
Abstract
The aim of this study was to investigate whether transformation of quiescent vascular smooth muscle cells (VSMCs) into proliferating secretory cells is accompanied by an expression of processing enzymes that activate de novo-synthesized growth factors. Three enzymes belonging to the family of the kexin/subtilisin-like mammalian proprotein convertases (PCs), furin, PC5, and PC7, were found to be upregulated after balloon denudation in vivo. To determine their importance in these cell processes, we investigated their gene regulation using a short-term organ culture system. After incubation of rat aorta for 4 and 24 hr in serum-free medium, we demonstrated a significant induction of VSMC proliferation. The affected subset of VSMCs, positive for α-smooth muscle actin, also expressed proliferating cell nuclear antigen (PCNA). Our results revealed a parallel upregulation of furin, PC5, and PC7 in PCNA-immunolabeled cells. As a substrate model for comparison with PCs we used nerve growth factor (NGF). NGF is known to be activated by PCs. As shown by Northern blotting analysis, NGF mRNA concentration was significantly increased in cultured explants. NGF was released into the culture medium. In conclusion, both PCs and NGF are coordinately modulated on induction of VSMC proliferation.
Keywords
P
Each of these enzymes, either alone or in combination with others, is responsible for the tissue-specific processing of multiple polypeptide precursors. This combinatorial mechanism generates a large diversity of bioactive molecules in an exquisitely regulated manner. This type of cleavage occurs in most growth factors and their receptors, polypeptide hormone precursors, surface glycoproteins, including adhesion molecules and viral envelope glycoproteins, and a host of other secretory proteins (Seidah et al. 1998).
The role of these proprotein convertases in vascular tissue remains largely unknown. We hypothesized that PCs might represent important intermediates in the biosynthesis and activation of growth factors involved in vascular remodeling.
In this study we tested the hypothesis that the induction of VSMC proliferation in vivo and in vitro involves, in addition to the de novo synthesis of growth factors, the expression of their processing enzymes as well. To test our hypothesis we used in vivo and in vitro models. After balloon denudation in rodents in vivo, we examined the regulation of PCs. In vitro we investigated the expression of PCs in VSMCs during early stages of VSMC proliferation using an organ culture system. This in vitro model offers the advantage of maintaining cell/cell and cell/matrix interactions, thus preserving an in vivo-like environment.
In this article we report that the rapid onset of VSMC proliferation in vitro triggers the enzymatic machinery including the upregulation of proprotein convertases and the de novo synthesis of growth factors.
Materials and Methods
Preparation of Arterial Explants
Adult male Sprague–Dawley rats weighting 300–350 g (Charles River; St Constant, QC, Canada) were used in this study. Rats were housed in an air-conditioned animal facility (12:12 hours light:dark cycle), fed a standard diet, and received water ad libidum. Rats were sacrificed by IP injection of 50 mg/kg sodium pentobarbital. The aorta was cannulated at the iliac bifurcation, opened at the region of the aortic arch, and blood carefully washed out with Ringer's lactate solution. Each artery was then opened longitudinally, cut into six pieces (about 1 × 1 cm) and transferred into Petri dishes containing PBS. Excessive fat and connective tissue were removed and explants washed three times in PBS. Single pieces of rat aorta were then transferred to six-well dishes (Becton–Dickinson; Franklin Lakes, NJ) and kept under 95% relative humidity and 5% CO2 at 37C in Dulbecco's modified Eagles medium (DMEM, catalogue #12100; Life Technologies, Burlington, ON, Canada), supplemented with gentamycin 26 μg/ml, with the endothelial side upwards.
After culturing, explants were either snap-frozen on dry ice at −80C for immunoblotting and RNA extraction or fixed with 4% formaldehyde in 0.1 M phosphate buffer (pH 7.2) overnight at 4C, then washed in 70% ethanol for 24 hr, embedded in paraffin, and cut into 3-μm slices.
In Vivo Balloon Denudation of Rat Aorta
Balloon injury of rat aorta was performed according to Schwartz et al. (1975). Briefly, animals were anesthetized with ketamine (60 mg/kg IM) and xylazine (6.6 mg/kg IM). Each animal received 100 IU heparin IV before surgery. The left common carotid was exposed by blunt dissection via a midline neck incision. The proximal artery was ligated and a 2F Fogarty catheter (Baxter Healthcare; Irvine, CA) was introduced. The catheter was placed in the distal aorta approximately above the bifurcation, inflated to create a distending resistance, and pulled back to the carotid. The procedure was repeated five times. Sham-operated animals were subject to the same procedure without balloon inflation. After removal of the catheter, the distal carotid was ligated and wounds closed with surgical clips.
Rats were sacrificed in groups of four to six at Day 2, 7, or 28 after the operation. The aorta was fixed in 4% formaldehyde in 0.1 M phosphate buffer (pH 7.2) for 24 hr, washed for 24 hr in 70% ethanol, and then embedded in paraffin.
Immunocytochemistry
The antibodies used are indicated in Table 1, providing information about the antigen, host, protein sequence, and dilution for both Western blotting and immunocytochemistry.
Immunocytochemical stainings were performed with the Zymed Histostain-Plus kit (Zymed Laboratories; San Francisco, CA) as recommended by the manufacturer. The immunoreactions were revealed with horseradish peroxidase activity producing brown staining and/or alkaline phosphatase activity producing blue and/or red staining. When necessary, double immunocytochemical stainings were performed by combining alkaline phosphatase and horseradish peroxidase activity as described (Marcinkiewicz et al. 1999). Blocking immunoreactivity was performed by preincubation of antibodies with an excess of antigen (10–6 M) in the case of furin, PC5, and PC7, or by omission of the first antibody in the case of PCNA and α-SMA. Statistical analysis was by Student's t-test.
Western Blotting Analysis
Immunoblotting was performed as described previously (Marcinkiewicz et al. 1999). Briefly, proteins were extracted in a buffer (Tris-HCl 50 mM, pH 7.4, EDTA 2.5 mM, and NaCl 150 mM) containing freshly dissolved protease inhibitors (leupeptin 2 μg/ml, aprotinin 2 μg/ml, pepstatin 100 μM, A-PMSF 100 μg/ml) and β-mercaptoethanol 2 mM. Up to 50 μg protein was mixed with sample buffer and applied on 8% or 12% Tris-Tricine SDS PAGE. Serum-free media were also used after 10-fold concentration by centrifugation through a #3 Centricon concentrator (Amicon; Beverly, MA). After migration, proteins were transferred onto PVDF membranes (Schleicher & Schuell; Keene, NH). The membranes were first incubated overnight at 4C with the primary antibody, followed by incubation with donkey anti-rabbit IgG–peroxidase (Jackson Laboratories; West Grove, PA) diluted 1:10,000. Peroxidase activity product was revealed with the BM Chemiluminescence blotting substrate POD (Boehringer–Mannheim; Mannheim, Germany). For controls, antisera were preadsorbed with synthetic peptides. Densitometry was done with the Northern eclipse program.
Northern Blotting Analysis
Northern blotting analysis was conducted as described elsewhere (Marcinkiewicz et al. 1999). Briefly, RNA was extracted with TRIzol reagent (Life Technologies; Gaithersburg, MD). Approximately 10 μg of total RNA and RNA ladder (GIBCO/BRL; Burlington, ON, Canada), was separated by electrophoresis on a horizontal gel apparatus in 1.2% agarose gel containing 20 mM HEPES (pH 7.8), 1 mM EDTA, and 6% formaldehyde and then transferred to a nylon membrane of 0.45-μm pore (Nytran Plus; Schleicher & Schuell) by capillary action. The filters were prehybridized at 65C for 90 min in a solution composed of 5% SDS, 0.4 M Na3PO4, 1 mM EDTA, 0.1% bovine serum albumin (BSA), and 50% formamide (GIBCO/BRL), then hybridized overnight at 65C in the presence of [32P]-UTP cRNA probes. These probes corresponded with cDNAs of rat (r)furin823–2053 bp, rPC51089–1925 bp, rPC72170–2911 bp, and mouse (m)NGF388–971 bp. After hybridization, the filters were washed in sodium citrate (SSC) buffer (NaCl 1.5 M, sodium citrate 0.15 M, 0.1 × SSC, 0.1% SDS, and 1 mM EDTA) at 70C for 2 hr, and exposed at −80C for 1–10 days to X-ray film (Kodak; Rochester, NY) with intensifying screen.
Results
Immunocytochemical Localization of PCs after Balloon Denudation In Vivo
Immunocytochemical localization of the proprotein convertases furin, PC5, and PC7 was studied in intact rat aorta and in rat aorta after balloon injury at various time points (Figure 1). In intact rat aorta, PC7 (Figure 1A), furin (Figure 1E), and PC5 (Figure 1G) immunoreactivity was found in rare cells throughout the media. Depicted as an example, immunocytochemical analysis revealed upregulation of PC7 on Day 2 in the media (Figure 1B) and on Day 7 in the beginning neointima (Figure 1C). After establishment of a mature neointima on Day 28, PC7 was found throughout the neointima (Figure 1D). Furin (Figures 1E and 1F) and PC5 (Figures 1G and 1H) paralleled the time course of PC7 upregulation (Days 2 and 7 not depicted). On the basis of staining of consecutive sections with α-SMA, medial and neointimal cells expressing PC immunoreactivity were identified as VSMCs (Figures 1I and 1J). Studied for comparison, the proprotein convertases PC1 and PC2 were not detected in VSMCs (data not shown).
As noted by Zeymer et al. (1992) and Wei et al. (1997), low levels of PCNA immunoreactivity were found in intact rat aorta. After balloon injury, strong upregulation of PCNA was noted on Day 2 in the media and after the appearance of a neointima on Day 7 therein. PCNA strongly decreased on Day 28, with only a few positive cells on the luminal surface of the neointima (data not shown). Therefore, neointimal cells expressing furin, PC5, and PC7 immunoreactivity outnumbered PCNA-positive cells after the establishment of a mature neointima. To define the relationship between PCs and VSMC proliferation more closely, we investigated their regulation using an in vitro organ culture system of rat aorta.
Western Blotting Analysis of Cell Proliferation in Explant of Rat Aorta: Evidence for Upregulation of PCs
To detect cell proliferation in rat aorta cultured for 24 hr in serum-free medium, we performed Western blotting analysis with proliferating cell nuclear antigen (PCNA) using an appropriate mouse monoclonal antibody (Table 1). PCNA is associated with DNA synthesis (Bravo et al. 1987) and is an established S-phase marker (van Dierendonck et al. 1991). Western blotting analysis revealed a strong upregulation of PCNA in extracts of cultured aorta (Figure 2A), also shown by immunocytochemistry (Figure 3). Comparative Western blotting analysis with furin, PC5, and PC7 demonstrated their upregulation as well (Figure 2B). This was characterized by a time-dependent increase of the levels of 87-kD furin, 117-kD PC5, and 80-kD PC7 proteins in arteries cultured for 24 hr as compared to control immunostaining with actin (Figure 2C). Semiquantitative densitometry depicted in Figure 2D for comparison provides additional evidence for the coordinate expression of PCNA and PCs in cultured aorta. Other proprotein convertases, such as PC1 and PC2, were detectable neither on immunoblotting nor by immunocytochemistry (data not shown).
Antibodies used in this study a
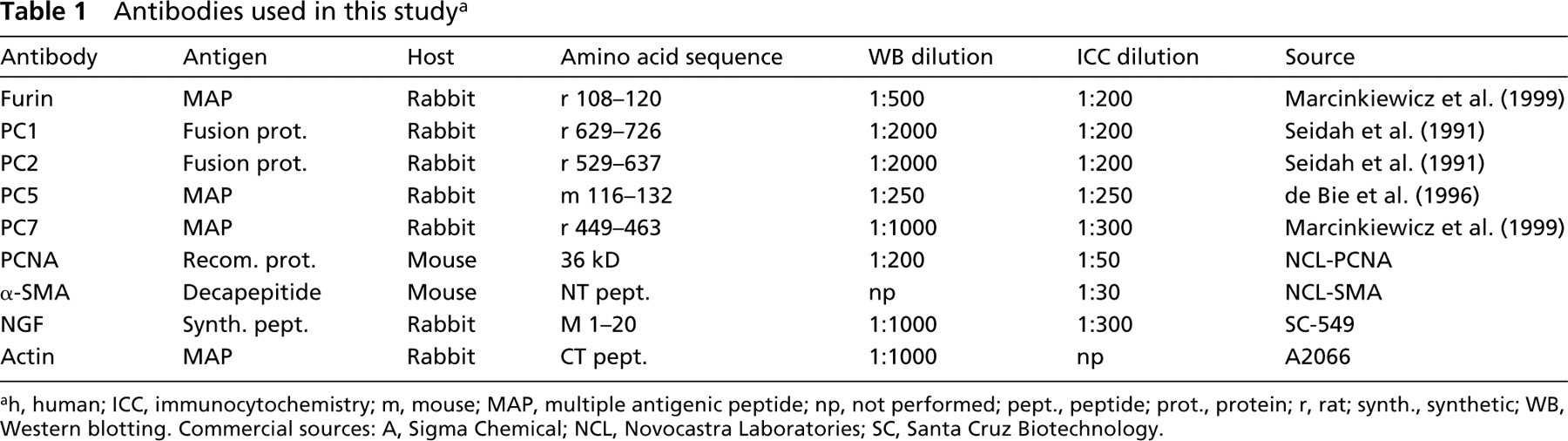
ah, human; ICC, immunocytochemistry; m, mouse; MAP, multiple antigenic peptide; np, not performed; pept., peptide; prot., protein; r, rat; synth., synthetic; WB, Western blotting. Commercial sources: A, Sigma Chemical; NCL, Novocastra Laboratories; SC, Santa Cruz Biotechnology.

Immunocytochemical localization of furin, PC5, and PC7 after in vivo balloon denudation of rat aorta. Rare PC7 (
Immunocytochemical Detection of Proliferating VSMCs in Aorta Explants
Counting of the total cell number in the media revealed no statistically significant difference between intact aorta and explants cultured for 24 hr (Table 2). PCNA immunocytochemistry demonstrated that cell proliferation occurred in clusters in the media of cultured arteries. Quantitative analysis demonstrated a statistically significant increase in PCNA-labeled nuclei density from ~1% in intact vessels to ~35% after 24 hr of culturing. As demonstrated by stainings on consecutive sections (Figures 3C and 3D), PCNA-labeled cells were further identified as VSMCs by their positive immunoreactivity to the cytosolic α-SMA antigen (Mason and Gatter 1987).
Immunocytochemical Localization of PCs in Cultured Rat Aorta and Co-localization with PCNA
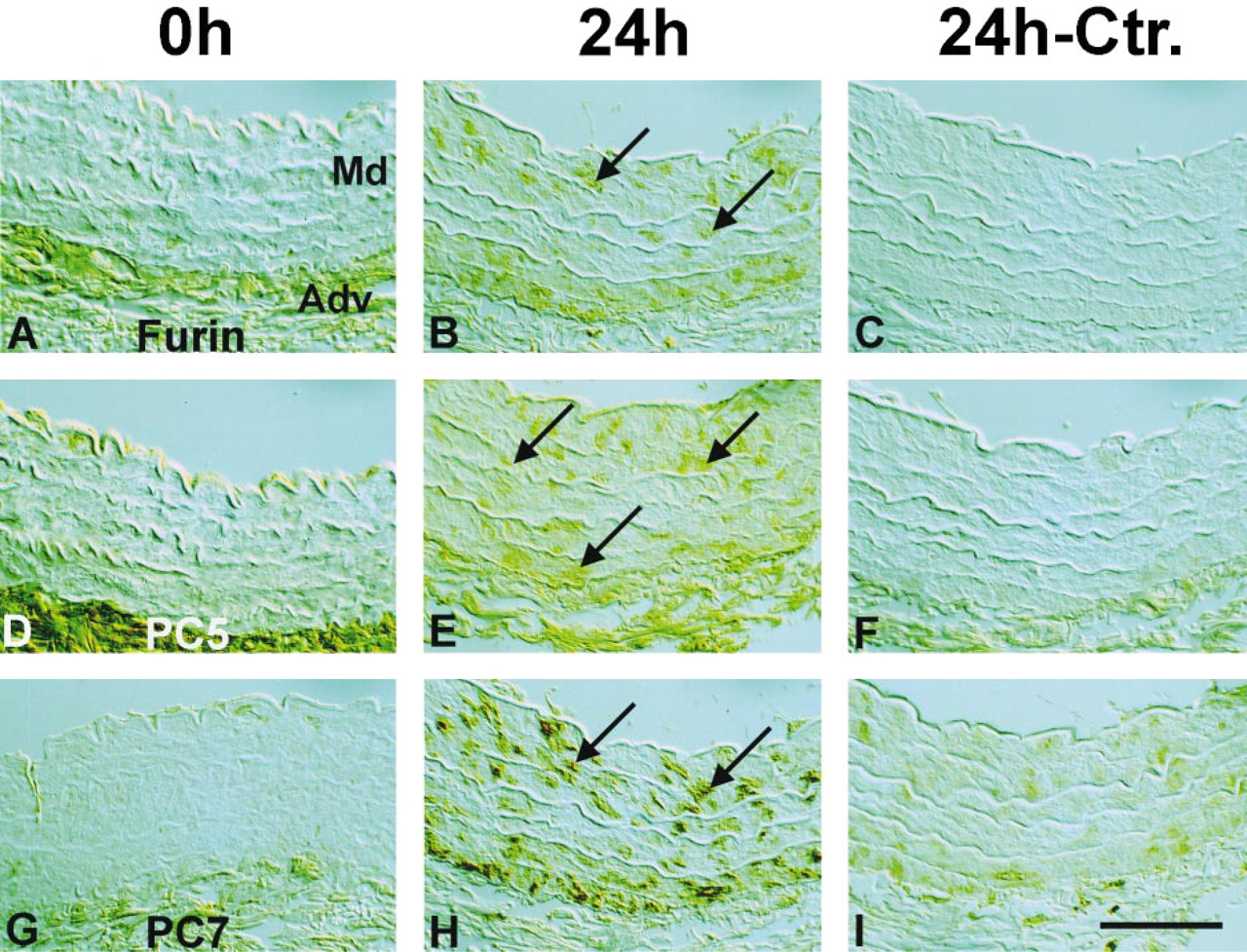
To address the question of whether the upregulation of furin, PC5, and PC7 demonstrated by immunoblotting parallels the pattern of PCNA distribution on immunocytochemistry, we performed immunocytochemical studies using rabbit antisera to furin, PC5, and PC7 (Table 1). Figure 4 shows that, in intact aorta, the expression of all three PCs was mostly confined to the adventitia, whereas after incubation for 24 hr, the distribution pattern of furin, PC5, and PC7 matched that of PCNA labeling observed. Subsequent co-localization studies demonstrated that all PCs were upregulated in proliferating PCNA-positive VSMCs (Figure 5). Quantitative analysis of furin, PC5, and PC7 revealed no statistical difference from PCNA-labeled nuclei after 24-hr incubation of explants (Table 2).
PCs mRNA Regulation During VSMC Proliferation
Northern blotting analysis demonstrated the mRNA upregulation of furin, PC5, and PC7 (Figure 1). Furin mRNA of 4 kb and PC7 mRNA of 3.0 and 3.7 kb increased maximally after 24 hr of incubation. In contrast, PC5 mRNA of 3.5 kb (PC5-A) (Lusson et al. 1993; de Bie et al. 1996) peaked after 4 hr of incubation (Figure 6B). Interestingly, a second PC5 mRNA isoform of 5.5 kb (PC5-B) coding for membrane-anchored PC5 (Lusson et al. 1993) did not display any significant regulation.
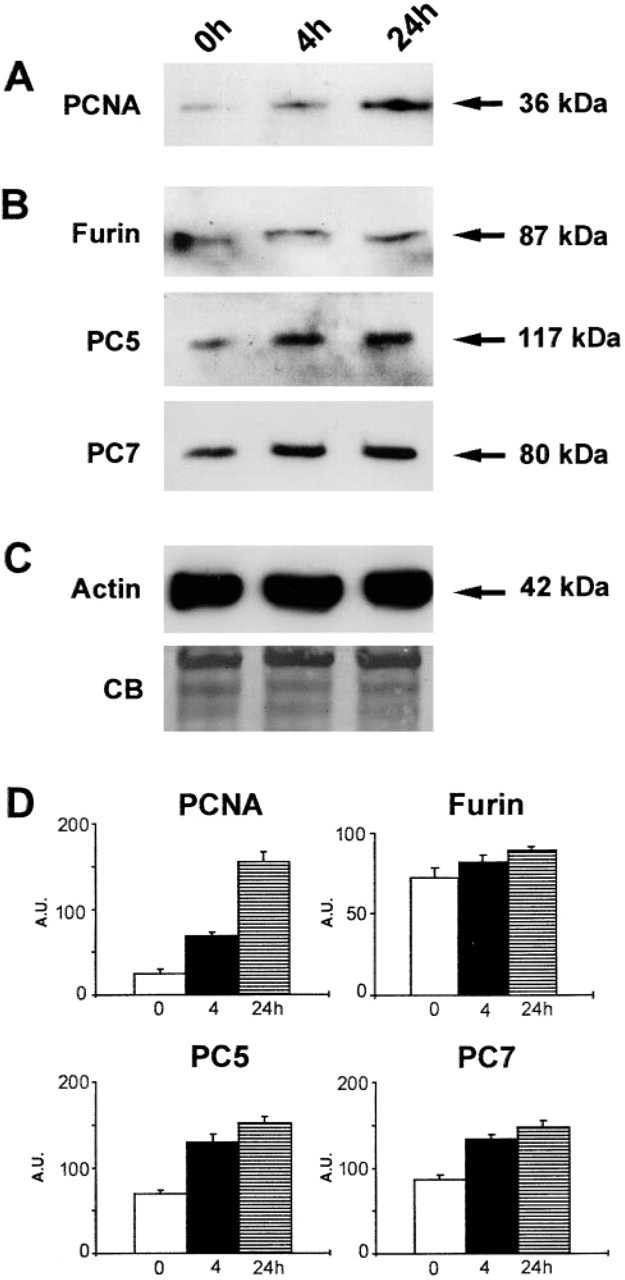
Western blotting analysis showing immunoreactive (
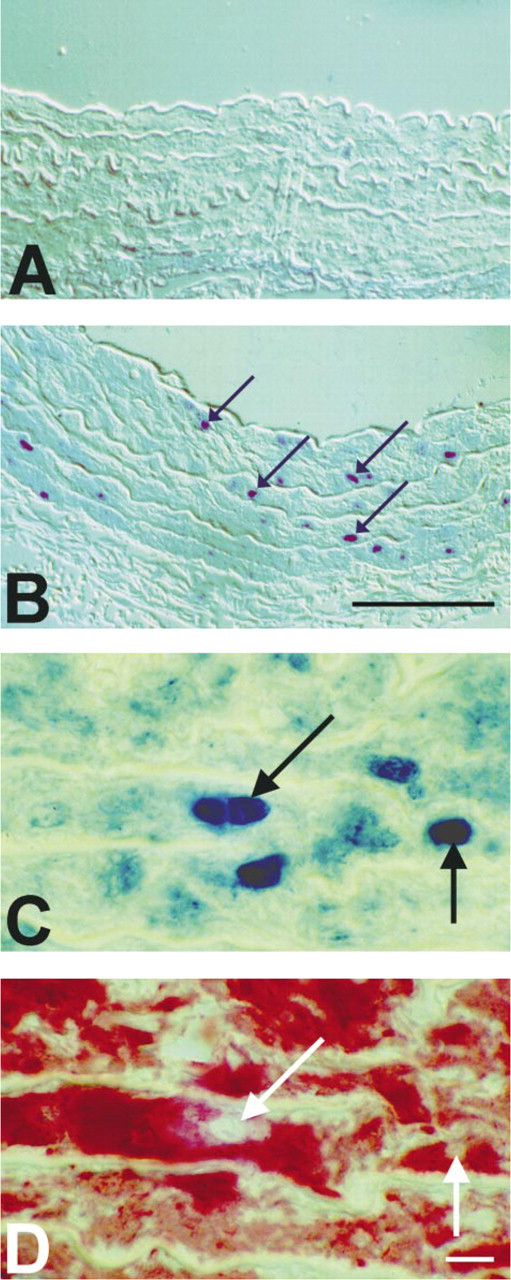
Immunocytochemical detection of PCNA in the media of aorta incubated for 0 hr (
Cell counting in cultured rat aorta a
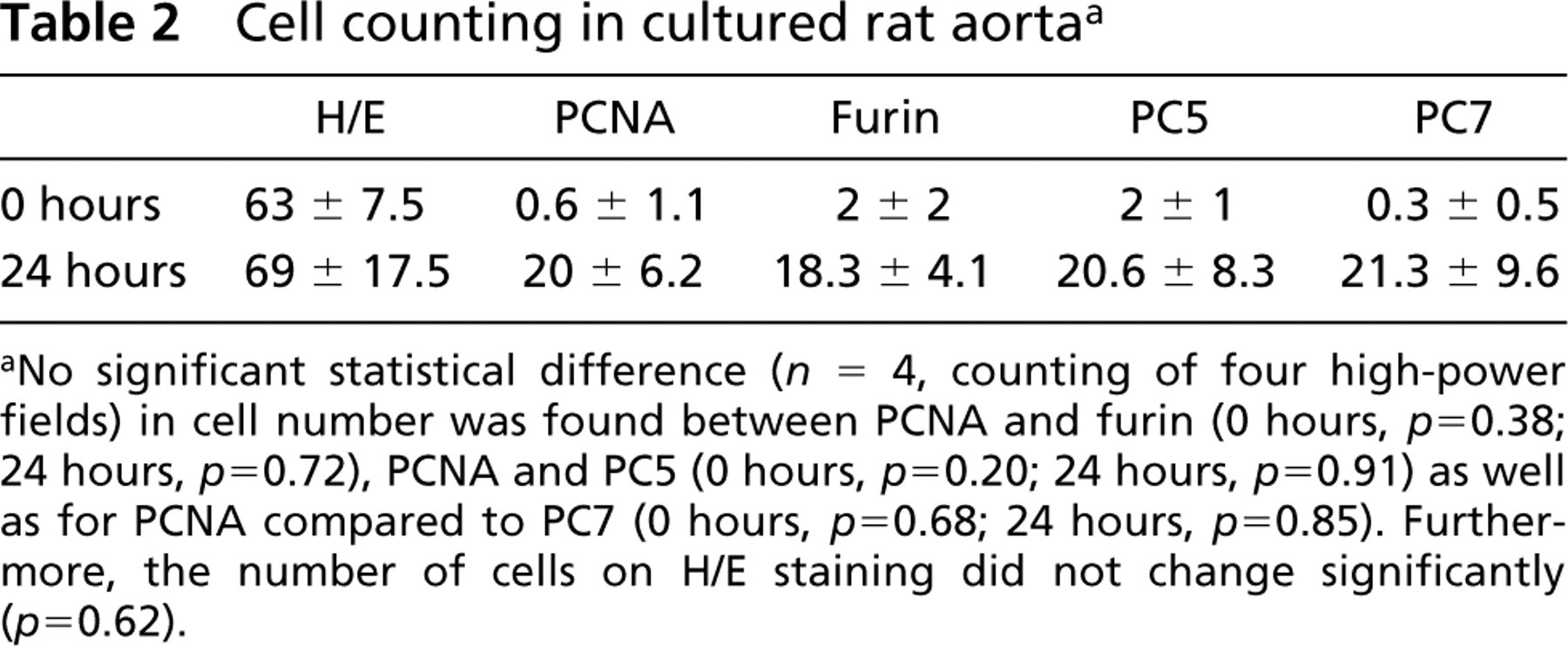
aNo significant statistical difference (n = 4, counting of four high-power fields) in cell number was found between PCNA and furin (0 hours, p = 0.38; 24 hours, p = 0.72), PCNA and PC5 (0 hours, p = 0.20; 24 hours, p = 0.91) as well as for PCNA compared to PC7 (0 hours, p = 0.68; 24 hours, p = 0.85). Furthermore, the number of cells on H/E staining did not change significantly (p = 0.62).
De Novo Synthesis of NGF in Cultured Rat Aorta: Evidence for Release of Growth Factors
To test our hypothesis that the upregulation of PCs in proliferating VSMCs reflects a de novo synthesis of growth factors, we investigated the mRNA and protein regulation of nerve growth factor (NGF) as a model. Among a large number of putative substrates for PCs, NGF is activated by PCs (Bresnahan et al. 1990; Seidah et al. 1996).
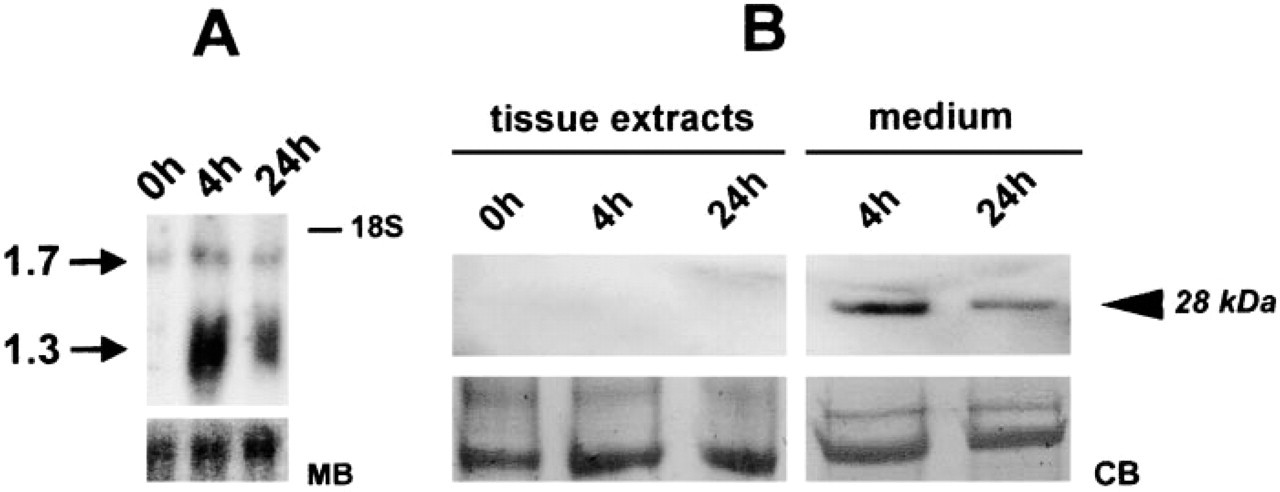
Northern blotting analysis demonstrated upregulation of two NGF mRNA isoforms (1.3 and 1.7 kb). The 1.3-kb NGF mRNA isoform was found to increase markedly after incubation for 4 hr and declined somewhat thereafter (Figure 7). A second NGF mRNA isoform with a molecular weight of 1.7 kb displayed much less regulation.
Attempts were made to co-localize NGF with PCs and/or PCNA by immunocytochemistry. Surprisingly, no NGF immunoreactivity could be produced in aorta explants. The same was true for Western blotting analysis with extracts from control and incubated explants, which were negative as well (Figure 7). Assuming that the absence of NGF in the tissue might result from its release, culture media were subjected to immunoblotting. Our data demonstrate that the upregulation of NGF mRNA on Northern blotting analysis is paralleled by the presence of immunoreactive NGF in explant culture media, because a 28-kD immunoreactive NGF was detected after 4 and 24 hr of incubation (Figure 7B).
Immunocytochemical detection of furin (
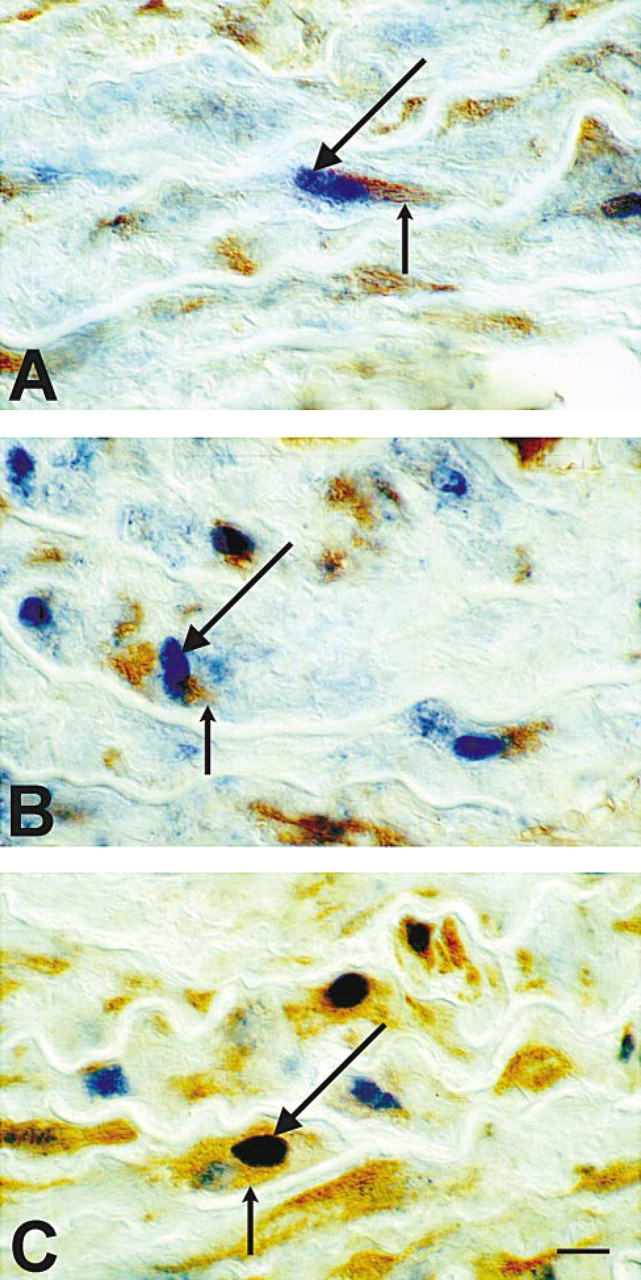
Immunocytochemical co-localization of furin (
Discussion
In this report we demonstrate that the proliferation of VSMCs in vivo and in vitro is accompanied by an up-regulation of several proprotein convertases. In vivo, we demonstrate the strong upregulation of furin, PC5, and PC7 in VSMCs after balloon denudation of rodent arteries. The increase in PCs immunoreactivity paralleled the time course of PCNA staining until a neointima was established, whereas later on furin-, PC5-, and PC7-positive cells outnumbered PCNA-positive cells. This finding suggests that PCs might participate in the very early phase of VSMC proliferation and in later stages of vascular remodeling. To define these events more closely, we investigated the regulation of PCs using an organ culture model of intact rat aorta explants, known to have low levels of VSMC cell divisions (Zeymer et al. 1992; Wei et al. 1997).
Here, we demonstrate a rapid onset of VSMC proliferation in intact arteries incubated over a brief time period (24 hr) without external trophic support. Proliferating VSMCs were identified by PCNA immunostaining, which has been demonstrated to be equivalent to BrdU and [3H]-thymidine labeling in vascular tissue (Zeymer et al. 1992). Even though no statistical significant increases in the total cell number were found during the short incubation time, we observed multifocal centers of PCNA-labeled VSMCs in the media, with no clear anatomic preference or polarity. These centers were not associated with sites of manipulation due to explant preparation. Upregulation of PCNA was paralleled by a de novo protein and mRNA synthesis of furin, PC5, and PC7. Other PCs, such as PC1 and PC2 which are known to be mostly expressed in neuroendocrine cells (Seidah et al. 1998), were not found. Quantitative analysis revealed no statistical difference in PCNA-, furin-, PC5-, or PC7-labeled VSMCs after 24-hr incubation of explants.
Furthermore, co-localization studies demonstrated that increases in furin, PC5, and PC7 immunoreactivity were mostly localized to PCNA-positive VSMCs after 24-hr incubation. Our results indicate that PCs might play an important role in the proliferation of VSMCs. The upregulation of furin, PC5, and PC7 in PCNA-labeled VSMCs indicates that these enzymes may constitute a novel set of gene markers for proliferating VSMCs. The finding of focal areas of PCNA-labeled VSMCs after incubation in our study supports the notion that subsets of VSMCs might initiate proliferation and subsequent changes without trophic support (Schwartz et al. 1990).
Other groups have reported similar results using serum-free organ culture systems of arteries. Hao et al. (1995) found expression of matrix Gla protein (MPG) and osteopontin (OPN) in subsets of medial VSMCs after incubation. MPG and OPN are two genes known to be preferentially expressed in late-passage dedifferentiated VSMCs (Shanahan et al. 1993). In addition, biosynthesis of proteins in serum-free organ cultures of rodent arteries has been demonstrated (Takasaki et al. 1991). Similar to our results, Hosoi et al. (1993) found an induction of an embryonic fibronectin isoform in rat aorta VSMCs appearing in clusters.
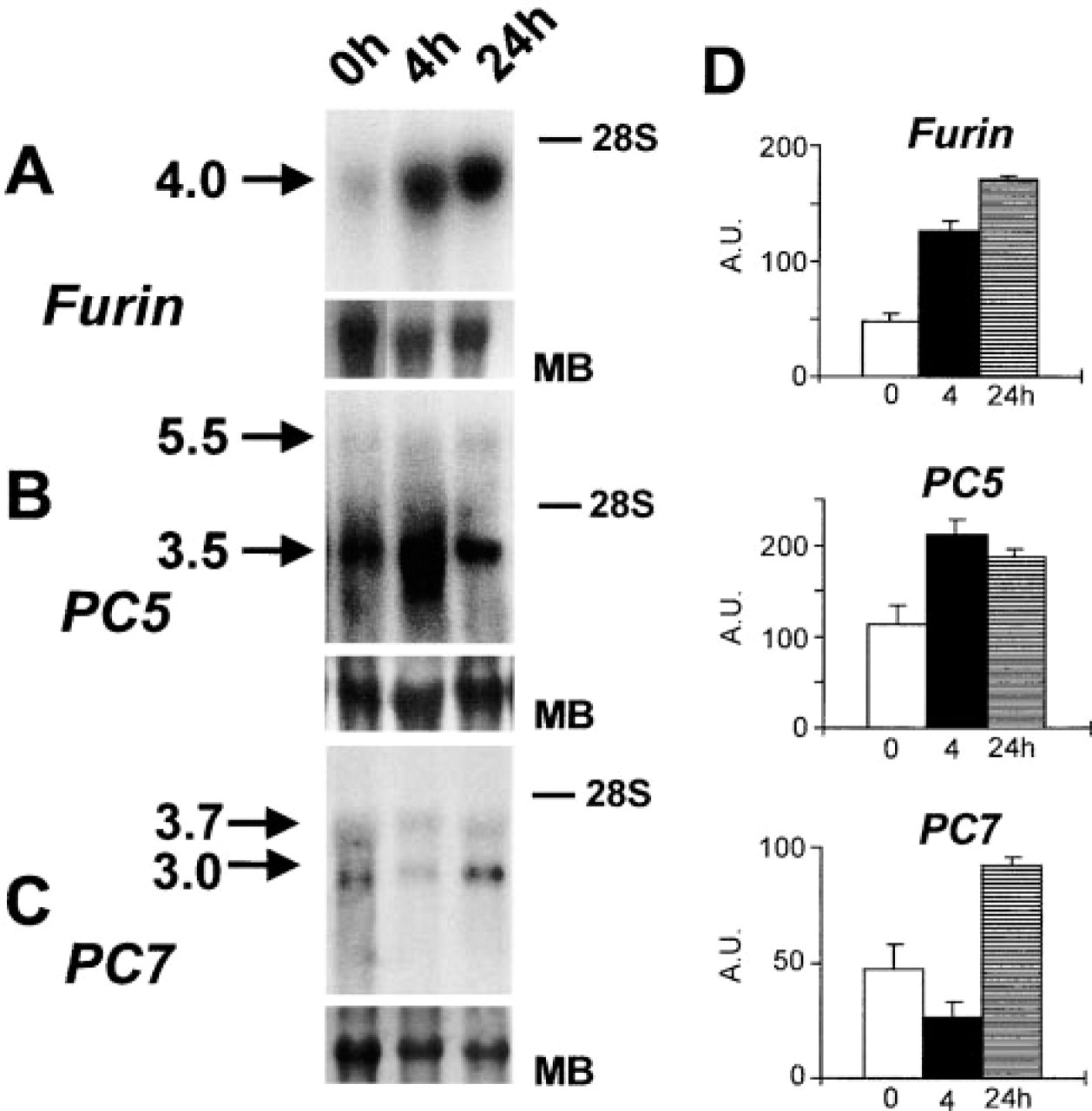
Northern blotting analyses detecting mRNAs of furin (
To test our hypothesis that VSMC proliferation involves the upregulation of growth factors, we attempted to compare their expression with that of nerve growth factor (NGF). Among a large number of putative substrates (Seidah et al, 1998), this growth factor is activated by furin-like enzymes by cleavage at the site RSKR↓ (Bresnahan et al. 1990; Seidah et al. 1996). NGF is synthesized in VSMCs and is a strong VSMC mitogen (Creedon and Tuttle 1991). It has been shown to be preferentially upregulated in neointimal VSMCs after vascular injury in vivo (Donovan et al. 1995), similar to the pattern we describe here for furin, PC5, and PC7 in vivo.
Although strong upregulation of NGF mRNA is evident in rat aorta cultured for 4 and 24 hr (Figure 7), to our surprise no detectable NGF could be found by immunocytochemistry or immunoblotting in aorta explants. Assuming that the strong upregulation of NGF mRNA found on Northern blotting might be paralleled by its release into the culture medium, we examined proteins released from aorta explants. Immunoblot results revealed a release of NGF into the medium after 4 and 24 hr of incubation.
(
In conclusion, this study demonstrates that short-term incubation of intact rat arteries induces significant histomorphological changes. One of these is the rapid onset of VSMC proliferation, paralleled by a selective upregulation of proprotein convertases in these cells. The importance of these enzymes is further supported by our in vivo data. The finding that VSMCs that are immunopositive for PCs outnumber VSMCs that are immunopositive for PCNA in the mature neointima during late stages of vascular remodeling in vivo indicates that they might be involved in the processing of extracellular matrix proteins as well (Raghunath et al. 1999). Accordingly, rates of VSMC replications in advanced human arteriosclerotic plaques have been reported as low (Gordon et al. 1990), but neointimal cells are known to remain metabolically active and display different growth characteristics (Orlandi et al. 1994).
Although the exact role of PCs in proliferating VSMCs has yet to be determined, these enzymes may well be implicated in the development of vascular pathologies.
Footnotes
Acknowledgements
Supported by a grant from the Medical Research Council of Canada (MT-14466). Dr Philipp Stawowy was a fellow of the Charité, Humboldt-University Berlin and German Heart Institute Berlin.
We thank Mr Andrew Chen for technical skills, Dr James Cromlish for review of the manuscript and Ms Sylvie Emond for secretarial assistance.