
Abstract
The rat stomach is rich in endocrine cells. The acid-producing (oxyntic) mucosa contains ECL cells, A-like cells, and somatostatin (D) cells, and the antrum harbours gastrin (G) cells, enterochromaffin (EC) cells and D cells. Although chromogranin A (CgA) occurs in all these cells, its processing appears to differ from one cell type to another. Eleven antisera generated to different regions of rat CgA, two antisera generated to a human (h) CgA sequences, and one to a bovine (b) CgA sequence, respectively, were employed together with antisera directed towards cell-specific markers such as gastrin (G cells), serotonin (EC cells), histidine decarboxylase (ECL cells) and somatostatin (D cells) to characterize the expression of CgA and CgA-derived peptides in the various endocrine cell populations of the rat stomach. In the oxyntic mucosa, antisera raised against CgA291–319 and CGA316–321 immunostained D cells exclusively, whereas antisera raised against bCgA82–91 and CgA121–128 immunostained A-like cells and D cells. Antisera raised against CgA318–349 and CgA437–448 immunostained ECL cells and A-like cells, but not D cells. In the antrum, antisera against CgA291–319 immunostained D cells, and antisera against CgA351–356 immunostained G cells. Our observations suggest that each individual endocrine cell type in the rat stomach generates a unique mixture of CgA-derived peptides, probably reflecting cell-specific differences in the post-translational processing of CgA and its peptide products. A panel of antisera that recognize specific domains of CgA may help to identify individual endocrine cell populations.
C
The ECL cells reside within the basal zone of the oxyntic mucosa and are characterized by their expression of histidine decarboxylase (HDC) and histamine (Håkanson et al. 1986, 1994). Histamine, released from the ECL cells in response to gastrin, is a mediator of acid secretion from the parietal cells (Sandvik and Waldum 1991; Andersson et al. 1996). The function of the A-like cells is unknown. Because no specific histochemical marker for the A-like cells has been reported, their identification is based on ultrastructural criteria (Solcia et al. 1975; Capella et al. 1991; Sundler and Håkanson 1991). D cells are located in both the oxyntic and antral mucosa, whereas G cells and EC cells occur in the antral mucosa only. These cell types are characterized by their content of somatostatin, gastrin, and serotonin, respectively.
The objective of this investigation was to use immunohistochemistry to examine the expression and proteolytic processing of CgA in the different endocrine cell populations of the stomach by employing antisera directed to specific domains of CgA and CgA-derived peptides.
Materials and Methods
Antisera Production
All antisera were raised against rat CgA sequences unless otherwise stated (Figure 1). The generation of eleven of the antisera has been described previously (for references see Table 1). One of these antisera (R880) was raised against a bovine (b) rather than a rat CgA sequence (bCgA82–91). This sequence differs from the corresponding rat sequence in two positions (F in rCgA, instead of Y, in position 82 and F in rat CgA, instead of L, in position 90). Antiserum (R814) was raised against a human (h) CgA sequence (343–356) that differs from the corresponding rat sequence in one position only (R in rCgA instead of K in position 345). An additional three CgA antisera were generated and used here for the first time. These antisera were raised against the C-terminal octapeptide APSKDTVE, representing residues 121–128 of rCgA, Y0SGEATDGARPQALPEPMQESK, representing residues 124–144 of hCgA (Bachem; La Jolla, CA), and the dodecapeptide KVAHQLQALRRG, representing residues 437–448 of rCgA. hCgA124–144 exhibits limited sequence homology with the corresponding rCgA sequence except in the sequence 132–143. Hence, the antiserum was expected to recognize the corresponding sequence in rCgA, which differs from the human in two positions only (F in rCgA, instead of L, in position 136 and K in rCgA, instead of M, in position 140). The peptides were synthesized by solid-phase methods using Fmoc strategy employing an automated peptide synthesizer (Fields and Noble 1990) and were purifed by reverse-phase HPLC. Their molecular mass numbers were verified by fast atom bombardment mass spectroscopy. They were coupled via glutaraldehyde to ovalbumin (rCgA121–128), keyhole limpet hemocyanin (hCgA124–144), and bovine serum albumin (Sigma; St Louis, MO) (rCgA437–448). Rabbits received an SC priming injection of the conjugates in an emulsion of Freund's complete adjuvant (200 μg/ml). A second booster injection [100 μg/ml; Freund's complete:incomplete 1:9 (v/v)] was administered after 6 weeks, followed by additional monthly boosts. The rabbits were bled 2 weeks after each booster injection. Rabbits with code number R876 (antigen rCgA121–128), R36D8 (antigen hCgA124–144), and R881 (antigen rCgA437–448) generated antisera suitable for immunohistochemistry (Table 1; Figure 1).
Details of the antisera used a
aAntisera to CgA fragments were raised in rabbits. All other antisera were raised in guinea pigs except for the serotonin antiserum, R448, which was raised in a rabbit.
Markers for Identification of Individual Endocrine Cell Populations
An antiserum raised in a guinea pig against rat HDC (code no. M9501; Dartsch and Persson 1998; a kind gift from Dr L. Persson, Institute of Physiological Sciences, University of Lund, Sweden), was used as an ECL cell marker (Håkanson et al. 1986, 1994). Guinea pig antisera against rat somatostatin (GP182) (Maule et al. 1990) and rat gastrin (GP360) (Eurodiagnostica; Malmö, Sweden) were used to immunostain D and G cells, respectively. A rabbit antiserum against serotonin (R448) was used to immunostain EC cells. A-like cells were identified in an indirect manner, i.e., as round epithelial cells in the oxyntic mucosa that contained CgA-derived peptides but failed to react with antiserum to either HDC or somatostatin.
Animals
Male Sprague-Dawley rats weighing 200–250 g were sacrificed by exsanguination from the abdominal aorta under chloral hydrate anesthesia (300 mg/kg, IP). The stomach was opened along the greater curvature, washed in 0.9% saline, and placed, mucosal side up, on a cold glass surface. Tissue specimens from the acid-producing part of the stomach were taken from the ventral side close to the rumen. Specimens from the antrum were collected from the ventral side 1–2 mm above the pyloric sphincter.
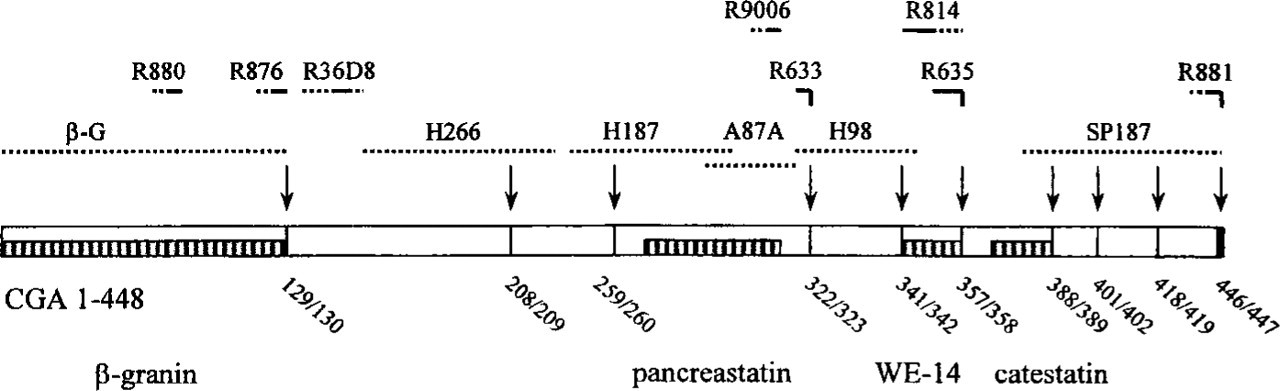
Rat CgA (1–448) with pairs of basic amino acids (representing potential cleavage sites, arrows). Dotted lines indicate CgA sequence used for immunisation and unbroken horizontal lines indicate identified epitopes for the various antisera. The code number of each antiserum is shown above its respective antigen. Well-known CgA-derived peptides are shown for comparison (bottom line).
Immunocytochemistry
Tissue specimens were immersion-fixed in 4% paraformaldehyde in 0.1 M PBS, (pH 7.2) for 20 hr at 4C and cryoprotected in 5% (w/v) and 30% sucrose in 0.1 M phosphate buffer containing 0.01% NaN3 before freezing in OCT embedding medium (Miles; Elkhart, IN). Frozen sections (6 μm) were cut using a Leitz cryostat and incubated (18 hr, 4C) with one of the primary antisera (Table 1). Immunoreaction was visualized using rhodamine-conjugated secondary swine anti-rabbit or fluorescein-conjugated rabbit anti-guinea pig IgG antibodies (1 hr, room temperature). Co-localization studies were performed as described previously (Curry et al. 1991). Briefly, sections were incubated with both primary guinea pig and primary rabbit antisera (Table 1) overnight. The sections were then washed in PBS (10 min) and incubated for 60 min with rhodamine-conjugated swine anti-rabbit antiserum, then with normal rabbit serum (0.1% in PBS, 30 min), followed by fluorescein-conjugated rabbit anti-guinea pig antiserum (60 min). Co-localization of antisera against CgA fragments and the rabbit antiserum against serotonin was examined by serial sectioning (4-μm-thick sections). Immunostaining was examined using a MRC 500 confocal scanning laser microscope (Bio-Rad Lasersharp; Richmond, CA) and an Olympus BH2 light microscope.
Control studies included liquid-phase preabsorption of each antiserum with its respective antigen (Table 1) and with the antigens used to raise the other antisera to CgA-derived peptides, and also the routine omission of primary antisera and substitution with non-immune rabbit, rat, and guinea pig serum. Truncated and C-terminally extended fragments of the respective antigens were synthesized (as above) and used to further characterize the epitope recognized by the various antisera (Table 2). These fragments included ELSEVLE, ELSEVLEN (of antigen YEDELSEVLE), SKDTVE (of antigen APSKDTVE), QELEK (of antigen Y0KGQELE), LGPPQGLFPG-amide, GPPQGLFPG, PPQGLFPG-amide, PQGLFPG-amide and GPPQGLFP (of antigen GPPQGLFPG-amide), WSKMDQLA, SKMDQLAKE, and ELTAEK (of antigens WSKMDQLAKELTAE (WE-14; Curry et al. 1992) and Y0KELTAE), and KVAHQLQALRRG and KVAHQLQALRR (of antigen Y0KVAHQLQALRRG). Fragments of antigen hCGA124–144 were derived by proteolytic cleavage. Briefly, 500 μg of the synthetic peptide Y0SGEATDGARPQALPEPMQESK (hCgA124–144, 500 μg ml–1) was subjected to trypsinization, 0.2 mg ml–1, 14 hr, 37C (Trypsin type XIII, 10,400 BAEE units; Sigma, St. Louis, MO). The digest was subjected to reverse-phase HPLC, employing a C18 column (4.6 × 250 mm) (Supelcosil Technology; Bellefonte, CA) and mass spectrometry with a Voyager-DE Biosystem. Purities >95% were achieved for all fragments. The masses of the peptide fragments were compared with those predicted for cleavage at specific residues. The following five peptide fragments were isolated and identified: Y0SGEATDGAR, Y0SGEATDGARPQ, Y0SGEATDGARPQALPEPM, PQALPEPMQESK, and ALPEPMQESK. Three fragments, Y0SGEATDGARPQALPEPM, PQALPEPMQESK, and ALPEPMQESK, were quantified by radioimmunoassay (Hogg et al. 1998) (the yield was 3, 5 and 11 μg, respectively). The yield of Y0SGEATDGAR and Y0SGEATDGARPQ was approximated (5 and 28 μg, respectively) by comparing the heights of the HPLC peaks (detector absorbance at 214 nm) to that of the synthetic peptide Y0SGEATDGARPQALPEPMQESK.
Results
Specificity of Antisera
No immunostaining was observed after omission of the primary antiserum and the inclusion of nonimmune rabbit or guinea pig serum. Liquid-phase preabsorption of each antiserum with its respective antigen (1 μg/ml, except for R876 100 μg/ml) overnight at 4C abolished immunostaining (Table 2). Preabsorption with antigens used to raise other CgA antisera did not affect immunostaining. Antisera were further characterized after preabsorption with truncated and/or N-terminally or C-terminally extended peptides to determine epitope recognition sites (Table 2). Antiserum R880, raised against bCgA82–91 (YEDELSEVLE), was shown to detect the mid- or C-terminal part of the antigen in that both ELSEVLE and ELSEVLEN abolished immunostaining (10 μg/ml). Immunostaining of antiserum R876 raised against CgA121–128 (APSKDTVE, C-terminal end of β-granin; Hutton et al. 1987) was abolished by SKDTVE (100 μg/ml), suggesting that the antibody recognizes the C-terminus of the antigen. Antiserum R36D8, raised against hCgA124–144 (Y0SGEATDGARPQALPEPMQESK), appears to detect the the mid-portion (ALPEPM) of the antigen in that Y0SGEATDGARPQALPEPM, PQALPEPMQESK, and ALPEPMQESK blocked the staining (1 μg/ml), whereas neither Y0SGEATDGAR nor Y0SGEATDGARPQ did so (nor did the first mentioned three fragments after methionine oxidization). R633, raised against CgA316–321 (Y0KGQELE), is directed against the free C-terminal end (QELE) in that QELEK failed to block the immunoreaction. Antiserum R9006, raised against CgA306–314 (GPPQGLFPG-amide, C-terminal end of pancreastatin; Tatemoto et al. 1986), is directed against the C-terminal octapeptide PPQGLFPG in that LGPPQGLFPG-amide, GPPQGLFPG-amide and PPQGLFPG-amide blocked the reaction, whereas GPPQGLFP and PQGLFPG-amide did not. The antiserum does not require the C-terminal amide because non-amidated GPPQGLFPG blocked the reaction. R814, raised against WE-14 (hCgA343–356, WSKMDQLAKELTAE), recognizes the N-terminal portion of the antigen because SKMDQLAKE, Y0KELTAE, and ELTAEK failed to block the immunostaining [whereas WSKMDQLA did (100 μg/ml)]. R635, raised against the C-terminal region of WE-14 (CgA351–356, Y0KELTAE), is directed against the free C-terminal end of ELTAE in that ELTAEK failed to block the immunoreaction. Likewise, antiserum R881, raised against CgA437–448 (YKVAHQLQALRRG), requires the C-terminal glycine residue for recognition because KVAHQLQALRRG (1 μg/ml), but not KVAHQLQALRR, blocked the reaction.
Characterization of the epitopes recognized by the various CgA antisera employing truncated and N- and C-terminally extended CgA fragments a
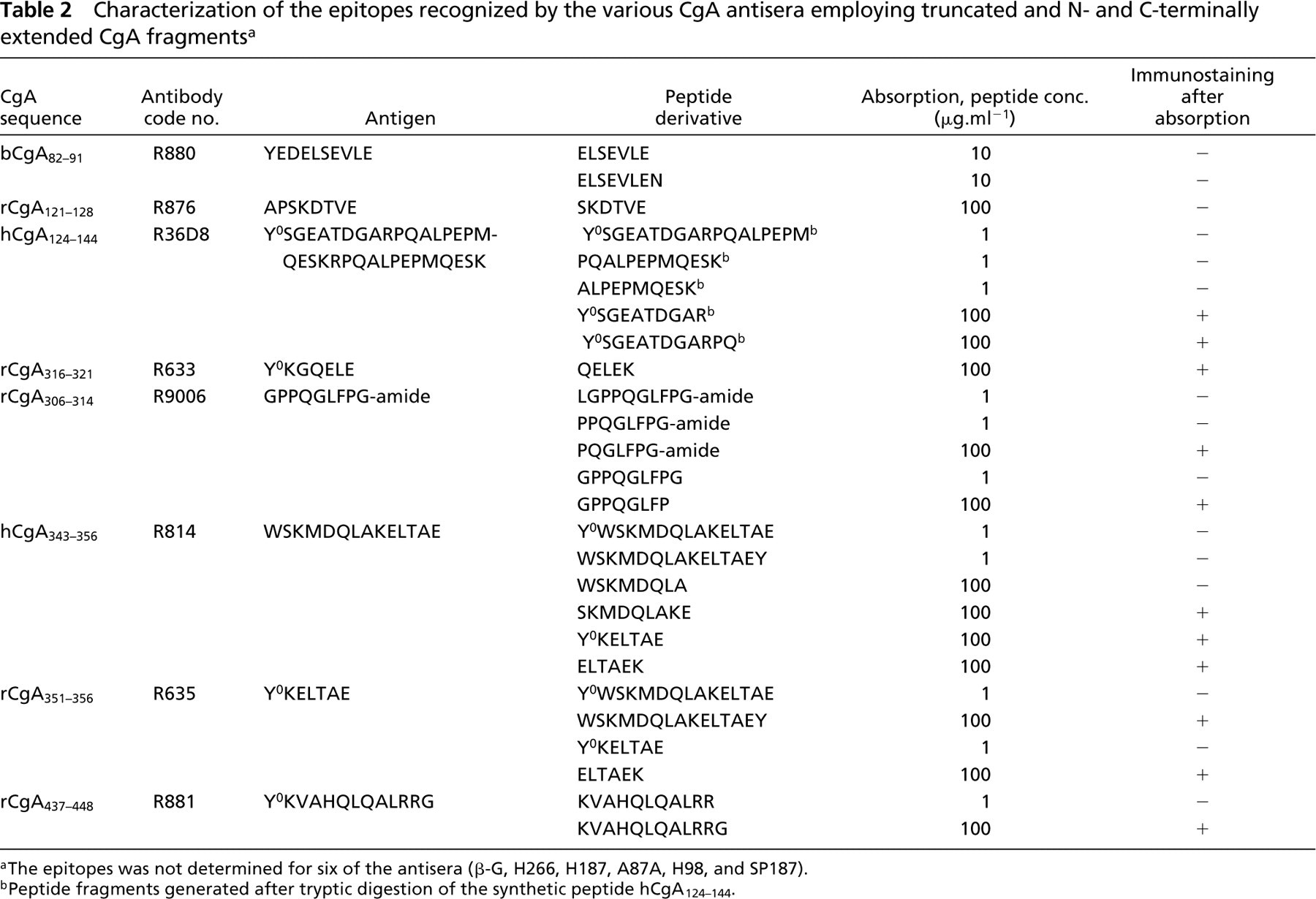
aThe epitopes was not determined for six of the antisera (β-G, H266, H187, A87A, H98, and SP187).
bPeptide fragments generated after tryptic digestion of the synthetic peptide hCgA124–144.
Distribution of CgA Immunoreactivity
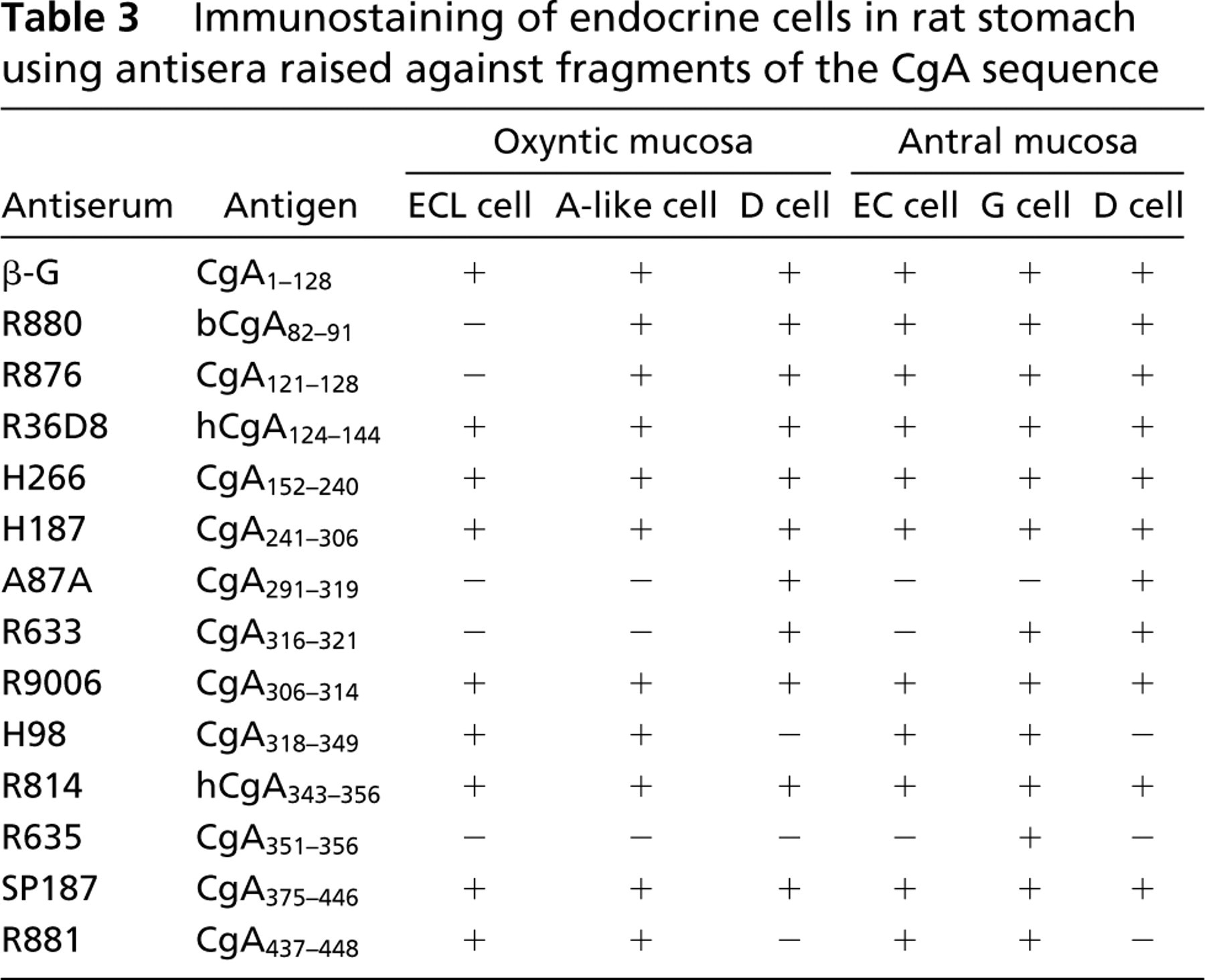
The pattern of immunostaining observed after application of the various CgA antisera to the different endocrine cell populations of the oxyntic mucosa and antrum are summarized in Table 3. Seven of the 14 antisera immunostained all of the major endocrine cell populations (ECL cells, A-like cells, and D cells) in the oxyntic mucosa, and nine immunostained all endocrine cell populations in the antrum (EC cells, G cells, and D cells). The remaining antisera displayed a selective pattern of immunostaining. In the oxyntic mucosa, the ECL cells, A-like cells, and D cells were immunonegative after incubation with antiserum R635 (raised against CgA351–356). However, a discrete population of cells in the oxyntic mucosa, possibly D1/P cells, did exhibit moderate immunostaining. In the antrum, this antiserum immunostained G cells exclusively.
Immunostaining of endocrine cells in rat stomach using antisera raised against fragments of the CgA sequence
Oxyntic Mucosa
ECL Cells. Nine of the antisera immunostained the ECL cells (Table 3), but none of them immunostained the ECL cells exclusively (Figure 2).
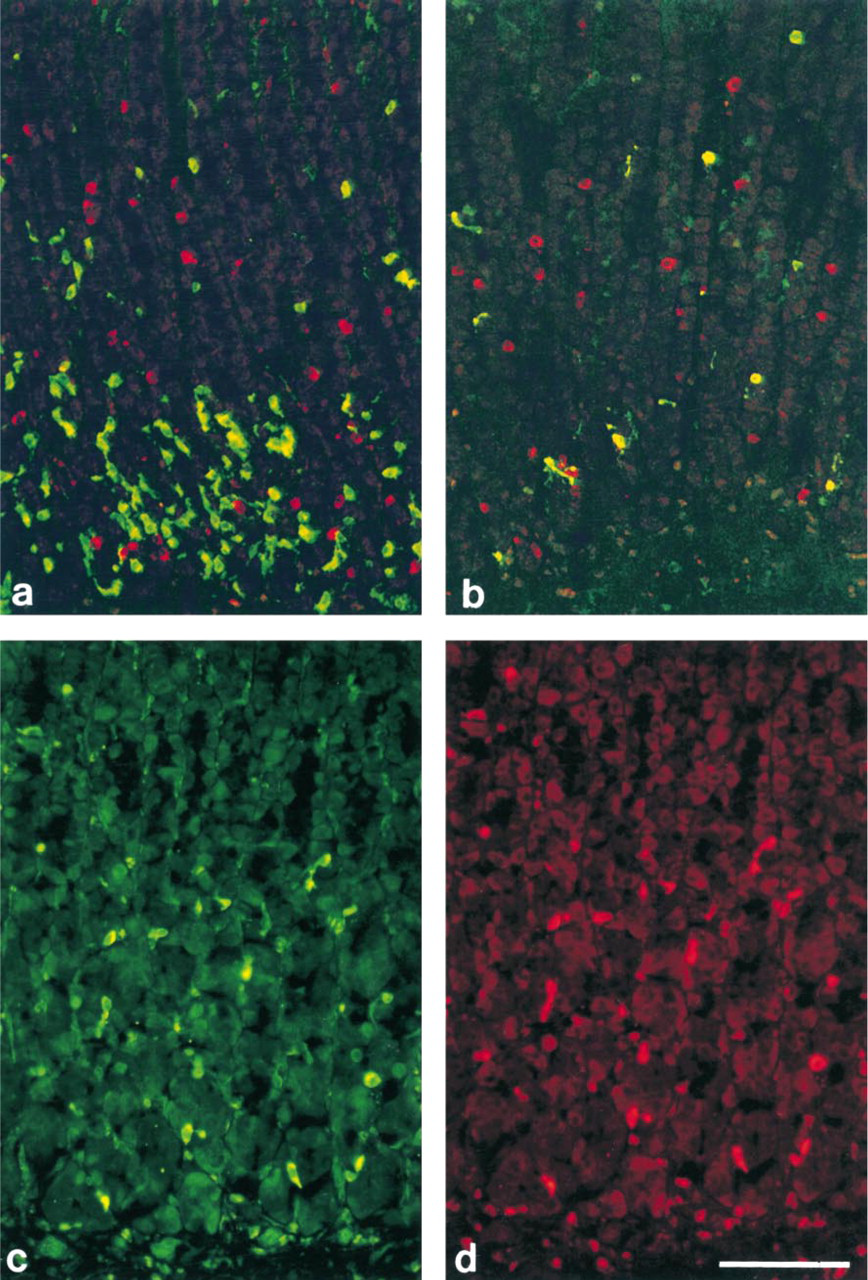
A-like Cells. Antisera R880 and R876 (raised against bCgA82–91 and CgA121–128, respectively) failed to stain the ECL cells, whereas D cells and an additional sub-population of endocrine cells did display staining. Double immunostaining with the somatostatin antiserum revealed that most of these immunopositive cells were distinct from the D cells. The immunopositive cells were larger than D cells, exhibited a distinct spherical shape, and were found throughout the glands of the oxyntic mucosa (Figures 3a and 3b). Their number and morphological features are reminiscent of the A-like cells (Sundler and Håkanson 1991). Eleven of the fourteen antisera immunostained this distinct cell population.
D Cells. Eleven of the 14 antisera demonstrated the D cells. Seven of these also immunostained the ECL cells, and nine immunostained the A-like cells. Three of the 14 antisera, H98 (raised against CgA318–349), R635 (raised against CgA351–356), and R881 (raised against CgA437–448), failed to stain the D cell population. Double immunostaining with the somatostatin antiserum revealed that both A87A and R633 (raised against CgA291–319 and CgA316–321, respectively) immunostained D cells exclusively (Figures 3c and 3d).
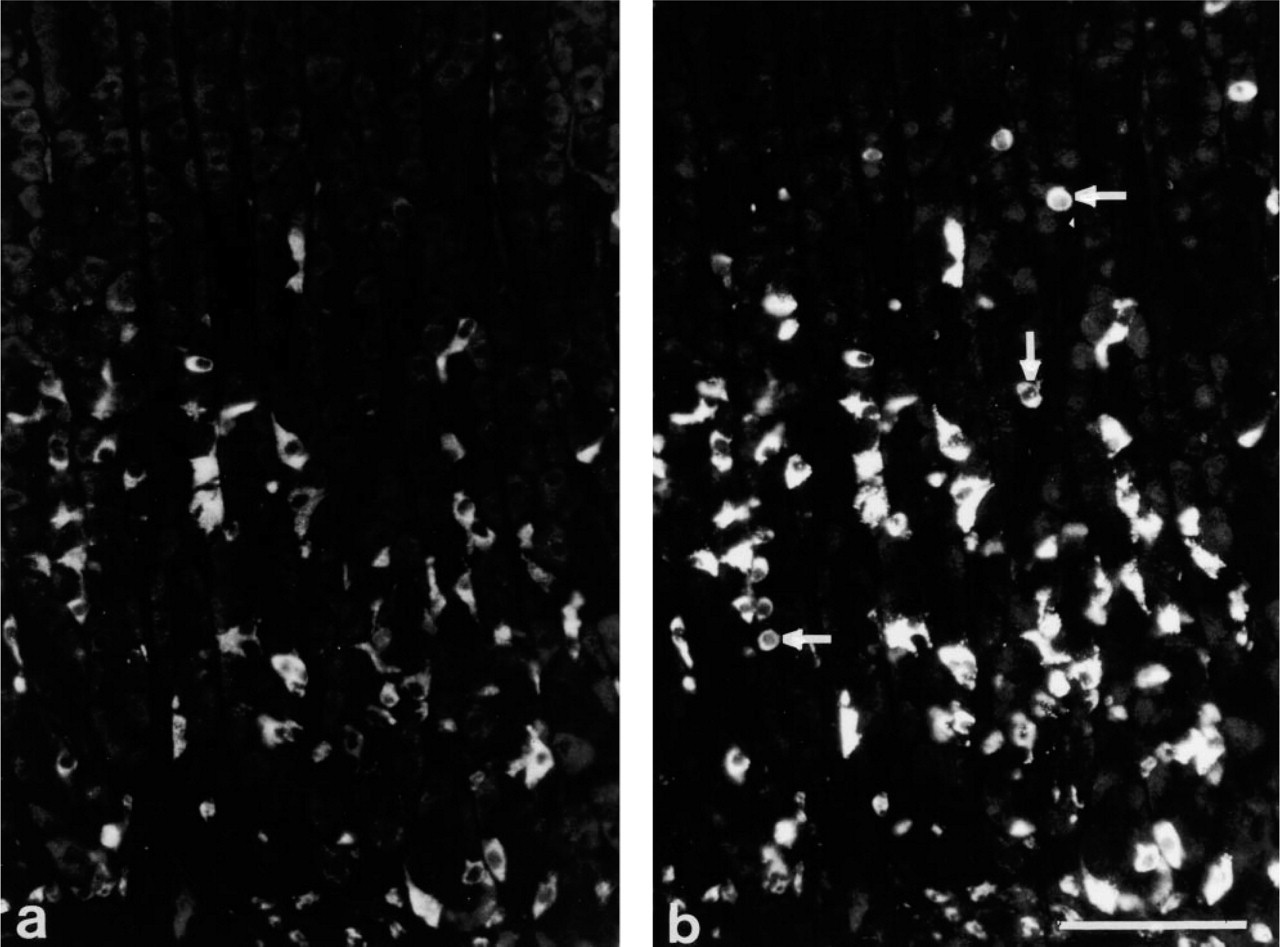
Immunostaining of oxyntic mucosa with antiserum M9501 (raised against HDC) (
(
Antral Mucosa
G Cells. Antiserum R635 (raised against CgA351–356), which failed to stain the ECL cells, A-like, or D cells in the oxyntic mucosa, immunostained the G cells in the antrum. Antiserum A87A (raised against CgA291–319) failed to stain the G cell population. All other CgA antisera immunostained the G cells.
EC Cells. None of the antisera immunostained EC cells exclusively. Most antisera immunostained the G, D, and EC cell populations.
D Cells. As in the oxyntic mucosa, A87A (raised against CGA291–319) immunostained D cells exclusively. Antisera H98, R635, and R881 (raised against CgA318–349, CgA351–356 and CgA437–448, respectively), failed to stain D cells in the antrum as well as in the oxyntic mucosa.
Discussion
Both light microscopic and electron microscopic techniques have been employed to localize CgA and CgA-derived peptides to gastric endocrine cells (Facer et al. 1985; Varndell et al. 1985; Håkanson et al. 1986; Rindi et al. 1986; Buffa et al. 1988; Grube et al. 1989; Lamberts et al. 1990; Curry et al. 1991; Portela-Gomes et al. 1997). Although CgA has been claimed to be a “marker” common to all neuroendocrine cells, endocrine cells of the gut are known to display heterogeneity of CgA immunoreactivity (Cetin et al. 1989). Variation in CgA immunoreactivity has been noted from one cell type to another and from one species to another. Clearly, the biological role of CgA is not uniform among these various cell populations. These variations may reflect the expression of different sets of processing enzymes. Alternatively, the enzymes might be the same but their rate of processing may differ greatly between the various cell types. CgA is subjected to post-translational modifications such as proteolysis and peptide amidation, glycosylation, and phosphorylation (Tatemoto et al. 1986; Winkler et al. 1986; Gorr and Cohn 1990; Barbosa et al. 1991). If the post-translational modifications are cell-specific, different epitopes will be expressed in different cell types, which may explain why certain antisera immunostain some endocrine cell types but not others. Moreover, the post-translational modifications may have a significant impact on the substrate recognition of the proteolytic enzymes and hence on which CgA-derived peptides are generated in the different cell populations. However, discrepancies among reports from different laboratories may also reflect different methodology (fixation and further processing of samples; Van Ewijk et al. 1984) and/or differences among the antisera applied rather than cell-specific differences in post-translational processing. To exclude methodological variables, all tissue samples in the present study were fixed and processed in a standardized way, thus facilitating a direct comparison of the various gastric endocrine cell types with one another. Fourteen site-specific CgA antisera were employed, together with cell-specific markers, to characterize the CgA immunoreactants in the different endocrine cell populations of the rat stomach. Previous investigations have reported differences in the expression of CgA-derived peptides in endocrine cells of the gastrointestinal tract. However, the various endocrine cell populations were not identified (Curry et al. 1991; Watkinson et al. 1991).
Oxyntic Mucosa
The ECL cells represent the predominant endocrine cell type of the oxyntic mucosa. In the rat stomach, histamine is found in both mast cells and ECL cells (Håkanson et al. 1986; Nissinen and Panula 1993), but only the ECL cells contain immunoreactive HDC (Kubota et al. 1984; Håkanson et al. 1986; Andersson et al. 1996). Nine of the 14 antisera to the different CgA domains displayed co-localization of CgA and CgA-derived peptides with HDC. However, each of these antisera immunostained additional cell populations in the oxyntic mucosa. Therefore, none of the antisera demonstrated the ECL cells exclusively.
The somatostatin-secreting D cells are spindle-shaped cells with long slender projections (Alumets et al. 1979; Kusumoto et al. 1979; Larsson et al. 1979). In the oxyntic mucosa they are uniformly distributed throughout the crypts and are of the closed type. Previous studies failed to detect CgA-derived peptides in gastric D cells (Varndell et al. 1985; Buffa et al. 1988; Wiedenmann and Huttner 1989; Portela-Gomes et al. 1997). This might reflect the fact that D cells produce only small amounts of CgA or that the specificity of the antisera used was such that they failed to detect the epitopes presented by the D cells. In this study, 11 of the antisera immunostained the D cells as well as other endocrine cell types. Two antisera, R633 (raised against CgA316–321) and A87A (raised against CgA291–319), immunostained the D cells exclusively. Antiserum R633 is specific to the free C-terminus of CgA316–321, which is generated by cleavage of dibasic amino acid residues CgA322–323. It therefore appears that cleavage at this site occurs in D cells but not in other endocrine cells in the oxyntic mucosa, suggesting that D cells process CgA differently from ECL cells and A-like cells.
To date, there is no recognized immunohistochemical marker for A-like cells, and their identification is based on indirect criteria. A-like cells are CgA-immunoreactive cells that do not display either HDC or somatostatin immunoreactivity and that exhibit a characteristic spherical shape and occur dispersed throughout the gastric glands in lower numbers than the ECL cells. Eleven of the CgA antisera immunostained this endocrine cell population. However, no single antiserum immunostained these cells exclusively. Two antisera, R880 (raised against CgA82–91) and R876 (raised against CgA121–128), which were raised against different regions of the N-terminal CgA domain (CgA1–128), referred to as β-granin (Hutton et al. 1987), immunostained D cells and the A-like cells. In contrast, another antiserum (β-G), raised against full-length β-granin, immunostained all endocrine cell populations in this location. Antiserum H98 and the C-terminal antiserum R881 immunostained ECL cells and the putative A-like cells but not the D cells. Again, this differential pattern of immunostaining supports the concept of cell-specific proteolysis of CgA. Antiserum R635, which is specific for the free C-terminus of WE-14 (CgA343–356; Curry et al. 1992), immunostained the G cells (see below) but failed to immunostain either ECL cells, A-like cells, or D cells in the oxyntic mucosa, suggesting that neither of these cell populations is able to produce WE-14.
Antrum
The predominant endocrine cell populations of the antral mucosa, the G cells, EC cells, and D cells, are localized in the basal part of the glands. All three endocrine cell populations are of the “open” type. Antiserum R635, specific for the C-terminal end of WE-14, immunostained G cells but not EC cells or D cells. This observation suggests that WE-14 is generated in G cells only.
Antral D cells were immunostained by 11 of the 14 antisera raised against CgA. One antiserum, A87A (raised against CgA291–319), immunostained the antral (and oxyntic) D cells but no other endocrine cells. R633 (raised against CgA316–321) immunostained antral (and oxyntic) D cells and G cells. Antiserum R36D8 (raised against CgA124–144) immunostained EC cells and D cells but not G cells. The fact that the D cells of both the oxyntic mucosa (closed type) and the antrum (open type) displayed identical patterns of CgA immunostaining is notable in view of the fact that they appear to differ from each other both morphologically and functionally (Alumets et al. 1979; Olesen et al. 1987).
Eleven of the 14 antisera immunostained the EC cells. The EC cell pattern of CgA immunostaining was identical to that of the A-like cells in the oxyntic mucosa, implying similarities in biosynthesis and processing of CgA.
Conclusions
In this study, 14 well-characterized CgA antisera were tested to generate information on the extent of cell-specific processing in the endocrine cells of the rat stomach. All major endocrine cell types were found to express CgA or CgA-derived peptides. In addition, the application of region-specific antisera raised against a range of CgA-derived peptides has revealed that each endocrine cell type expresses a unique mixture of CgA-derived peptides. This implies that each endocrine cell type has a characteristic spectrum of processing enzymes, e.g., prohormone convertase, carboxypeptidase, and amidating enzymes. It cannot be excluded, however, that additional differences with respect to glycosylation, phosphorylation, sulfation, or the rate and degree of processing will influence the ability of the processed products to react with the antisera. We propose that antisera that recognize specific domains of CgA be used as tools to identify individual endocrine cell populations. This may aid in the identification of cell populations for which no cell-specific markers exist or which, for some reason, are difficult to immunostain with a cell-specific marker (such as D cells and G cells in somatostatin or gastrin knockout animals). The functional significance of the present findings may perhaps be discussed in relation to the prohormones produced by the various endocrine cell types (such as preprogastrin in G cells or prosomatostatin in D cells); the CgA processing pattern may predict how the prohormones are being processed.
Footnotes
Acknowledgments
Supported by grants from the Swedish MRC (04X-1007), the Cancer Foundation (2542-B95–07XCC), the A. Påhlsson Foundation, the Medical Faculty, (University of Lund; Lund, Sweden), and Ipsen Pharmaceuticals, (London, UK).