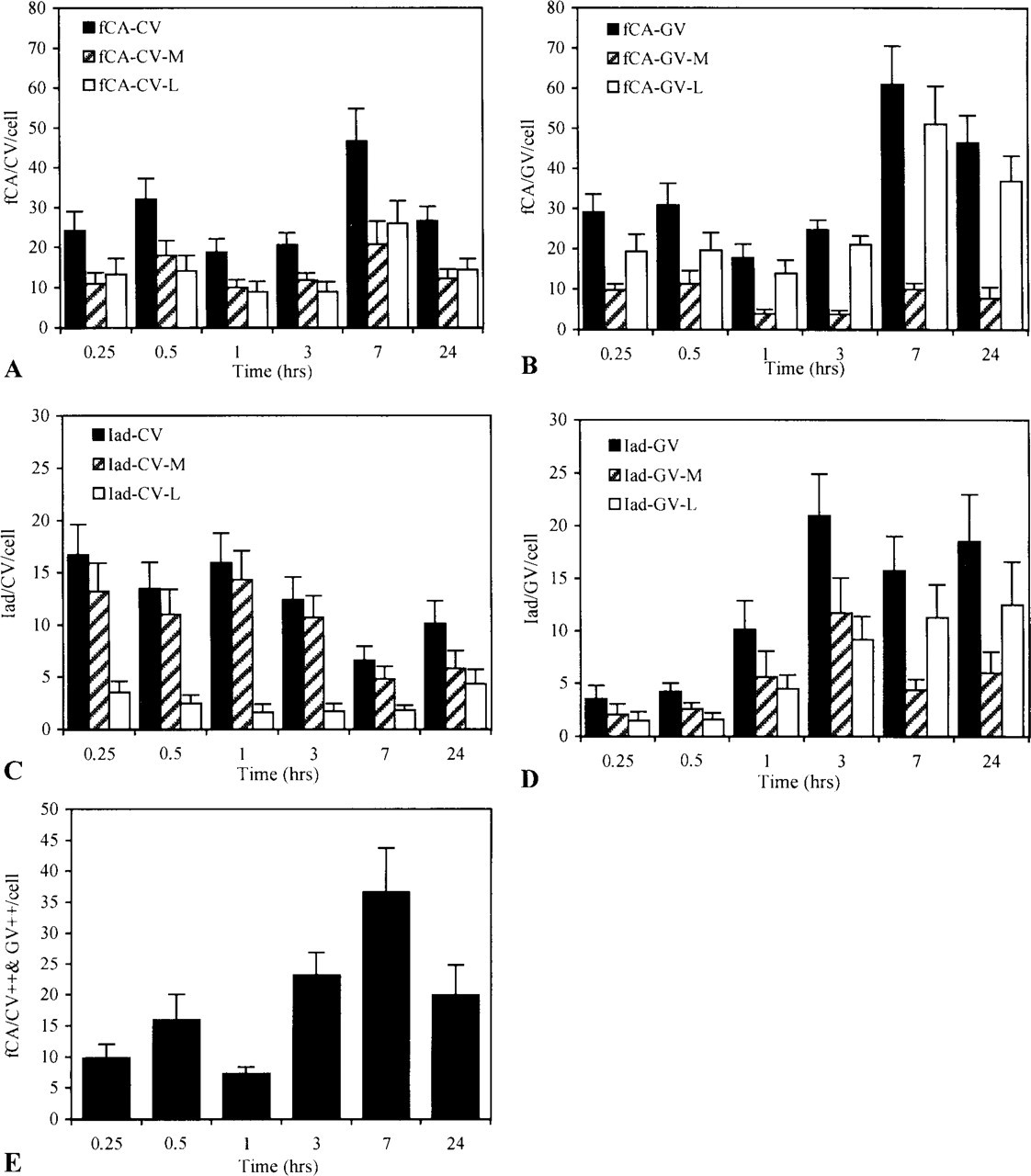
In CVs, two peaks of CA were detected (Figure 3A) and both forms of processed antigen, the luminal and membrane forms, were detected throughout the kinetics, although in different ratios. Whereas the membrane form was detected at a constant level throughout the chase period, the luminal form largely dominated the second wave, accounting for most of the increase observed at 3 hr. Under this form, CA could be liberated into the external milieu as demonstrated by ELISA analysis of sensitized macrophages supernatants (data not shown). The movements of CA in CVs were correlated with opposite movements in GVs (Figure 3B). However, in those vesicles the antigen was essentially recovered under the luminal form.
In both CVs and GVs, processed CA could associate with MHC-II products because Iad molecules were also detected in these compartments (Figures 3C and 3D). Like conalbumin, Iad was recovered in two forms: a membrane form, representing more probably the functional form of the molecule, and a luminal form, which we considered as a breakdown product of the whole molecule. Whereas the membrane form was the dominant form in CVs, the two forms were detected in almost equivalent proportion in GVs throughout the chase period. Strangely, at 3 hr, an abrupt and large increase in the detection of Iad was observed in CVs. The increase correlates in time with a similar increase at the plasma membrane (Figure 4C). It was also observed in GVs, but later on and with less intensity.
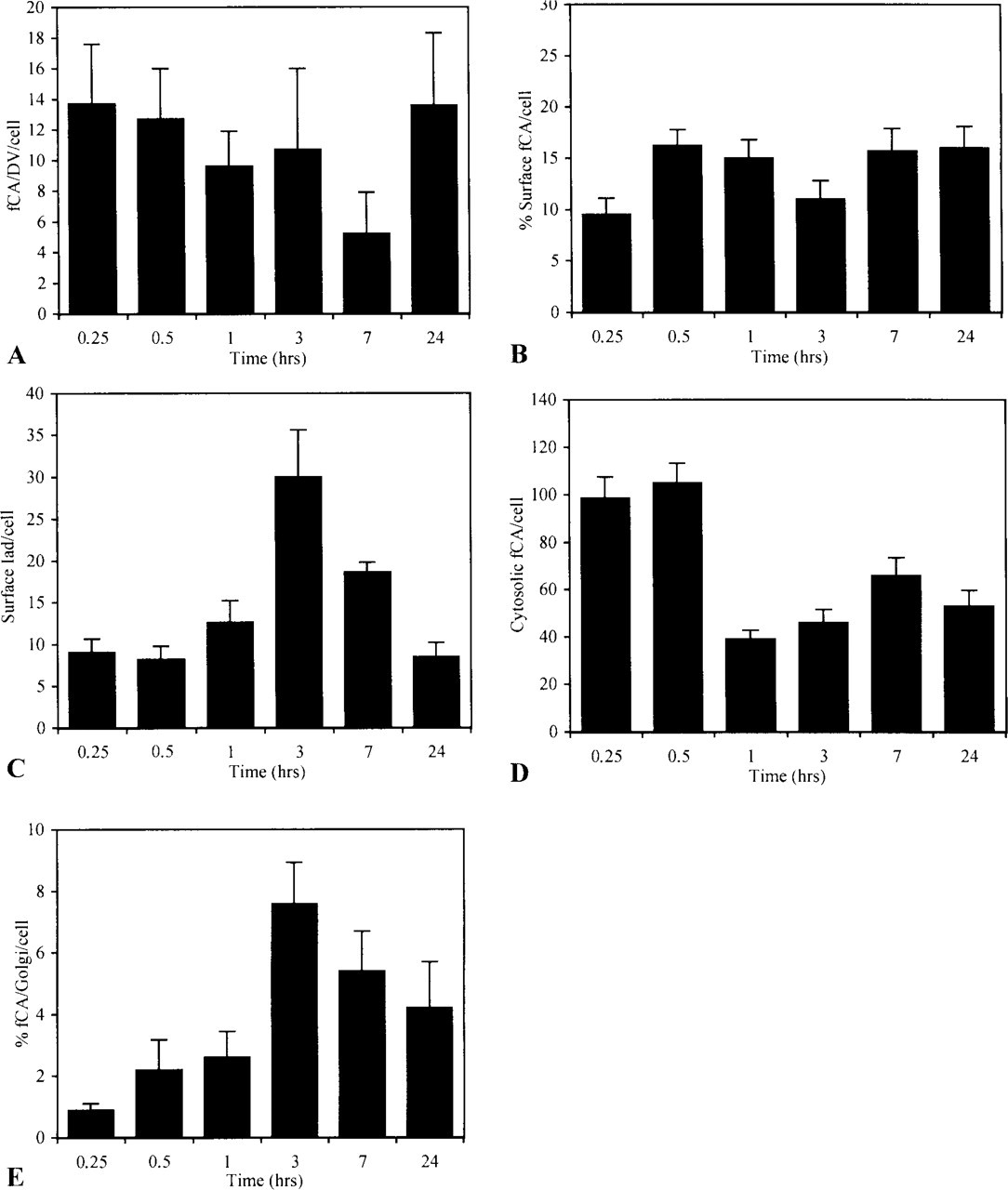
Intracellular trafficking of encapsulated antigen. Macrophages pulsed for 15 min with encapsulated antigen were chased in RPMI for various periods of time. At each time, cells were fixed and analyzed for the presence of fCA in different cellular compartments. For each time point, 16 cells were analyzed. (
Iad and CA did not circulate through the endocytotic pathway with identical kinetics but their capacity to interact should be closely related to their simultaneous identification at the same place and at the same time. Although it was exceptional to detect Iad in close proximity to fCA (Figure 5), both were nevertheless often shown to co-localize (Figure 3E). During the chase, two peaks of CA were in fact identified in Iad-containing vesicles: a rapid one already maximal at the beginning of the chase and a late one of almost similar intensity occuring at 3–7 hr of chase.
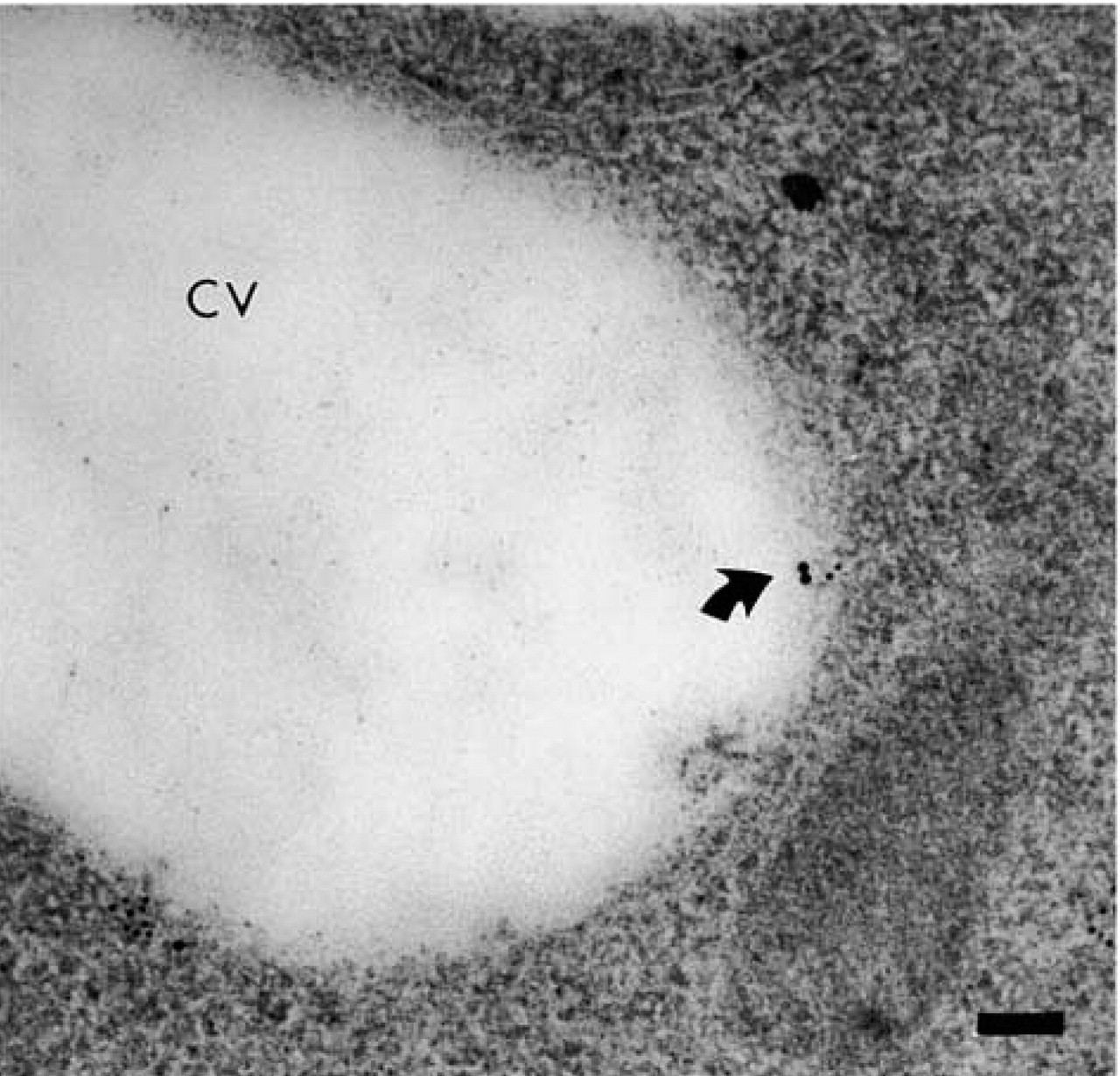
Peptide-MHC complexes. A peptide-MHC complex formed between Iad and fCA (arrow) recovered at the membrane of a clear vesicle (CV) after 3 h of transit. fCA was detected with 10-nm gold particles and Iad with 15-nm gold particles. Bar = 0.1 μm.
Conalbumin was also rapidly detected in DVs, the end of the line of the endocytotic pathway (Figure 4A). Labeling was already maximal at the earliest time of the chase and did not change significantly over time.
From the endocytotic pathway, fCA gained access to the plasma membrane in two waves of similar intensity (Figure 4B): a rapid one of short duration, which peaks at 30 min, and a slow, long-lasting one that reaches its maximum at 7 hr. Iad, as expected, was also detected at the plasma membrane (Figure 4C).
Despite the acid resistance of our liposomal formulation, which is supposed to restrict the release of antigen into the cytosol (Harding et al. 1991), encapsulated CA was shown to rapidly escape the endocytotic pathway (Figure 4D). This rapid release was mainly related to fusion events occurring at the time that liposomes were still intact, as supported by the many figures of liposomes stuck to the endosomal membrane or in the process of fusing that we observed at the earliest times of the kinetics (Figure 6A and 6B). At later times, a second peak of cytosolic CA was observed, indicating that some fragmented peptides generated into the endocytotic pathway after destabilization of the liposomal vehicle also escaped into the cytosol. From the cytosol, encapsulated antigen reached the exocytotic pathway, where association with MHC-I products could take place, as indicated by the presence of fCA over the Golgi apparatus in a time frame consistent with the overall process (Figure 4E). However, the kinetics also indicated that only the first wave of cytosolic antigen reaches the Golgi to a significant extent.
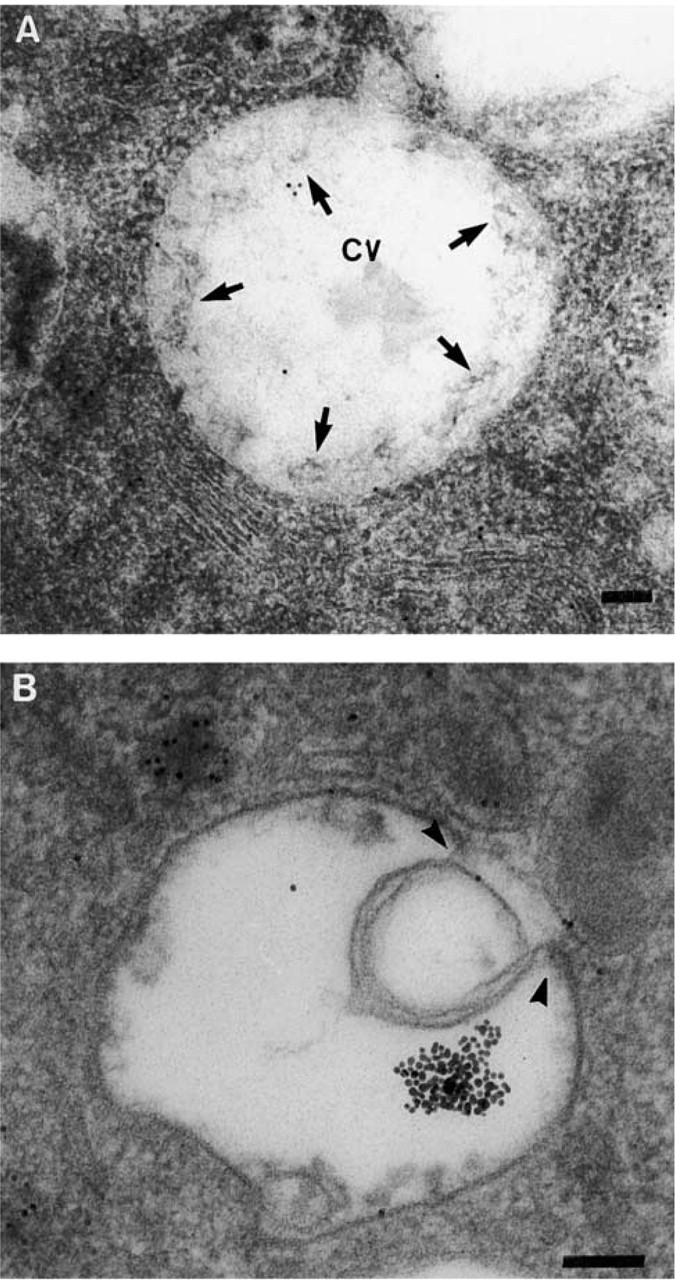
Fusion of encapsulated liposomal antigen with endosomal membranes. (
Intracellular trafficking of surface-linked antigen and Iad molecules. Macrophages pulsed for 15 min with surface-linked antigen were chased in RPMI for various periods of time. At each time, cells were fixed and analyzed for the presence of fCA and Iad molecules in CVs and in GVs. For each time point, 16 cells were analyzed. (
Intracellular Trafficking of Surface-linked Conalbumin
Like encapsulated antigen, surface-linked antigen was internalized by macrophages through macropinocytosis and peptides generated from the exposed antigen were rapidly detected in all cellular compartments, although with different kinetics. In CVs, CA was detected in two successive waves: a rapid one that peaks at 30 min and a slower but more intense one that peaks at 7 hr (Figure 7A). Both the membrane and the luminal forms were represented but, in contrast to what was observed with encapsulated antigen, the two forms were recovered at a constant ratio throughout the kinetics. More interestingly, the luminal form was not found exclusively free in the vesicular space but was often associated with intraluminal membranous structures that were particularly abundant at the latest times of the chase (Figure 8B). The movement of CA in GVs (Figure 7B) was correlated with that observed in CVs, the second wave being also the most important. In these GVs, the luminal form was always the dominant form, although it was present in increasing proportion with time. MHC-II products were shown to circulate through these compartments according to their own kinetics. While in CVs, Ia was present at its highest level during the first chase hour (Figure 7C); the opposite was true in GVs (Figure 7D). Moreover, whereas in CVs the largely predominant form of Iad was the membranous form, the luminal form was always largely represented in GVs. In both compartments, the luminal form was detected in increasing proportions with time but, in contrast to what was observed with encapsulated antigen, this late luminal form was often found as part of membranous luminal structures with which fCAs were also associated (Figure 8B). In all these movements, the co-localization of fCAs and Iad molecules followed two phases, one of low intensity occurring early during the chase, with a maximum at 30 min, and a second one much more intense, with its maximum at 7 hr of chase (Figure 7E).
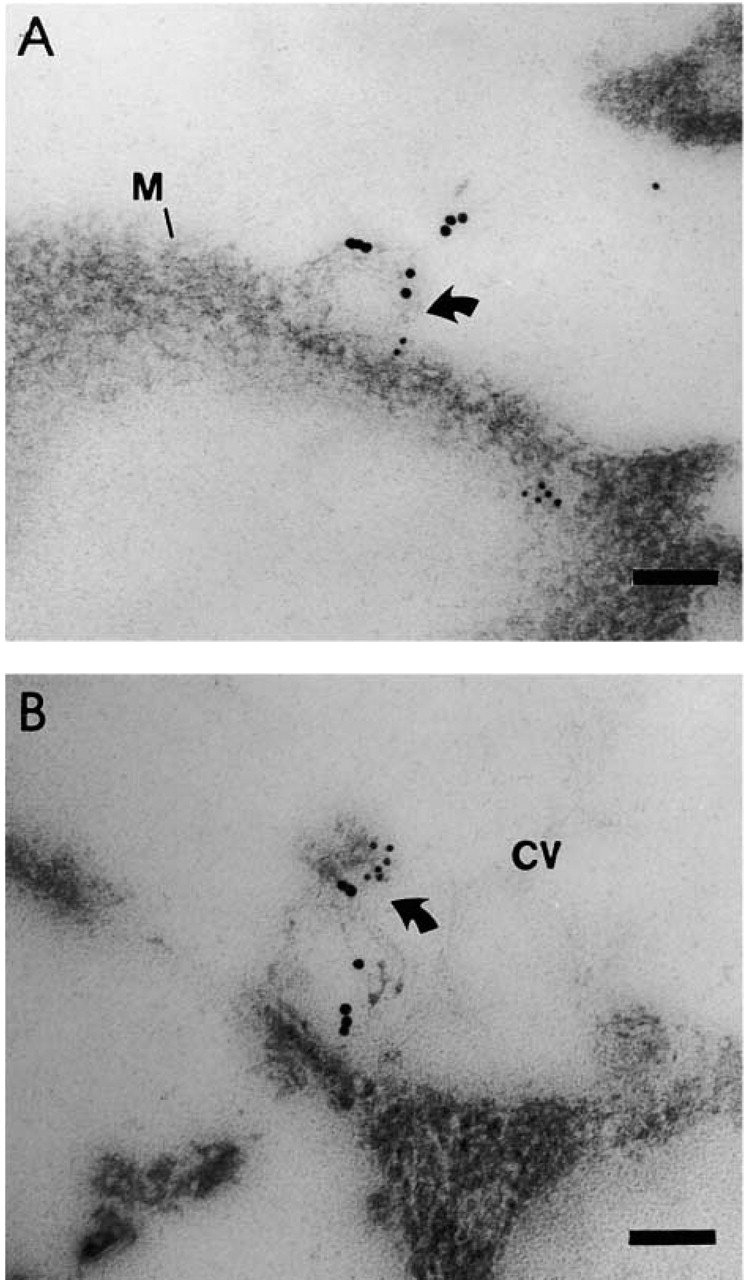
Luminal and extracellular membranous vesicles associated with surface-linked antigen. (
Like encapsulated antigen, surface-linked antigen was rapidly seen in DVs (Figure 9A) but, in contrast to what was observed with the encapsulated formulation, this labeling was rapidly lost.
At the membrane level (Figure 9B), the detection of fCAs increased rapidly during the first hour of chase, then slowly declining after 3 hr, whereas that of Iad remained constant during the entire chase period (Figure 9C). Membranous structures labeled for both Iad and CA, and similar to those identified in the lumen of CVs and GVs, were often seen in close proximity to the plasma membrane (Figure 8A).
Surface-linked CA also escaped the endocytotic pathway for accessing the cytosol (Figure 9D). However, the leakage appeared to be more gradual than that observed with encapsulated antigen and, according to our electron microscopic observations, did not involve fusion of liposomes with the vesicular membranes. Apparently, few of these cytosolic CA fragments were rescued by the cytosolic processing pathway for presentation in the context of MHC-I molecules, as suggested by the negligible amount of antigen detected over Golgi structures in a time frame consistent with the processing and loading events (Figure 9E). Unexpectedly, however, surface-linked antigen was detected in the Golgi at the earliest times of the kinetics.
Discussion
Our immunoelectron microscopic study highlights the very specific interactions that can be established between macrophages and encapsulated or surface-linked antigens. This is seen both in the specific morphological influences induced by the two liposomal formulations on macrophages and in the different kinetics of their intracellular trafficking. This observation lends support to the initial hypothesis concerning the involvement of macrophages in the fine tuning of the immune response to liposomal antigens.
With the simultaneous detection of MHC-II products and processed antigens, the presentation pathways preferentially used by the two liposomal formulations could be analyzed. The unique properties of the polyclonal antibodies specifically developed against fCA were of major importance in this respect because they allow not only detection of the processed antigen but also of some of the MHC-peptide complexes that could be formed after processing. Despite its intrinsic advantages, our detection tool has also some limitations that must be considered in analyzing the data and that probably explain the low level of detection observed in some cell compartments. For example, it does not necessarily recognize the same antigenic forms throughout the kinetics, and its detection capacity is expected to be lost on complete degradation of the antigen. Moreover, only a fraction of the relevant MHC-peptide complexes can more probably be identified and the simultaneous detection of Iad and pep-tide on the same complex is more certainly generally impeded as a consequence of steric hindrance. Taking these factors into consideration, our data nevertheless lead to the conclusion that surface-linked antigen utilizes more thoroughly than encapsulated antigen the various possibilities of the MHC-II pathway and compensates for its lower access to the endogenous pathway by the production of exosome-like structures.
Intracellular trafficking of surface-linked antigen. Macrophages pulsed for 15 min with surface-linked antigen were chased for various periods of time. At each time, cells were fixed and analyzed for the presence of fCA in different cellular compartments. For each time point, 16 cells were analyzed. (
Effects on Macrophage Morphology
One of the most significant and striking effects of encapsulated and surface-linked antigen on macrophages is their long-lasting and specific influences on cell morphology. These observations, although they may in part result from the use of IFNγ-activated macrophages, contradict the general belief that the adjuvant character of liposomes derives mainly from their ability to passively and rapidly transport optimal amounts of antigen to APCs (Alving et al. 1992; Gregoriadis 1994; Van Rooijen 1995). The results more clearly indicate that encapsulated and surface-linked antigen actively transform macrophages and induce some unique yet unidentified cellular activities. These may involve the production of inflammatory cytokines, such as induced by bacterial lipids or lipopeptides (Rouaix et al. 1994; BenMohamed et al. 1997; Schmidt et al. 1997), or the induction of danger signals now considered so crucial to the development of an immune response (Matzinger 1994). Interesting in this respect is the observation that the expression of heat shock proteins recognized as potent danger signals (Chen et al. 1999; Todryk et al. 1999) can be be modulated by various lipophilic substances or alterations in membrane fluidity (Zhang et al. 1994; Vigh et al. 1998).
Fate of Liposomal Antigens
Conalbumin, whether it is encapsulated or surface-linked, rapidly reaches all compartments of the endocytotic pathway, in agreement with the results of Racoosin and Swanson (1993). Although processed in these compartments, the antigen appears to be in great part protected from total degradation because a high level is still detected after 24 hr of chase. The protection can scarcely be attributed to a rescue of generated peptides by MHC-II molecules because most of the antigen recovered at the latest times of the chase is found as a luminal form rather than as a membrane-associated form. However, the intense membrane movements initiated in macrophages by IFN-γ and exacerbated by liposomes might be important in this respect by favoring the back-and-forth movement of peptides through the endosomal compartments, thus impeding their too long residence in lysosomes and their ensuing total degradation. Heat shock proteins have also been proposed to protect peptides from extensive degradation and might therefore also contribute to maintaining the presence of antigen into the endosomal pathway (Panjwani et al. 1999).
At the end of the kinetics, large amount of luminal CA accumulates in the CV network where exchanges with the external milieu are believed to take place, suggesting that the pre-processed antigen can be released from the cell and possibly taken up by more efficient APCs. The identification of fCA in the supernatant of sensitized macrophages (data not shown) supports this conclusion and agrees with the regurgitation hypothesis already proposed by others (Van Rooijen 1993; Gong et al. 1994; Harding and Song 1994; Nair et al. 1995). Our results indicate, however, that encapsulated and surface-linked antigens are released in the external milieu in different forms. Whereas encapsulated antigen is essentially regurgitated as a free soluble form, some of the surface-linked antigen is released as part of small vesicular structures with which Iad molecules are also associated. Although of slightly larger diameter, these structures resemble exosomes, the 60- to 80-nm internal vesicles of the MHC-II compartment, which have been shown to accumulate a number of cellular proteins involved in immunity, such as MHC-II products and heat-shock proteins or integrins (Thery et al. 1999), and to participate in several immune processes including immunological memory, self-tolerance, T-cell activation, and cell-mediated cytotoxicity (Raposo et al. 1996; Zitvogel et al. 1998; Arnold and Mannie 1999; Brenner and Wainberg 1999).
The presence of a significant amount of CA in the cytosol throughout the chase indicates that the endocytic pathway initially followed by exogenous antigen, such as liposomes, is not as hermetic as initially believed. Although leakage apparently occurs at almost all levels along the path, the massive efflux initially observed identifies early endosomal compartments as a major source of cytosolic antigen. Macropinosomes in which liposomes are internalized might be more specifically involved because they have been shown to be especially leaky when loaded with particular antigens (Germain et al. 1996). Once in the cytosol, the antigen can reach the MHC-I presentation pathway. However, the differences observed in the intensity and the kinetics of labeling over Golgi-like structures suggests that only the earliest molecules delivered into the cytosol access this pathway to any significant extent. Because association with MHC-class I products requires that peptides of specific length and molecular characteristics be generated by the proteasome and transferred into the reticulum lumen by the TAP-1 and TAP-2 proteins (York et al. 1999), endosomal processing might impede cytosolic processing so that only intact or lightly processed antigen can be usefully rescued in the cytosol. In line with this proposal is the observation that surface-linked antigen, which is already accessible to endosomal enzymes, is not as easily transferred to the Golgi as its encapsulated counterpart by the classical MHC-1 pathway. However, surface-linked antigen is detected very rapidly over Golgi structures and at a relatively high intensity. The mechanism and the significance of this rapid occurrence in the Golgi remain obscure, although it might involve transfer of lipopeptides such as observed with fluorescent lipids or such as promoted by amino-phospholipids (Kagiwada et al. 1993; Rao et al. 1999).
Presentation of Encapsulated and Surface-linked Antigens
The intracellular movements of both Iad and antigens give important clues about the pathways used by activated macrophages to present liposomal antigens. Both liposomal antigens are recovered at the endosomal and plasma membranes in the first hour of chase, excluding the involvement of newly synthesized MHC-II molecules, which take on average 3–4 hr to transit from the peptide loading compartment to the cell surface (Castellino and Germain 1995; Ferrari et al. 1997). This rapid expression of peptides is more consistent with the use of an alternative pathway, independent of protein synthesis, invariant chain, and HLA-DM (Pinet and Long 1998), in which recycling MHC-II molecules reinternalized from the cell surface into early endosomal compartments promote the formation of unstable peptide-MHC complexes (Bénaroch et al. 1995; Warmerdan et al. 1996). With encapsulated antigen, a second wave of membrane surface expression is observed later but the coincidence of this second wave with the increased unregulated expression of Iad directly at the membrane suggests that the alternative pathway is still preferentially operating. The instability of the peptides-MHC complexes thus formed, as well as the capacity of encapsulated antigen to access the MHC-I loading compartment, might contribute to rapidly turning off the influences of macrophages, labeling the cell for CTL action, and impeding the complete maturation of helper functions. This interpretation agrees with the intense but short-lasting antibody response and the pattern of cytokine expression associated with the encapsulated formulation (Thérien et al. 1990; Fortin et al. 1996).
Surface-linked antigen does not access as readily as the encapsulated antigen to the classical MHC-I pathway but uses more thoroughly the various possibilities of the MHC-II pathway. Rapid and late expression of membrane-associated CA is observed and both correlate with high relative expression of membrane Iad. If recycling MHC molecules are more probably involved at the earliest times of the kinetics, with the consequent formation of unstable complexes, newly synthesized molecules entering the endocytic pathway at the CV level, loaded in the more mature GVs and in lysosomes, then returned to CVs for surface expression, also contribute to presentation. This is indicated by the increasing presence of membrane Iad in GVs until 3 hr of chase, the increased detection of membrane CA at 7 hr in CVs, and the more or less constant recovery of surface CA with time. These observations are consistent with the works of Weaver and Voss (1999) regarding the use of multiple loading compartments for MHC-II molecules in macrophages. More important is the observation that the formation of exosome-like structures is initiated in GVs and that these structures, recovered later in CVs, are regurgitated into the extracellular milieu. These overall data suggest that surface-linked antigen is more efficiently processed and presented by macrophages than encapsulated antigen and therefore can contribute more actively to the sensitization of other APCs and to the propagation of activation, in agreement with the long-lasting and more mature response that is induced by this kind of liposomal formulation. This conclusion is further supported by the observed proliferation of D10 G4.1 cells, a Th2 cell line recognizing conalbumin peptide 131–146 in the I-Ak context, in response to macrophages pulsed with encapsulated or surface-linked antigen (data not shown).
Conclusion
This comparative immunocytochemical study of the intracellular circulation of encapsulated and surface-linked liposomal antigens in macrophages contributes to a better understanding of the mechanisms involved in their immunopotentiating properties. Although, as previously demonstrated, both types of antigens can participate in the potentiation of cell-mediated cytotoxicity, distinct mechanisms appear to operate, among which are classical MHC-I presentation and exosome-like secretion. However, only the surface-linked formulation was shown to sustain presentation in the MHC-II context, which is essential for the enhancement of those helper functions necessary to cellular immunity and memory induction. These properties should be exploited to advantage in the development of alternatives to conventional adjuvants that mainly stimulate humoral immunity.
Footnotes
Acknowledgments
Supported by a grant from the Natural Sciences and Engineering Council of Canada.
We sincerely thank Dr John Abrams for his help in revising the manuscript.