
Abstract
Class kappa glutathione S-transferases are a poorly characterized family of detoxication enzymes whose localization has not been defined. In this study we investigated the tissue, cellular, and subcellular distribution of mouse glutathione S-transferase class kappa 1 (mGSTK1) protein using a variety of immunolocalization techniques. Western blotting analysis of mouse tissue homogenates demonstrated that mGSTK1 is expressed at relatively high levels in liver and stomach. Moderate expression was observed in kidney, heart, large intestine, testis, and lung, whereas sparse or essentially no mGSTK1 protein was detected in small intestine, brain, spleen, and skeletal muscle. Immunohistochemical (IHC) analysis for mGSTK1 revealed granular staining of hepatocytes throughout the liver, consistent with organelle staining. IHC analysis of murine kidney localized GSTK1 to the straight portion of the proximal convoluted tubule (pars recta). Staining was consistent with regions rich in mitochondria. Electron microscopy, using indirect immunocolloidal gold staining, clearly showed that mGSTK1 was localized in mitochondria in both mouse liver and kidney. These results are consistent with a role for mGST K1–1 in detoxification, and the confirmation of the intramitochondrial localization of this enzyme implies a unique role for GST class kappa as an antioxidant enzyme.
G
The murine class kappa transferase, mGSTK1–1, was recently identified (Jowsey et al. 2003b). The catalytic properties of this enzyme were investigated using several typical substrates, and although it exhibited substantial GSH-conjugating activity towards 1-chloro-2,4-dinitrobenzene (CDNB) and other aryl halides, significant transferase activity was not demonstrated for other physiologically relevant GST substrates. The biological role of class kappa GST therefore remains uncertain.
The soluble transferases are frequently referred to as cytosolic GSTs, inferring that they are all cytoplasmic. However, the rat class kappa GST was originally isolated from the matrix of liver mitochondria (Harris et al. 1991), and we have previously shown that in murine liver mGSTK1–1 is associated with a subcellular fraction enriched with mitochondria (Jowsey et al. 2003b). However, it is important to note that mitochondrial association is not exclusive to the class kappa GSTs. It has been reported in the mouse that GSTA1–1, GSTA4–4, and GSTM1–1 are present in both mitochondrial and cytosolic fractions (Addya et al. 1994; Raza et al. 2002; Robin et al. 2003), but it remains to be established whether GSTK1 is the only transferase subunit located exclusively in the mitochondria.
Recent structural studies of rGSTK1–1 suggest that class kappa transferases evolved in parallel with the main class α, μ, and π GSTs from a thioredoxin/glutaredoxin progenitor (Ladner et al. 2004). To provide further insight into the biological role and significance of the kappa class GST, we investigated the expression and localization of the mGSTK1–1 in a range of murine tissues using multiple immunolocalization techniques. Where appropriate, expression of GSTK1 was compared with that of cytochrome C oxidase because this protein is an endogenous marker for mitochondria. In addition, comparisons were made between the cellular localization of mGSTK1 and mGSTA3 because we have shown GSTA3 to be associated with mitochondria.
Materials and Methods
Chemicals
Unless stated otherwise, these were all obtained from Sigma-Aldrich (Poole, Dorset, UK).
Animals
Male C57BL/6 mice aged 6–8 weeks were obtained from Harlan (Bicester, Oxfordshire, UK) and were sacrificed by asphyxiation in a rising concentration of CO2.
Antibodies
Polyclonal antiserum was prepared in female New Zealand White rabbits by immunization with recombinant mGSTK1–1 (Jowsey et al. 2003b). Antiserum specificity was confirmed by immunoblotting murine tissue homogenates. The antiserum reacted with a single protein that corresponded in size to purified recombinant mGSTK1 (data not shown). Antisera specific for mGSTA1/2, mGSTA3, rGSTA4, mGSTM1/2, rGSTM5, mGSTP1/2, and rGSTT1 have been previously described (Hayes 1986; Hayes and Mantle 1986; McLellan et al. 1991; McLeod et al. 1997; Hayes and McLellan 1999; Hayes et al. 2000). Manganese superoxide dismutase (Mn-SOD) antibodies were kindly provided by Dr. Lesley I. McLellan (Biomedical Research Centre; University of Dundee, Scotland, UK). The monoclonal mouse anti-human cytochrome oxidase subunit I antiserum was supplied by Molecular Probes Europe BV (Leiden, The Netherlands) (Hanson et al. 2002; Miyazaki et al. 2003). Antiserum specific for rat lactate dehydrogenase (LDH) has been described previously (Ellis et al. 1996). The antibody incubation conditions used for IHC analysis are presented in Table 1.
Summary of immunohistochemical primary antibody conditions
Western Blotting Analysis
Organs from three male C57BL/6 mice were homogenized in ice-cold SET buffer (0.25 M sucrose, 5 mM EDTA, 20 mM Tris base, pH 7.4). Liver mitochondria were isolated as described by Jowsey et al. (2003b). Tissue samples (10–15 μg protein/well) were separated by SDS-PAGE (12% polyacrylamide resolving gel) and transferred to a PVDF membrane. Membranes were blocked with 10% skimmed milk, incubated with primary antibody, and the localization of the immobilized GST proteins was detected with HRP conjugates of goat anti-rabbit IgG or rabbit anti-sheep IgG antibody (Bio-Rad; Hercules, CA) and visualized by enhanced chemi-luminescence (Amersham Biosciences; Piscataway, NJ). All primary antibodies were diluted 1:3000 except for mGSTK1 antiserum, which was diluted 1:1000.
Tissue Preparation
For light microscopic analysis, small tissue sections (10 × 10 × 3 mm) were fixed in 10% (v/v) neutral buffered formalin at room temperature (20C) for a minimum of 24 hr and embedded in paraffin. Serial 3-μm sections were cut for each tissue and mounted on Superfrost Plus slides (Menzel-Gläser; Braunschweig, Germany). For electron microscopic analysis, small tissue sections (3 × 3 × 3 mm) were fixed in Karnovsky's primary fixative (1% (w/v) paraformaldehyde, 3% (v/v) glutaraldehyde in 70 mM sodium cacodylate buffer, pH 7.3) for a minimum of 24 hr at 4C and embedded in LR White acrylic resin. Ultrathin sections were cut to a thickness of 70–90 nm and transferred to nickel grids.
Light Microscopy and IHC Staining
Slides were stained using the DAKO EnVision + System (Ely; Cambridge, UK), which uses 3,3′-diaminobenzidine (DAB) as chromogen (Sabattini et al. 1998). Briefly, paraffin-embedded tissue sections were mounted on slides, deparaffinized, and rehydrated by sequential placement in aqueous solutions containing increasing amounts of ethanol (70–95%). Antigen retrieval was achieved by microwave heating for 5 min on full power followed by 5 min on simmer in a citrate buffer or by proteolytic digestion using DAKO Pronase solution (DAKO; Glostrup, Denmark). Endogenous peroxide activity was quenched with an aqueous solution of 3% (v/v) hydrogen peroxide and nonspecific antibody binding was blocked by incubating sections with DAKO Protein Block Serum-Free solution or by incubation in 100 μl of a 5% (w/v) BSA solution in 20 mM Tris-buffered saline, pH 7.4, containing 0.2% (v/v) Tween (TBST) for 20 min at 20C. Sections were incubated in appropriate dilutions of primary antibody for 1 hr at 20C. Dilutions and wash steps were carried out using TBST. Incubation in preimmune serum or incubation in the absence of primary antibody served as negative controls. After primary antibody incubation, sections were washed in TBST for 5 min at 20C and incubated with DAKO EnVision+ System-labeled polymer (peroxidase-labeled polymer conjugated to goat anti-rabbit or goat anti-mouse immunoglobulins) for 30 min at 20C. Sections were washed in TBST for 5 min at 20C. Peroxidase activity was developed with a buffered DAB chromogen solution. Tissues were counterstained in Carazzi's hematoxylin before being dehydrated, cleared, and mounted.
Electron Microscopy and Immunogold Labeling
Immunostaining of ultrathin sections for electron microscopy was carried out using an indirect immunocolloidal gold detection method. Briefly, the sections were equilibrated with TBST for 5 min at 20C. Then the sections were incubated with rabbit anti-mouse GSTK1–1 primary antibody (1:75) diluted with TBST for 1 hr at 20C. Negative control sections were incubated in TBST for 1 hr at 20C. Sections were incubated for 1 hr at 20C in an anti-rabbit IgG immunogold conjugate diluted 1:10 using TBST (BB International; Cardiff, UK). Counterstaining was carried out for 1 min in a 1% (w/v) uranyl acetate solution and in a 50% (v/v) lead citrate solution for 30 sec. Sections were observed in the Hitachi H7100 transmission electron microscope.
Results
Tissue-specific Expression of the mGSTK1 Subunit
The expression of GSTK1 protein in murine tissues has not been reported previously and was investigated to identify potential endogenous functions of this transferase. Immunoblotting analysis of total homogenates from mouse tissues revealed that GSTK1 protein expression was highest in the liver and stomach. Moderate expression was observed in the kidney, heart, large intestine, testis, and lung. By contrast, little or no GSTK1 was detected in small intestine, brain, spleen, and skeletal muscle (Figure 1). Use of LDH as a loading control confirmed that the GSTK1 expression profile reflects genuine differences in the tissue levels of this protein.
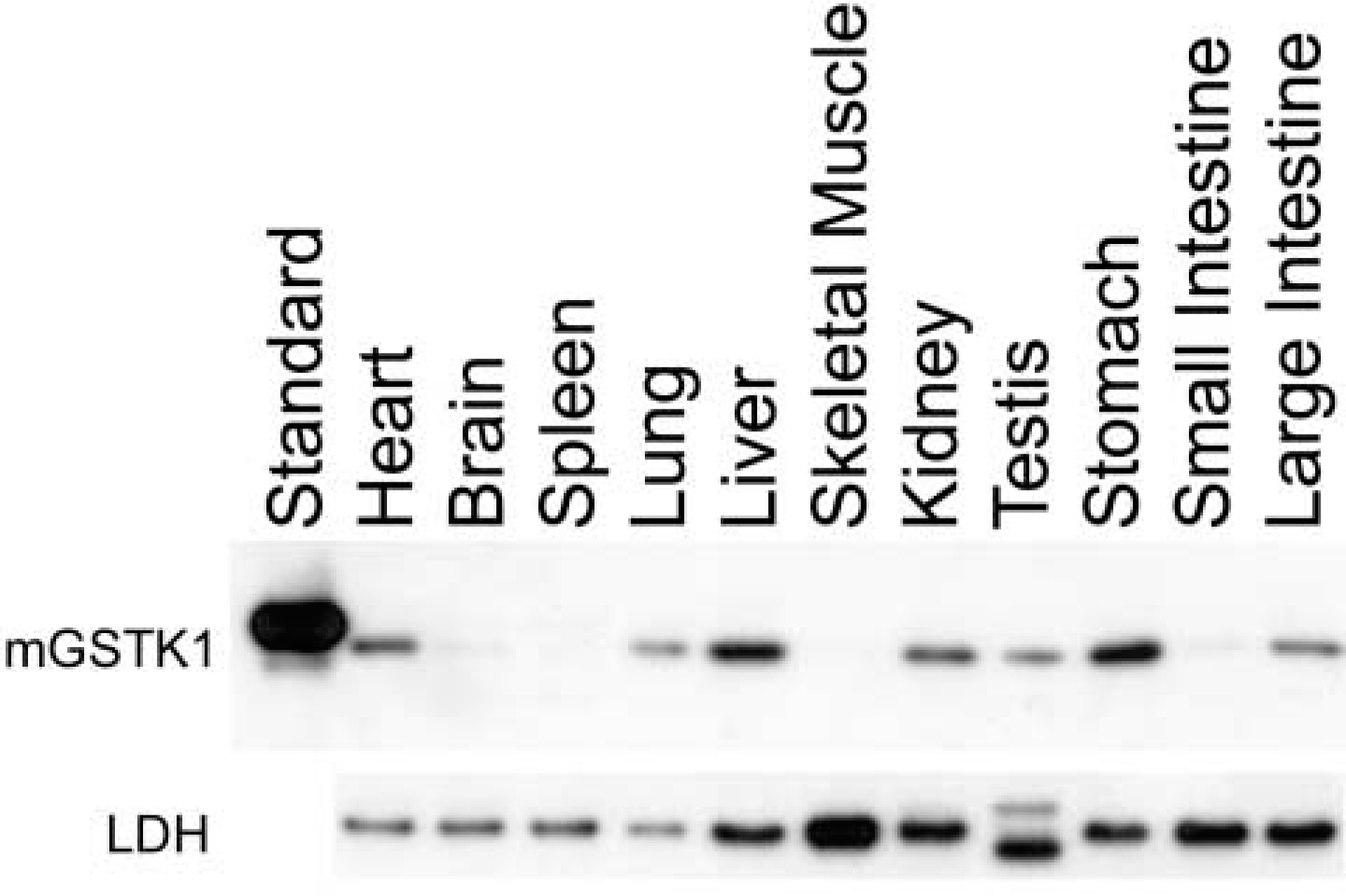
Western blotting analysis of mGSTK1 and LDH in murine tissues. Immunoblots of 10-μg total protein of whole mouse tissue homogenates were incubated with polyclonal anti-mGSTK1 antiserum or polyclonal anti-rLDH.
Cellular Localization of mGSTK1 in Liver and Kidney
IHC analysis was performed on liver and kidney sections from five male C57BL/6 mice using polyclonal anti-mGSTK1 serum or preimmune serum (Figure 2). IHC analysis of murine liver samples revealed intense hepatocyte staining throughout the entire organ (Figure 2A) and the granular appearance was consistent with specific staining of subcellular organelles (Figure 2B). Murine GSTK1 protein was not detected in nuclei or biliary epithelium, and only minimal staining was observed in control sections incubated with preimmune serum (Figure 2C).
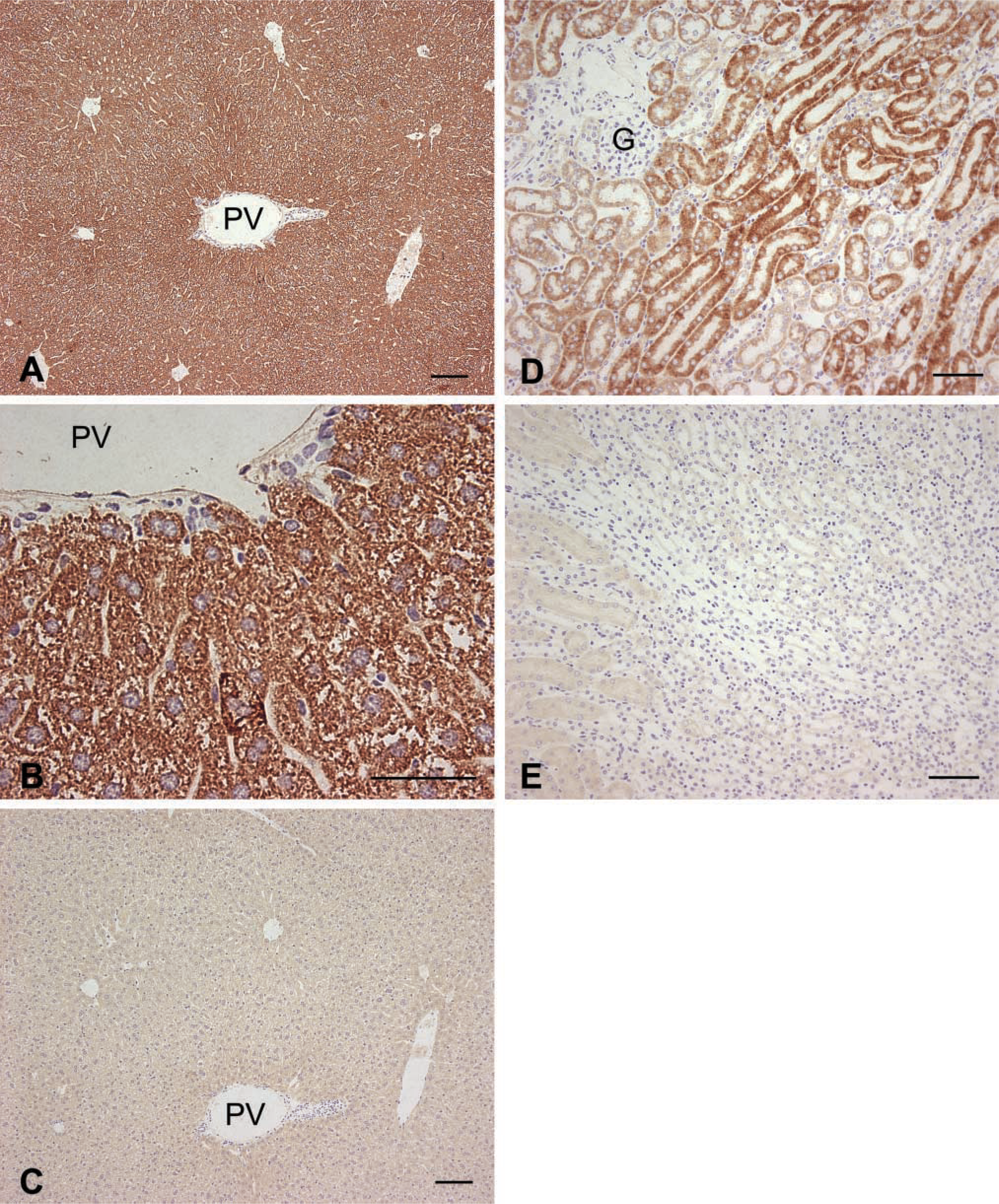
IHC detection of mGSTK1 protein in murine liver and kidney. Sections were incubated with polyclonal anti-mGSTK1 antibodies (
IHC analysis of kidney sections (Figure 2D) revealed intense mGSTK1 staining in the straight portion of the proximal convoluted tubule (pars recta). This region of the proximal convoluted tubule (PCT) is responsible for highly selective active ion re-absorption, and therefore is rich in mitochondria to provide ATP for these activities. Staining was moderate in the pars convoluta but by comparison was only slight in the distal convoluted tubule (DCT). Murine GSTK1 protein was not present in the loop of Henle, collecting ducts, or glomeruli. Control sections incubated with preimmune serum were devoid of staining (Figure 2E).
Distribution of GSTK1 in Mouse Kidney Is Distinct from That of Cytochrome C Oxidase or GSTA3
As described previously, GSTK1 expression was most intense in the pars recta (Figures 3A and 3B). Featherlike staining of GSTK1 in this region was observed at high magnification (Figure 3C). The glomeruli, loop of Henle, DCT, and collecting ducts showed no evidence of GSTK1 expression by IHC examination. The mitochondrial protein cytochrome C oxidase was detected throughout the kidney, with the most intense staining observed in the pars recta (Figure 3D). Expression of this oxidase was also observed in the loops of Henle, DCT, and pars convoluta. Although the collecting ducts in the papilla showed only moderate cytochrome C oxidase expression, staining of mitochondria in this region was very granular in appearance (Figure 3E). A feather-like staining was seen in the convoluted tubules at high magnification (Figure 3F), and the glomeruli lacked detectable expression of the oxidase.
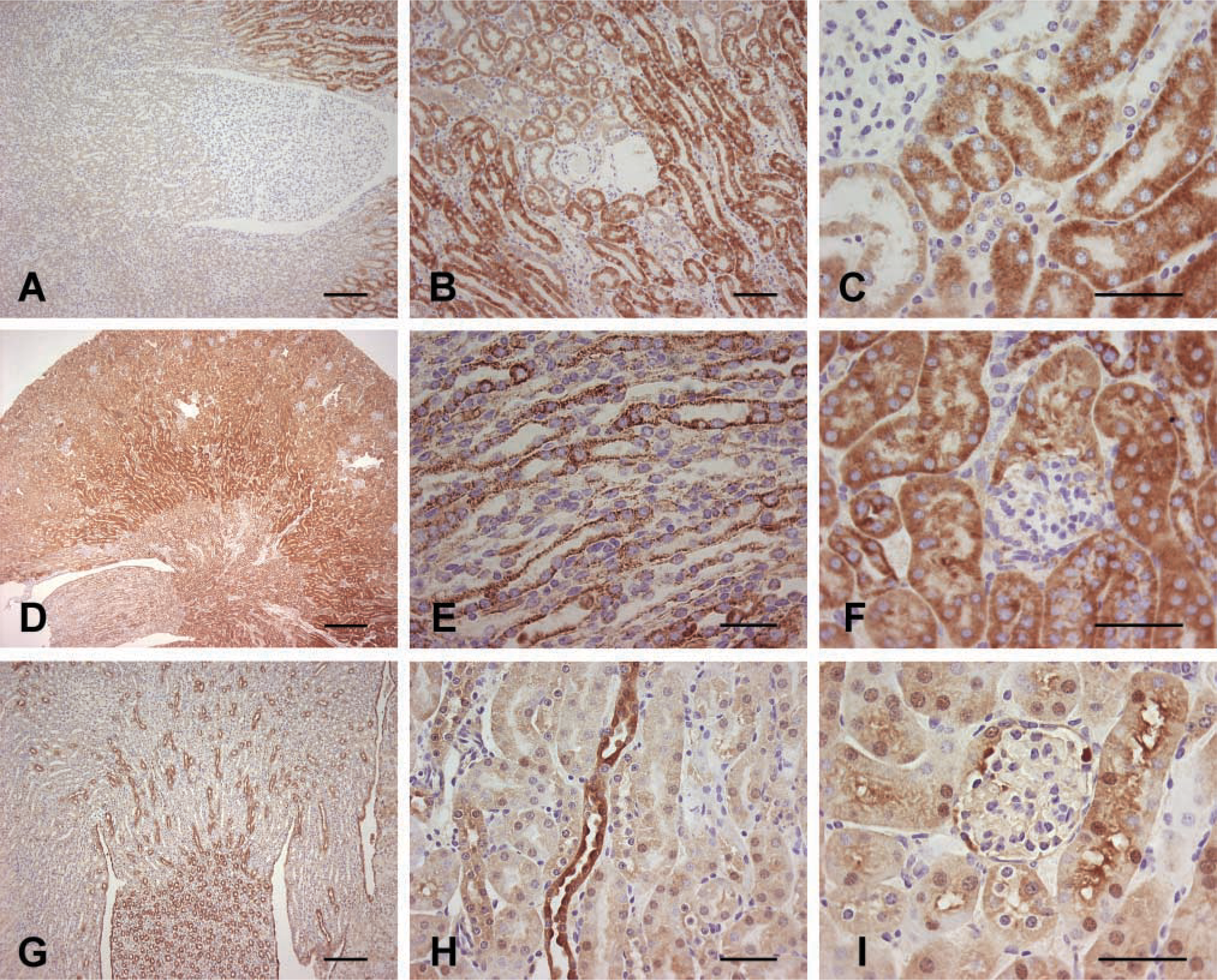
IHC detection of GSTK1, cytochrome C oxidase, and GSTA3 in murine kidney. Sections were incubated with polyclonal anti-mGSTK1 (
GSTA3 was observed predominantly in the collecting ducts of the papilla (Figure 3G). However, in contrast to cytochrome C oxidase, GSTA3 staining was not granular in appearance. Collecting tubules in the renal medulla and cortex were also stained (Figure 3H). By contrast with GSTK1, expression of GSTA3 was weak in the pars recta and only moderate in the pars convoluta and glomeruli (Figure 3I). Staining of the pars convoluta was especially intense in the region of the brush border.
Electron Microscopy Reveals That mGSTK1 Is Located in Mitochondria of Liver and Kidney
Subcellular localization of mGSTK1 protein in liver and kidney was investigated using an immunogold-labeled polyclonal antibody. In both hepatic and renal sections, strong staining for mGSTK1 was observed in mitochondria (Figures 4A and 4B). In both organs, staining was predominantly located within the matrix region of mitochondria and almost no staining was associated with the outer mitochondrial membrane or the intermembrane space. Hepatic and renal mitochondria have densely packed cristae and it is therefore difficult to determine whether mGSTK1 is associated with the inner mitochondrial membrane or the matrix. By making comparisons with control sections immunostained in the absence of primary antibody, the gold particles observed outside mitochondria were regarded as due to nonspecific binding (data not shown).
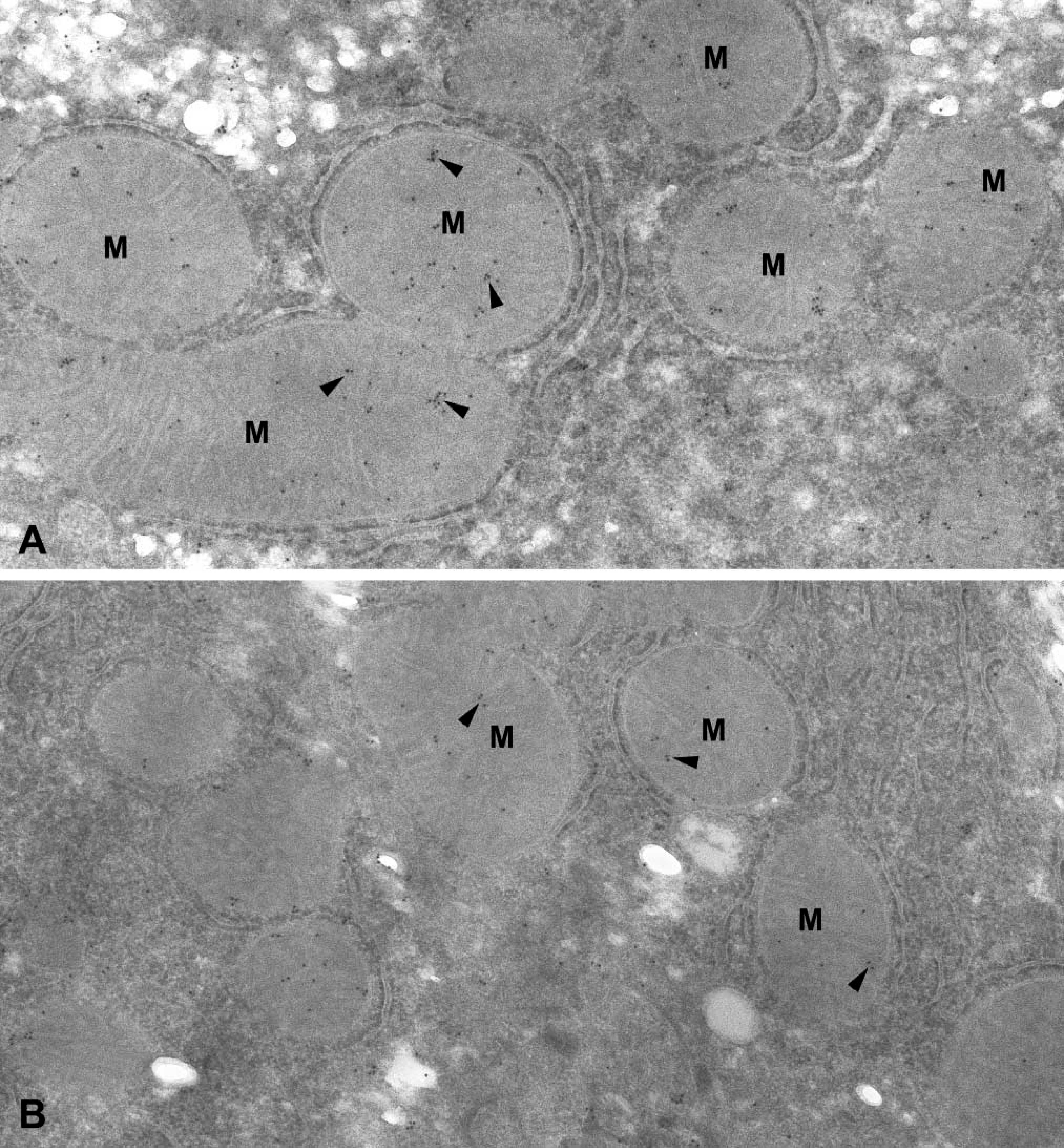
Immunogold-labeled polyclonal anti-mGSTK1 antibody of murine liver and kidney. Arrows denote the localization of murine GSTK1 to mitochondria (M) in both liver (
Class Kappa Transferase Exhibits a Unique Subcellular Distribution Among GSTs
In view of reports that class alpha and class mu transferases can associate with mitochondria (Addya et al. 1994; Raza et al. 2002; Robin et al. 2003), experiments were performed to determine whether mGSTK1 is unique in its exclusive location in mitochondria (Figure 5). Western blotting analysis of murine hepatic subcellular fractions showed that no GSTK1 protein was recovered in the cytosolic fraction (Figure 5d). By contrast, approximately similar levels of GSTA3 were found in both the mitochondrial and cytosolic fractions (Figure 5b). GSTA1/2 and GSTM1/2 were also detected in both mitochondrial and cytosolic fractions (Figures 5a and 5e, respectively), although in both cases the transferase subunits were not abundant in the mitochondrial fraction. By contrast with GSTA1/2 and GSTM1/2, GSTA4, GSTM5, GSTP1/2 and GSTT1 were essentially recovered only in the cytosolic fraction (Figures 5c and 5e–5h). Western blot analysis of fractions using antibodies specific for predominantly mitochondrial (MnSOD) and cytosolic (LDH) proteins confirmed the purity of these fractions (Figures 5i and 5j).
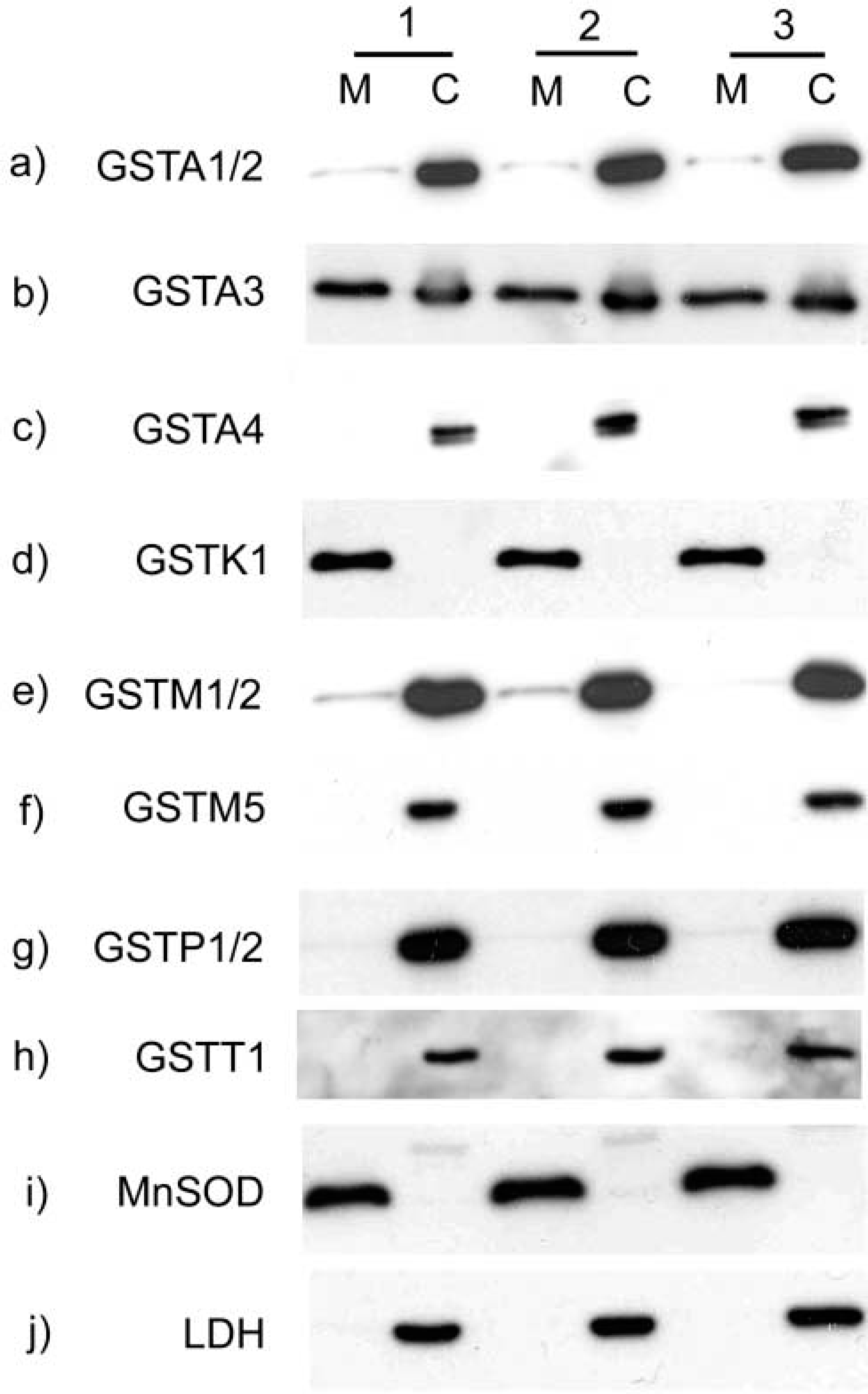
Subcellular localization of GST isoenzymes. Mitochondrial (M) and cytosolic (C) fractions were prepared by centrifugation from livers isolated from three mice (individual preparations numbered 1–3). Protein samples were resolved by SDS-PAGE and analyzed by Western blotting using antisera specific for GSTA1/2 (
In accordance with the reports of Raza et al. (2002), we showed that GSTA1/2 and GSTM1/2 were present in both mitochondrial and cytosolic murine liver fractions (Figures 5a and 5e, respectively). However, in contrast to the findings of Raza et al. (2002) and Robin et al. (2003), we did not observe an association between GSTA4 and the mitochondrial fraction (Figure 5c). This discrepancy might be explained by the fact that the mitochondrial protein concentration resolved by SDS-PAGE in this study was only 10 μg compared with at least 80 μg in the studies described by Raza et al. (2002) and Robin et al. (2003).
Discussion
Among the eight mammalian families of soluble GST, class kappa enzymes are the least well characterized. Essentially nothing is known about the physiological properties and regulation of GSTK1. Because class kappa enzymes are only distantly related to the other soluble GST families (Sherratt and Hayes 2002; Ladner et al. 2004), it is likely that this class performs unique biological functions. The present study was therefore undertaken to allow predictions of the physiological role of class kappa to be made on the basis of its tissue-specific expression and its subcellular distribution in the mouse. Furthermore, these properties also allow assessment of whether class kappa GST performs roles that are distinct from those of other soluble transferases.
Tissue-specific Expression of mGSTK1
Western blotting clearly shows that, in the mouse, GSTK1 expression is most intense in liver and stomach, organs that are usually associated with detoxification of harmful endogenous and exogenous compounds. GSTK1 is also expressed at moderate levels in kidney, heart, large intestine, testis, and lung. Several other murine GSTs show a similar distribution profile to mGSTK1. Class theta GSTs, for example, are present in mouse liver and lung (Mainwaring et al. 1996). In the mouse, GSTA3, GSTP1, and GSTM1 show high expression levels in the liver, lung, kidney, and testis (Mitchell et al. 1997; Jowsey et al. 2003a) and these three isoenzymes have been associated with the detoxification of carcinogens/mutagens, environmental pollutants and anticancer drugs (Hayes and Pulford 1995). These class alpha, mu, and pi GSTs have been shown to be inducible in gastrointestinal tissues by cancer chemopreventive agents, suggesting their involvement in protection against carcinogenesis (McLellan et al. 1992; McMahon et al. 2001; Andorfer et al. 2004). Because mGSTK1 is expressed at relatively high levels in liver and at moderate levels in kidney, its cellular distribution within these organs was investigated to gain a better idea of the cell types in which the transferase may confer protection against cytotoxic compounds.
Expression of GSTK1 in Murine Liver Is Restricted to Hepatocytes
IHC shows that mGSTK1 is expressed in hepatocytes throughout the liver. Staining for mGSTK1 is not observed in bile canaliculi or sinusoid lining cells, and there appears to be no significant difference in periportal or centrilobular staining. The granular pattern of expression is consistent with organelle staining, and this observation is in agreement with our previous finding that GSTK1 is associated with the mitochondrial subcellular fraction prepared from murine liver (Jowsey et al. 2003b). Hepatocytes contain many mitochondria, which may explain the high expression of mGSTK1 in these cells. However, because alpha, mu, omega, theta, and zeta GSTs also localize to hepatocytes in the liver (Lantum et al. 2002), it is possible that expression of hepatic mGSTK1 in liver is not simply due to the presence of mitochondria but is related to a detoxification function that is common to these classes of transferase.
GSTK1 Displays a Distinct Localization in Murine Kidney
The distribution of mGSTK1 in the kidney was specific to the pars recta portion of the PCT. This region of the PCT is rich in mitochondria, thereby providing ATP for the active and selective reabsorption of essential ions. To help establish whether GST class kappa is localized to the pars recta merely as a consequence of its association with mitochondria or because it possesses a unique function, comparisons were made between the renal distribution of mGSTK1 and the mitochondrial protein cytochrome C oxidase. We postulated that if mGSTK1 were localized to the pars recta solely because of its mitochondrial association, cytochrome C oxidase would be detected in exactly the same region. Cytochrome C oxidase expression is observed throughout the kidney (Figure 3D), with the most intense staining in the pars recta. Magnification of this region emphasizes similarities between the feather-like staining of mGSTK1 (Figure 3C) and cytochrome C oxidase (Figure 3F). This specific staining pattern might be caused by the alignment of mitochondria perpendicular to the basal epithelium of the tubule. The similarities between the localizations of mGSTK1 and cytochrome C oxidase clearly support the mitochondrial association of mGSTK1. However, despite this similarity GSTK1 expression is not identical to that of cytochrome C oxidase. Class kappa GST is specifically localized to the pars recta and is not present in other regions of the kidney that express cytochrome C oxidase. The specific mGSTK1 staining of the pars recta cannot be explained simply by the presence of mitochondria in this region. Therefore, the enrichment of mGSTK1 in the pars recta may be necessary because the transferase exercises an important function required for selective ion reabsorption. Moreover, in contrast to the ubiquitous renal expression of the mitochondrial protein cytochrome C oxidase, the fact that mGSTK1 expression is localized predominantly to the pars recta strongly suggests that mGSTK1 expression is specifically regulated.
Because of the observation that GSTA3 is localized to both the mitochondrial and the cytosolic fraction of murine liver (Figure 5b), comparisons were made by IHC analysis between the localizations of GSTK1 and GSTA3 in murine kidney. By contrast with GSTK1, the highest levels of expression of mGSTA3 in kidney were detected in the collecting ducts (Figures 3G and 3H). With antibodies against GSTA3, there is staining of the glomeruli and the brush border of the pars convoluta but no significant staining of the pars recta (Figure 3I). These data show that, despite the apparent shared association of GST K1–1 and GST A3–3 with mitochondria, the localizations of these two isoenzymes differ markedly.
Subcellular Localization of mGSTK1 in Murine Liver and Kidney
The first description of a class kappa GST was a report of purification and N-terminal sequencing of GST13–13 from rat liver mitochondria (Harris et al. 1991). We have subsequently shown an association between mGSTK1 and the mitochondrial subcellular fraction isolated from mouse liver (Jowsey et al. 2003b). The precise localization of the mGSTK1 in this organelle and its mitochondrial association in other organs have not been investigated previously. In the present study, the subcellular localization of mGSTK1 in the mitochondria of hepatic and renal tissue has been demonstrated by electron microscopy using an immunogold-labeled polyclonal antibody (Figures 4A and 4B). We have clearly shown that GSTK1 is not simply associated with outer or inner mitochondrial membranes but rather is present in the matrices of these organelles. Mitochondria are responsible for generating ATP via oxidative phosphorylation through an electron transport chain. A consequence of this process is the generation of reactive oxygen species (ROS) such as superoxide anion radicals (Cadenas and Davies 2000). The balance between production and removal of superoxide anion radicals is maintained by antioxidant enzymes such as superoxide dismutase (Samavati et al. 2002). Detection of mGSTK1 in the mitochondria of hepatic and renal tissue suggests that this class kappa transferase is also highly likely to play an antioxidant role in this organelle.
The present study describes the tissue, cellular, and subcellular localization of murine glutathione S-transferase class kappa. Similarities between the tissue distribution profile of mGSTK1 protein and known detoxification GSTs imply that mGST K1–1 might play a role in xenobiotic detoxification. The confirmation that exclusively mitochondrial localization is unique to the class kappa GST suggests that this isoenzyme may have a novel antioxidant role in protection of mitochondria from the damaging effects of ROS.
Footnotes
Acknowledgements
Supported by a Biotechnology and Biological Sciences Research Council (BBSRC)/AstraZeneca collaborative PhD studentship (RT).
We thank Mr Paul Brislane for preparation of the paraffin-embedded material used in this study.