
Abstract
Omega class glutathione transferase (GSTO) has been recently described in a number of mammalian species. We used immunohistochemistry to determine the cellular and tissue distribution of GSTO1–1 in humans. Expression of GSTO1–1 was abundant in a wide range of normal tissues, particularly liver, macrophages, glial cells, and endocrine cells. We also found nuclear staining in several types of cells, including glial cells, myoepithelial cells of the breast, neuroendocrine cells of colon, fetal myocytes, hepatocytes, biliary epithelium, ductal epithelium of the pancreas, Hoffbauer cells of the placenta, and follicular and C-cells of the thyroid. These observations and the known activity of GSTO1–1 suggest biological functions that are not shared with other GSTs. (J Histochem Cytochem 49:983–987, 2001)
G
Recently, a new human GST isoenzyme was identified by analysis of the Expressed Sequence Tag (EST) database (Board et al. 2000). This isoenzyme has novel structural and functional characteristics and represents a new GST class termed omega (GSTO), in accordance with the established human GST nomenclature convention (Mannervik et al. 1992). GSTO1-1 is a dimer of identical subunits with a characteristic GST fold composed of an N-terminal glutathionebinding domain and a C-terminal domain composed entirely of α-helices (Board et al. 2000). Unlike other GSTs, GSTO has thiol transferase and dehydroascor-bate reductase activities. In addition, recent studies have shown that GSTO has structural similarities to a family of intracellular chloride channels (CLIC proteins) and is able to modulate cardiac ryanodine receptor calcium release channels (Dulhunty et al. 2001).
To learn more about the role and significance of the omega class GSTs we investigated the expression of GSTO1-1 using immunohistochemistry in a wide range of normal human tissues.
Materials and Methods
Polyclonal antiserum was prepared in rabbits by immunization with recombinant human GSTO1-1 that had been expressed in E. coli and purified by means of an N-terminal His tag (Board et al. 2000). To remove antibodies to E. coli and other nonspecific crossreacting components, the antiserum was absorbed against ultrasonicated E. coli before dilution and use in immunohistochemical studies. The specificity of the antiserum was confirmed on a Western blot of homogenates from human liver and Jurcat cells. The antiserum reacted with a single protein that corresponded in size to purified recombinant GSTO1 (data not shown).
Archival paraffin blocks of surgical pathology specimens or postmortem tissue specimens from the Department of Anatomical Pathology at the Canberra Hospital were used to obtain tissue sections for immunohistochemistry. In all cases the tissue had been fixed in 10% neutral phosphate-buffered formalin before processing and embedding in paraffin. A pathologist (JED) examined the hematoxylin and eosin sections to exclude light microscopic pathology. At least three samples of each site from different subjects were selected.
Distribution and localization of GSTO1-1 in human normal tissues determined by immunohistochemistry and compared with Northern blotting analysis and EST screen reported by Board et al. (2000) a
n.d., not done.
Sections were cut at 3 μm onto slides coated with 3-aminopropyltriethoxy-silane (Sigma; St Louis, MO). Sections were deparaffinized and endogenous peroxidase blocked with 3% hydrogen peroxide in PBS. Nonspecific binding was blocked by incubating sections with 10% normal goat serum in PBS for 20 min at room temperature (RT). Sections were then incubated in rabbit anti-human GSTO1-1 (1:300) in antibody diluent (1% bovine serum albumin, 0.05% sodium azide in PBS) overnight at RT in a moist box. Pre-immune rabbit serum (1:300) and antibody diluent served as negative controls. After 24 hr, sections were washed (three times for 5 min) in PBS and incubated with biotinylated goat antiserum to rabbit IgG (1:1000 in PBS) (Vector Laboratories; Burlingame, CA) for 1 hr at RT. Sections were washed with PBS and streptavidin (Vector Laboratories) labeled with horseradish peroxidase (1:1500 in PBS). Peroxidase activity was developed using 3,3′-diaminobenzidine. Sections were counterstained with hematoxylin, then dehydrated, cleared, and mounted. Sections were viewed by a pathologist and photographed.
(
(
(
(
Results
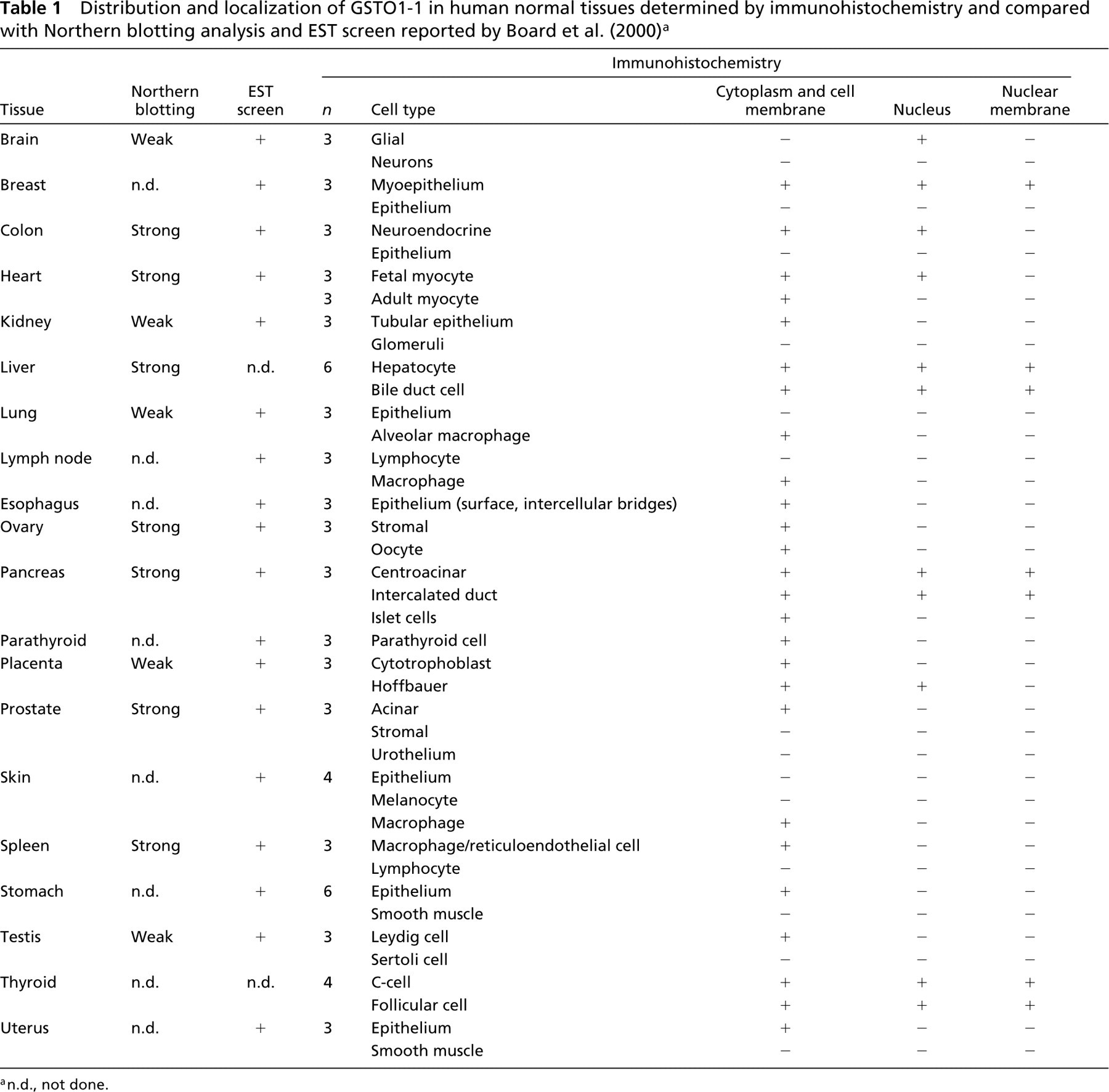
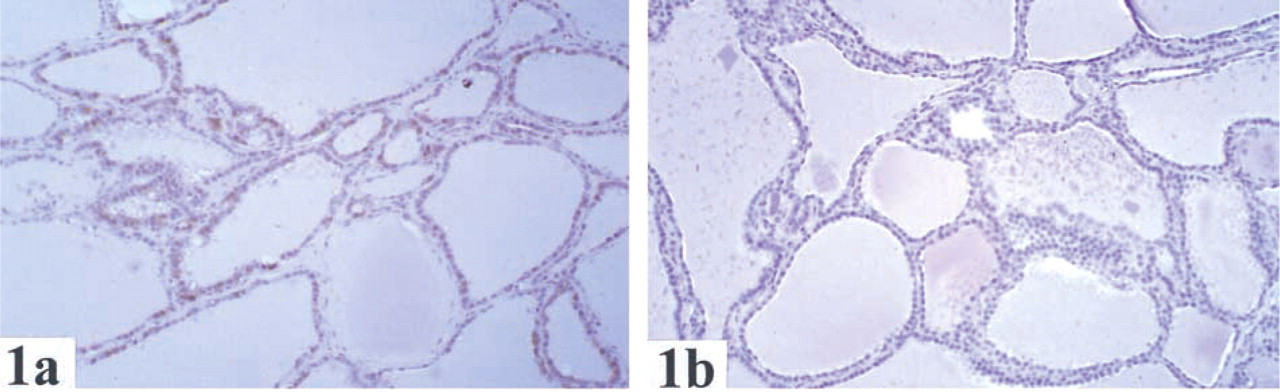
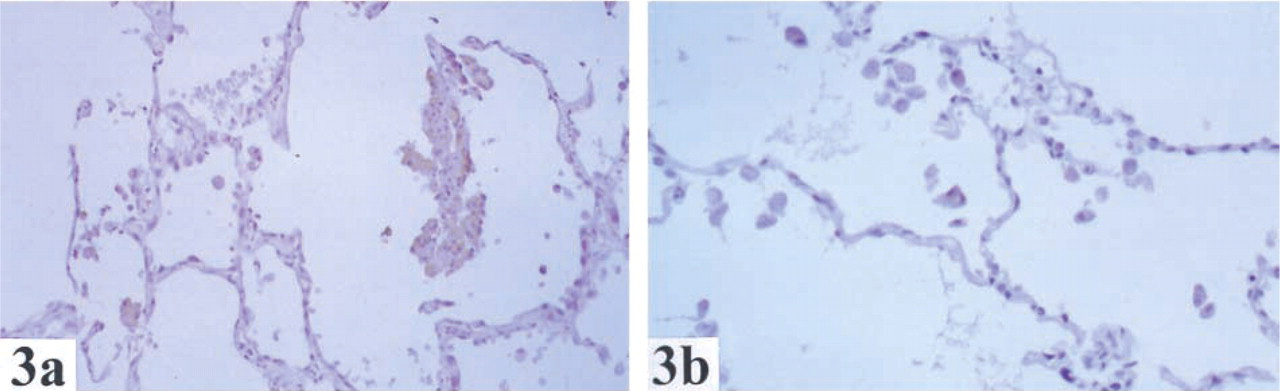
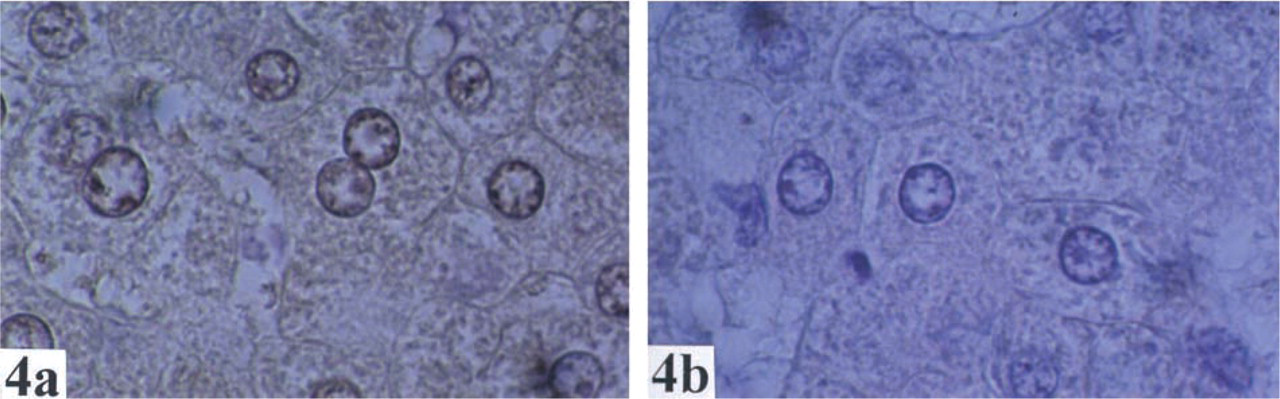
The distribution and localization of GSTO1-1 in human normal tissues are summarized in Table 1. Cytoplasmic staining of GSTO1–1 was observed in most tissues. Of note, there was staining in endocrine cells (e.g., thyroid (Figure 1), pancreas, Leydig cells of testis, neuroendocrine cells of the colon, stromal cells of ovary), glial cells (Figure 2), macrophages (Figure 3), hepatocytes (Figure 4), and epithelial cells in some but not all tissues.
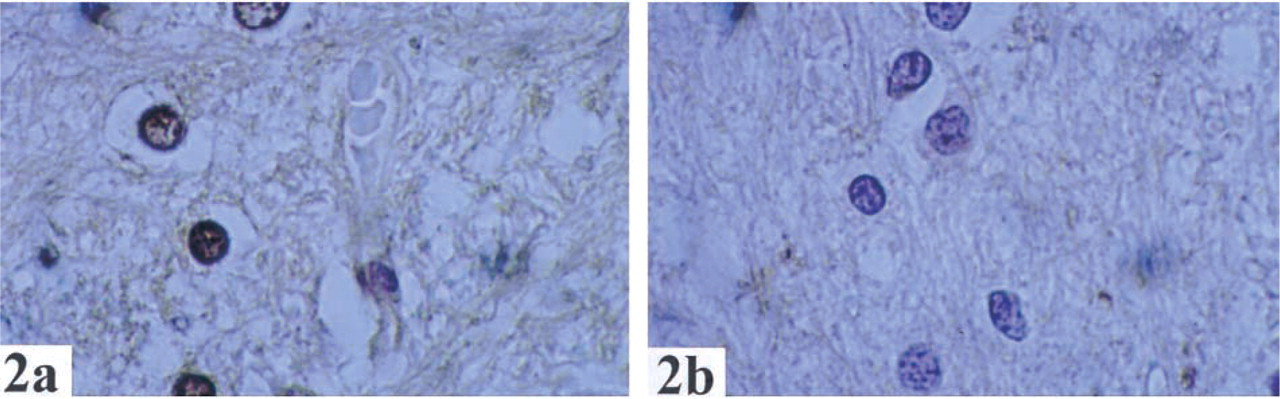
In many tissues, GSTO1–1 was seen in only one cell type (Table 1). For example, GSTO was expressed in neuroendocrine cells in the colon, macrophages in lymph nodes, tubular cells of kidney, and glial cells of the brain. In the skin and lung (Figure 3), staining was seen only in macrophages. GSTO1–1 was not detected in endothelial cells, smooth muscle cells, or lymphocytes in any organ examined.
Nuclear staining was seen in glial cells, myoepithelial cells of the breast, neuroendocrine cells of colon, fetal myocytes, hepatocytes, biliary epithelium, ductal epithelium of the pancreas, Hoffbauer cells of the placenta, and follicular and C-cells of the thyroid (Table 1).
Discussion
Recently, Board et al. (2000) reported the characterization of a new class of GSTs termed omega (GSTO). Northern blotting analysis and screening of EST databases suggested expression of GSTO1–1 in a wide range of human tissues (Table 1). Using immunohistochemical analysis of human tissues we have confirmed widespread expression of GSTO1–1 and in addition have gained information about the cellular location of GSTO1–1 and the cell types in the tissues that express GSTO. Specifically, GSTO1–1 was localized in the nuclei and nuclear membranes of many cell types, including glial cells, myoepithelium of the breast, neuroendocrine cells of colon, fetal myocytes, hepatocytes, biliary epithelium, ductal epithelium of the pancreas, Hoffbauer cells of the placenta, and follicular cells of the thyroid (Table 1). GSTs are usually thought to be cytoplasmic enzymes. However, nuclear localization has been recognized. Nuclear expression of GSTP has been reported in normal epithelium of esophagus (van Lieshout et al. 1999) and cervix (Randall et al. 1990) and in neoplastic lesions of skin (Shimizu et al. 1995), cervix (Randall et al. 1990), esophagus (van Lieshout et al. 1999), mouth (Zhang et al. 1998), and gliomas (Ali-Osman et al. 1997). Nuclear expression of a theta class GST (GSTT) has also been observed in hepatocytes and bile duct cells (Quondamatteo et al. 1998). The biological role of nuclear GSTs presumably relates to protection of DNA from xenobiotic and oxidative injury or breakdown of peroxidized DNA (Ketterer et al. 1987). Although a complete study of the nuclear expression of GSTs is not available, it does appear that GSTO1–1 is unusual in the extent of its expression within the nuclei of many tissues.
The potential nuclear activity of GSTO1-1 is unknown. A mouse GST called p28 with 72% sequence identity with GSTO1 and clearly a member of the omega class has been identified in a radiation-resistant cell line (Kodym et al. 1999). Kodym et al. (1999) reported that p28 moves from its cytosolic location in 3T3 cells to the nucleus in response to heat shock and suggested that p28 is a component of the stress response mechanism. Board et al. (2000) suggested that GSTO1-1 has a significant role in the cellular response to oxidative stress because of its dehydroascorbate reductase and thioltransferase activities. Together, these observations indicate that GSTO1-1 may have an important role as a nuclear antioxidant system. Recent studies have predicted structural similarities between the omega class GSTs and CLIC-1, a soluble protein that forms nuclear chloride channels (Valenzuela et al. 1997; Tulk et al. 2000; Dulhunty et al. 2001). It is not yet clear if GSTO1-1 is also capable of forming channels in the nuclear membrane. The molecular bases for the nuclear membrane interactions exhibited by GSTO1-1 and CLIC-1 are yet to be determined. However, there may be a common mechanism, given the predicted structural similarities.
An additional advantage of immunohistochemical techniques over Northern blotting analysis and EST screening is the ability to determine which cells in each tissue express GSTO. For example, we found specific expression in neuroendocrine cells of the colon, macrophages in lymph nodes, Hoffbauer and cytotrophoblastic cells in the placenta, and glial cells in the brain. In the skin and lung, expression of GSTO1-1 was seen only in macrophages. It is likely that expression of GSTO in lung and skin detected by Northern blotting and EST database screening (Board et al. 2000) reflects expression of this gene in macrophages rather than cells specific to skin and lung. GSTO1-1 was found in epithelial cells in the upper gastrointestinal tract and organs (esophagus, stomach, biliary epithelium, centroacinar and intercalated duct cells of pancreas) but not in the epithelium of the lower gastrointestinal tract. There also appeared to be wide-spread expression in endocrine cells (follicular cells of thyroid, Leydig cells of testis, stromal cells of ovary, neuroendocrine cells of colon).
The widespread distribution of GSTO1-1 suggests that it has important biological functions. It is possible that the glutathione-dependent dehydroascorbate reductase and thiol transferase activities of GSTO1-1 may be generally required for normal cellular processes (Board et al. 2000). Despite its characteristic GST-like fold, GSTO1-1 clearly exhibits a number of atypical structural and functional features (Board et al. 2000; Dulhunty et al. 2001). The pattern of expression of GSTO may reflect the particular requirements of these tissues for specialized xenobiotic metabolism, anti-oxidant activity, or ion channels not provided by other GSTs. Our observations that GSTO1-1 can modulate the calcium release activity of cardiac ryanodine receptors have revealed a novel role for this protein. Because ryanodine receptors are found in the endoplasmic reticulum of many cells, this new role is compatible with the wide distribution of GSTO1-1.
In conclusion, GSTO1-1, like other GSTs, has a distinct pattern of tissue expression. In particular, we noted expression in nuclei of many tissues. These observations, coupled with prior understanding of the unique activity of GSTO1-1, suggest biological functions not shared with other GSTs.
Footnotes
Acknowledgments
Supported in part by the Private Practice Trust Fund of the Canberra Hospital.