
Abstract
The epithelial cells of the testis are involved in the production, differentiation, and sustenance of sperm, and those of the epididymis play a major role in sperm maturation, protection, and storage. These tissues express various proteins that respond differently to androgens. Cathepsin A is a multifunctional lysosomal carboxypeptidase that also functions as a protective and an activator protein for neuraminidase and β-galactosidase. In this study, cathepsin A was immunolocalized by light and electron microscopy using a polyclonal affinity-purified antibody on the testis and epididymis of normal, orchidectomized with or without testosterone supplementation, efferent duct-ligated, and hypophysectomized adult rats. In normal rats, cathepsin A expression was noted in lysosomes of Sertoli and Leydig cells but not in germ cells of the testis, as well as nonciliated cells of the efferent ducts. In the epididymis, a cell- and region-specific distribution of cathepsin A was noted. In experimentally treated animals, no changes were noted in the expression of cathepsin A. Immunolabeling of tissues examined at the electron microscopic level revealed that lysosomes were reactive. These data indicate cell- and region-specific expression of cathepsin A in cells of the testis and epididymis and also indicate that cathepsin A expression is not regulated by testicular or pituitary factors. (J Histochem Cytochem 48:1131–1146, 2000)
The lysosomal protective protein/cathepsin A (cathepsin A) is a serine acid carboxypeptidase present in lysosomes, where it associates with and forms a fully functional and stable high molecular weight multienzyme complex with β-d-galactosidase and N-acetyl-α-neuraminidase (d'Azzo et al. 1982; Verheijen et al. 1982; Hoogeveen et al. 1983; Yamamoto and Nishimura 1987). Cathepsin A as a 54-kD precursor is probably targeted to lysosomes via the mannose-6-phosphate receptor, where it is processed into its mature 32/20-kD, disulfide-linked two-chain form (Galjart et al. 1988; Zhou et al. 1996). One role of cathepsin A is facilitation of the intracellular routing, lysosomal localization, and activation of neuraminidase (Bonten et al. 1996; van der Spoel et al. 1998). Cathepsin A also protects β-galactosidase against rapid proteolytic degradation (d'Azzo et al. 1982; Morreau et al. 1992). As a member of the serine protease family, cathepsin A exerts its own catalytic activity at an acidic pH and its esterase/C-terminal deamidase activity at a neutral pH, both functions being fully separable from its protective function (Galjart et al. 1991). The activity of cathepsin A is essential for the complete degradative procession of selected bioactive peptides within lysosomes, including substance P, oxytocin, and endothelin I (Jackman et al. 1990, 1992; Hanna et al. 1994; Itoh et al. 1995; Rottier et al. 1998).
Cathepsin A deficiency in humans causes a lysosomal storage disease called galactosialidosis, which is characterized by the deficiency of both neuraminidase and β-galactosidase. The importance of lysosomal enzymes as constituents of lysosomes is emphasized in lysosomal storage diseases in which there is an increase in the number, volume, and distribution of lysosomes, resulting in abnormalities to cells affected and serious consequences to the health of the animal (Neufeld et al. 1975; Gravel et al. 1995; Hammel and Alroy 1995; Phaneuf et al. 1996; Trasler et al. 1998).
We have previously examined the differential expression of several lysosomal enzymes within lysosomes of various cell types of the testis and epididymis (Hermo et al. 1992a, 1997; Igdoura et al. 1995). Each of these enzymes showed a cell-specific distribution within the testis. Similarly, the cellular distribution in the epididymis varied for each lysosomal enzyme, and a region-specific distribution per given cell type was also observed. This suggested that the lysosomes of these different cell types acted on different substrates that were derived from the endocytosis of substances from the epididymal lumen and in this way created an environment suitable for sperm differentiation in the testis and maturation in the epididymis. The expression of cathepsin A was investigated by Rottier et al. (1998) and Sohma et al. (1999) using in situ RNA hybridization and immunocytochemistry in Sertoli and Leydig cells of the testis and in epithelial cells of the epididymis. Their studies, however, did not examine the various cell types involved and the region-specific distribution of cathepsin A expression in the epididymis, nor did the authors perform electron microscopic or regulation studies.
The purpose of the present study was to immunolocalize cathepsin A in the different cell types of the testis and epididymis of normal adult rats at the light and electron microscopic levels to determine its cell- and region-specific distribution in these tissues. Orchidectomy studies with and without testosterone replacement, ligation of the efferent ducts, and hypophysectomy were used to verify the role, if any, of testicular and pituitary factors in the expression of cathepsin A in these tissues.
Materials and Methods
Animals and Protocols
Adult male Sprague–Dawley rats (350–450 g) were obtained from Charles River Laboratory (St-Constant, QC, Canada). The animals were subdivided into six groups. The first group consisted of normal untreated animals. Bilateral ligation of the efferent ducts constituted the second group. After an IP injection of sodium pentobarbital (Somnitol; MTC Pharmaceuticals, Hamilton, ONT, Canada), the testes and epididymides of four rats were exposed through an incision of the anterior abdominal wall. A ligature was placed around both right and left efferent ducts at a site close to the rete testis. The animals were sacrificed at 3, 7, 14, or 21 days after surgery. Bilateral orchidectomy constituted the third group. After anesthesia, both testes of four rats were removed after a ligature was placed around the efferent ducts and testicular blood vessels. The animals were sacrificed at 3, 7, 14, or 21 days after surgery. Bilaterally orchidectomized rats that received three 6.2-cm testosterone-filled implants constituted the fourth group. Testosterone-filled polydimethyl-siloxane (silastic) implants were prepared according to the method of Stratton et al. (1973) and have well-characterized steroid release rates (Brawer et al. 1983). After anesthesia, both testes were removed from four rats and the implants placed SC immediately after orchidectomy. The rats were sacrificed at 3, 7, 14, or 21 days after surgery.
The fifth group consisted of hypophysectomized rats with four rats per group being sacrificed at 7, 14 21, or 28 days after hypophysectomy. The sixth group consisted of four sham-operated animals, two of which received three empty 6.2-cm-long implants, with all rats being sacrificed 14 days after initiation of the experiment.
All experimentation was carried out with minimal stress and discomfort to the animals both during and after surgery, as set up by the guidelines and approval of the University Animal Care Committee.
Tissue Preparation for LM Immunocytochemistry
At the end of each experiment, the testes (normal and hypophysectomized animals) and epididymides (all procedures) of each rat were fixed by perfusion with Bouin's fixative via the abdominal aorta for 10 min. After perfusion, the testes and epididymides were removed. The latter were cut so that given sections would include all of the major regions of the epididymis, i.e., the initial segment, intermediate zone, caput, corpus, and cauda (Hermo et al. 1991). The tissues were then immersed in Bouin's fixative for 72 hr, after which they were dehydrated and embedded in paraffin.
LM Immunostaining
Sections 5 μm thick were cut and mounted on glass slides. They were then deparaffinized with xylene and hydrated in graded concentrations of ethanol (from 100% to 50%). During hydration, residual picric acid was neutralized by immersing the tissues in 70% ethanol containing 1% lithium carbonate for 5 min. To inactivate any endogenous peroxidase activity, the tissue sections were incubated for 5 min in 70% ethanol containing 1% (v/v) hydrogen peroxide. After hydration, the sections were incubated (5 min) in a 300 mM glycine solution to block free aldehyde groups. The tissue was then blocked with 40 ml of 10% goat serum diluted in TBS (20 mM Tris-HCl–saline containing 0.1% bovine serum albumin) at pH 7.4 for 25 min at room temperature. The slides were then washed with Tween buffer solution (TBS with 0.1% Tween-20) (TWBS).
Light micrographs of seminiferous tubules of the testis at different stages of the cycle immunostained with anti-cathepsin A antibody. (
A dilution factor of 1:100 in TBS was used for the polyclonal anti-cathepsin A antibody. The antibody was provided by Dr. Y. Suzuki (Tokyo Metropolitan Institute of Medical Science; Tokyo, Japan), and its characterization and specificity have been documented in Satake et al. (1994). Each tissue section was incubated in the primary antibody for 1.5 hr. After incubation, the sections were washed by immersing them in four consecutive wells of TWBS for 2 min each. The sections were then blocked with 40 ml of 10% goat serum and incubated with goat anti-rabbit IgG conjugated to peroxidase (Sigma; St Louis, MO) at a dilution of 1:250 in TBS and incubated for 30 min at 35C in a humidified incubator. After incubation with a secondary antibody, the tissue was washed by immersion in four wells of TWBS for 2 min each.
The final reaction product was obtained by incubating the slides for 10 min in 250 ml of TBS containing 0.03% hydrogen peroxide, 0.1 M imidazole, and 0.05% diaminobenzidine tetrahydrochloride (DAB), pH 7.4. The sections were counterstained with 0.1% methylene blue (2 min) and then dehydrated in a graded series of ethanol solutions (30 sec each) and xylene (3 min). Coverslips were mounted on glass slides with Permount. Incubation with normal rabbit serum at a dilution of 1:100 in TBS and incubation of tissues in secondary antibody alone served as controls.
EM Immunocytochemistry
Four additional adult male Sprague–Dawley rats (350–450 g) obtained from Charles River Laboratories were anesthetized with an IP injection of sodium pentobarbital (Somnitol). The testes and epididymides of the rats were fixed by perfusion through the abdominal aorta for 10 min with 0.5% glutaraldehye in 0.1 M phosphate buffer containing 50 mM lysine at pH 7.4. After fixation, the testes and epididymides were removed, cut into small pieces (1 mm3), immersed for 2 hr in the same fixative at 4C, washed with 0.15 M PBS containing 4% sucrose (pH 7.4) at 4C, and then treated with PBS containing 4% sucrose and 50 mM NH4Cl for 1 hr at 4C. The tissue was then washed, dehydrated in graded methanol up to 90%, and embedded in LR White.
Ultrathin sections of selected testicular and epididymal regions were mounted on 300-mesh formvar-coated nickel grids. Each section was floated for 15 min on a drop of 20 mM Tris-HCl-buffered saline containing 5% bovine serum albumin (TBS) and then incubated for 1 hr on 15- μl drops of a polyclonal anti-cathepsin A antibody diluted 1:5 in TBS. Sections were washed four times for 5 min each in TWBS, transferred for 15 min to drops of TBS containing 5% bovine serum albumin, and incubated for 30 min on 20- μl drops of goat anti-rabbit IgG antibodies conjugated to 15-nm colloidal gold particles. The sections were subjected to three 5-min washes in TWBS followed by 5-min washes in distilled water. Sections were counterstained with uranyl acetate in 30% ethanol and lead citrate. Photographs were taken on a Philips 400 electron microscope. Normal rabbit serum at a dilution of 1:5 served as a control.
Results
Light Microscopy
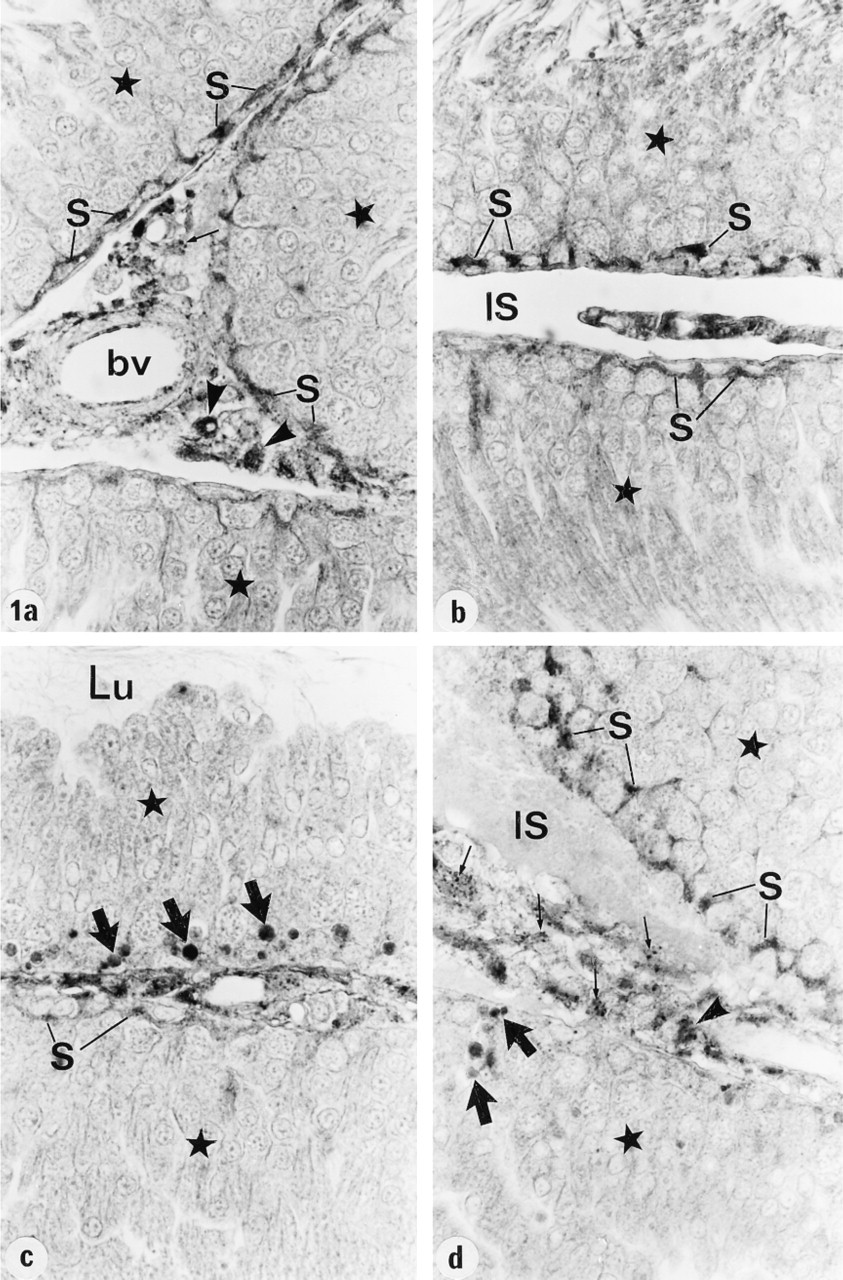
In the testis, expression of cathepsin A was noted in Sertoli cells at all stages of the cycle of the seminiferous epithelium. Whereas the immunoperoxidase reaction was intense in the basal region of Sertoli cells at Stages I–VIII and XIII-XIV, it was weak at Stages IX-XII (Figures 1a-1d). There was no apparent staining of germ cells at any stage of the cycle. Late residual bodies, as judged by their position near the base of seminiferous epithelium at Stages IX-XII, were intensely reactive (Figures 1c and 1d), but early residual bodies situated closer to the lumen were unreactive. In the interstitial space, Leydig cells were identified as those cells showing several intensely reactive lysosomes, whereas macrophages known to contain many lysosomes appeared to be represented by those cells showing intense cytoplasmic staining (Figures 1a and 1d).
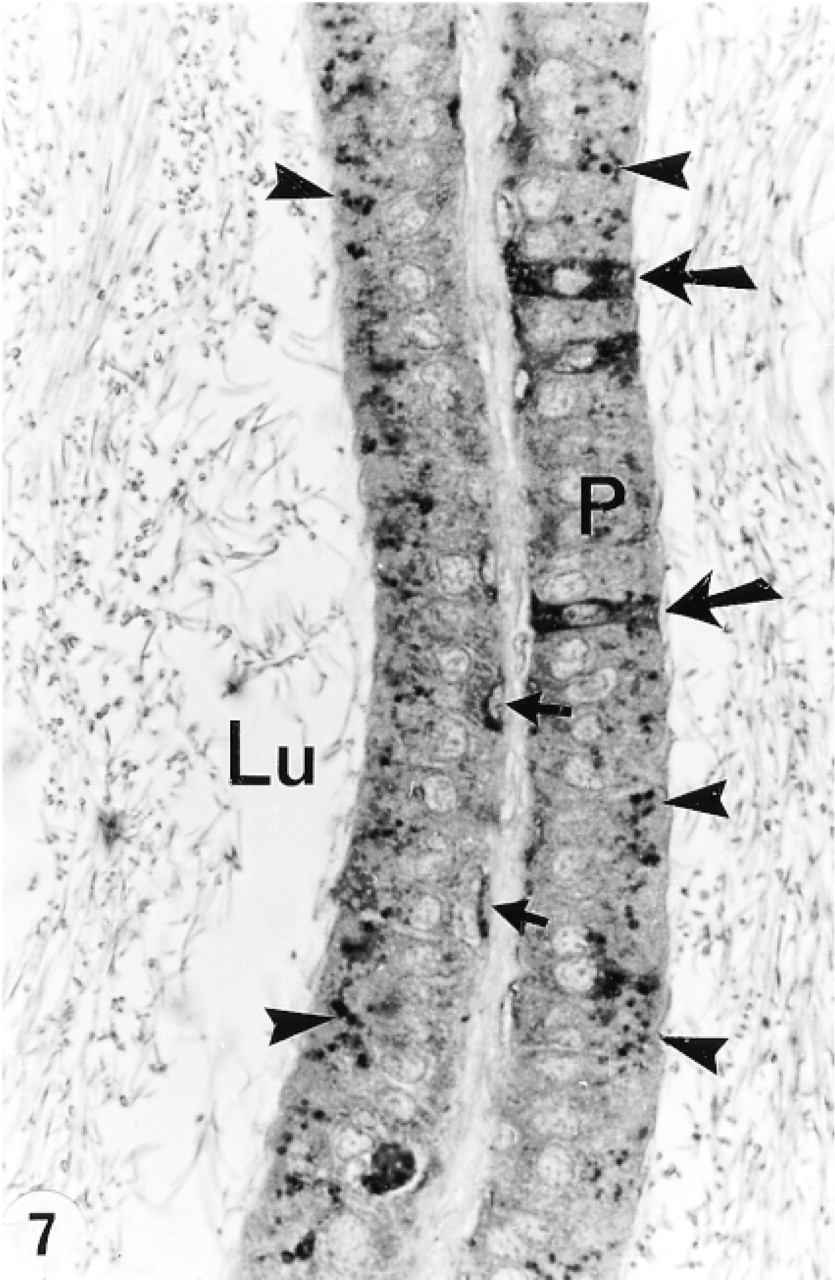
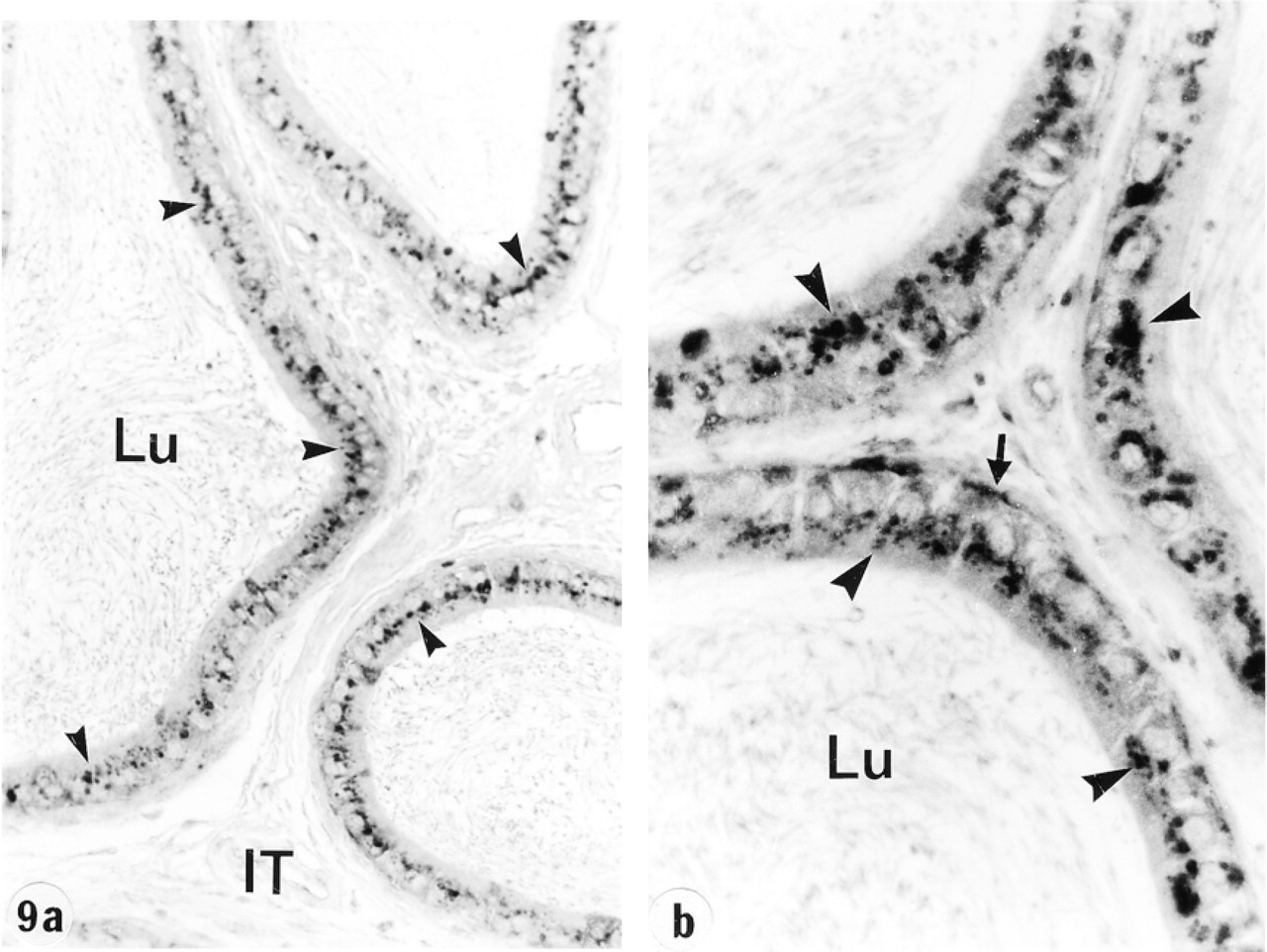
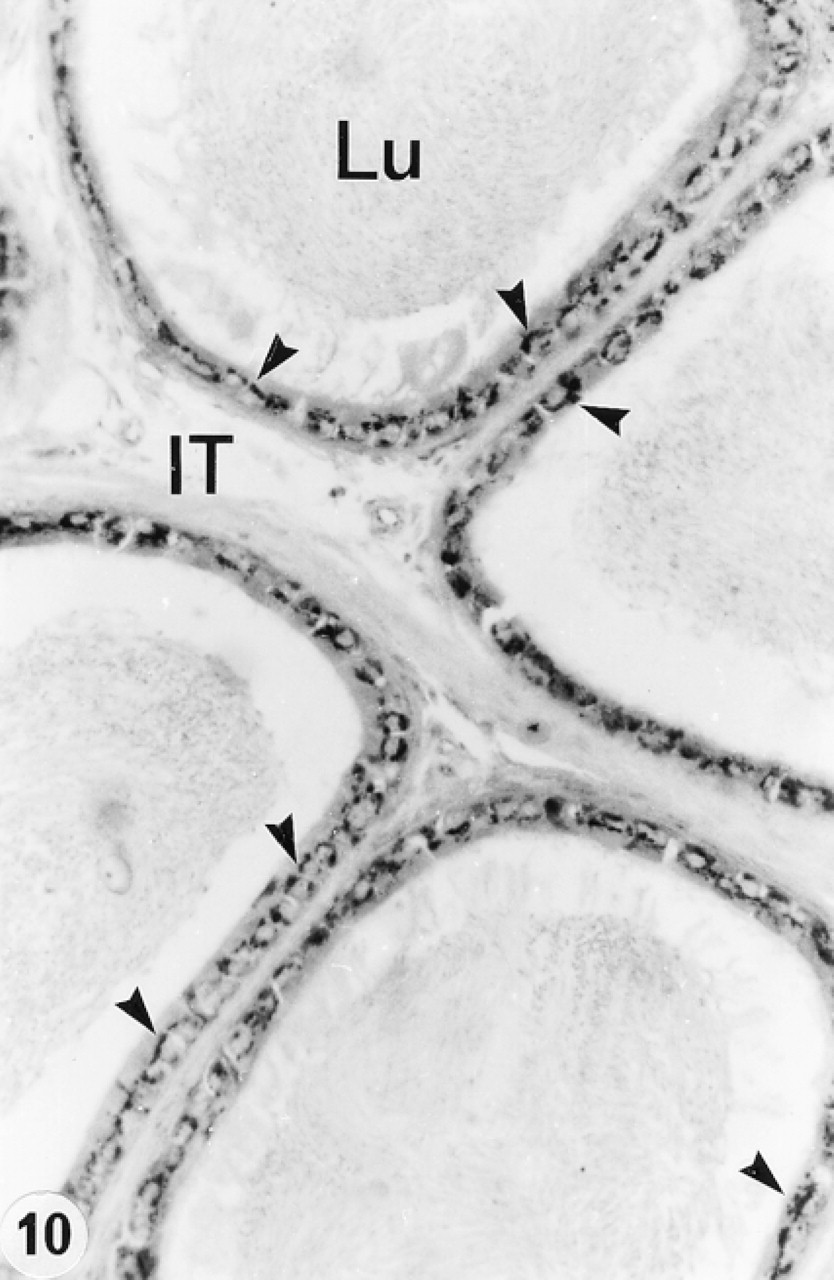
In the efferent ducts, the supranuclear lysosomes of nonciliated cells were intensely reactive (Figures 2a and 2b). In the epididymis, expression of cathepsin A in principal cells was region-specific. Many reactive segment, where they occupied the principal cell's supranuclear region (Figures 3a, 3b, 4, and 5). The intermediate zone, characterized by the presence of giant endosomes located in the apical region of principal cells, revealed immunostaining that differed between its proximal and distal regions (Figures 4 and 6). The former showed many reactive lysosomes that occupied a distinct region over the nucleus of the principal cell (Figure 6), whereas few reactive lysosomes were noted in the distal region (Figure 4). A difference was also noted between the proximal and distal caput regions. In the proximal region, many reactive lysosomes were noted in the majority of principal cells (Figure 7). In the distal region, many lysosomes of some principal cells were intensely reactive, whereas others showed few reactive lysosomes or none at all (Figure 8). Principal cells of the corpus and proximal cauda epididymidis revealed many intensely reactive lysosomes (Figures 9 and 10).
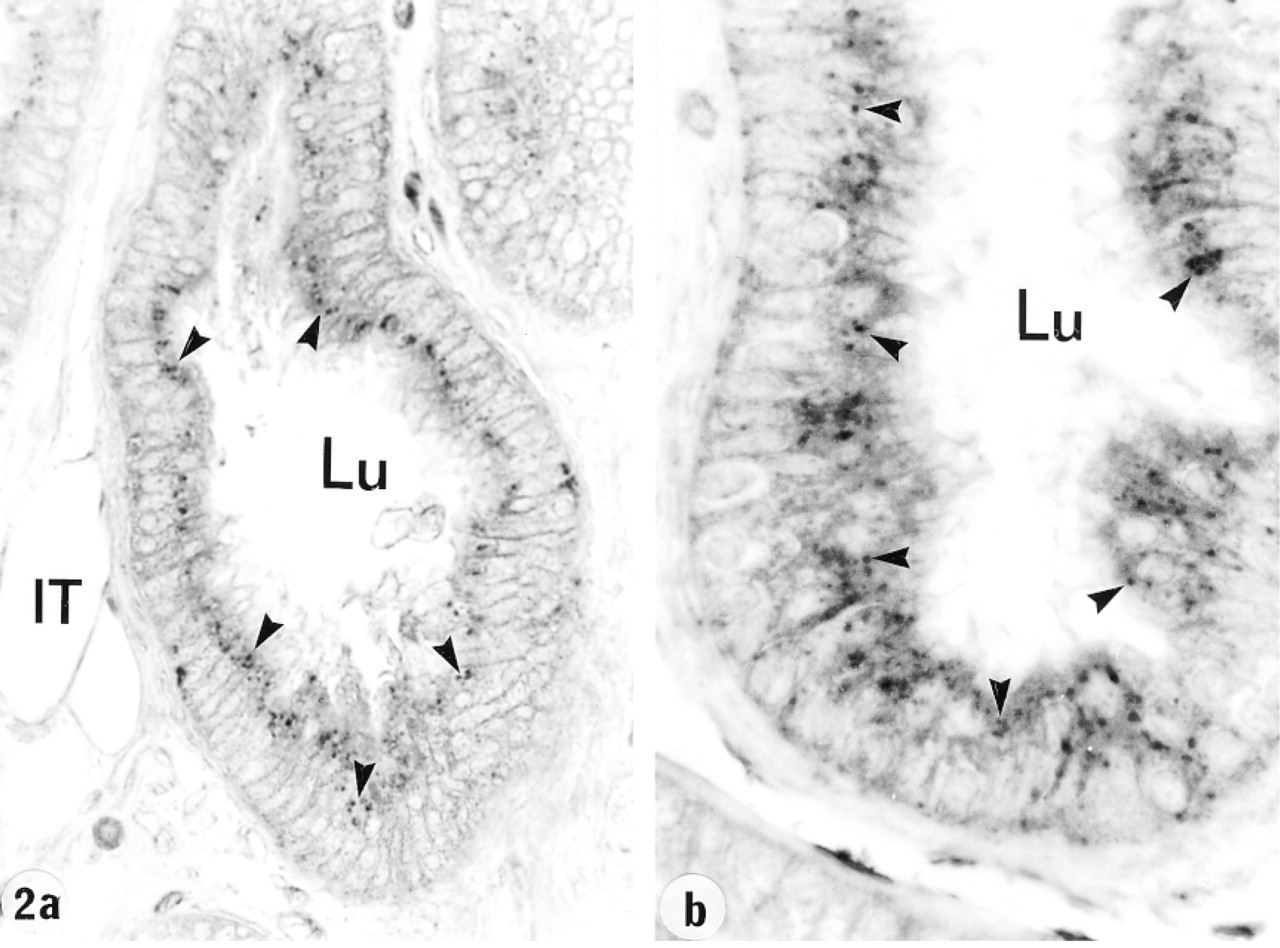
Efferent ducts at low (
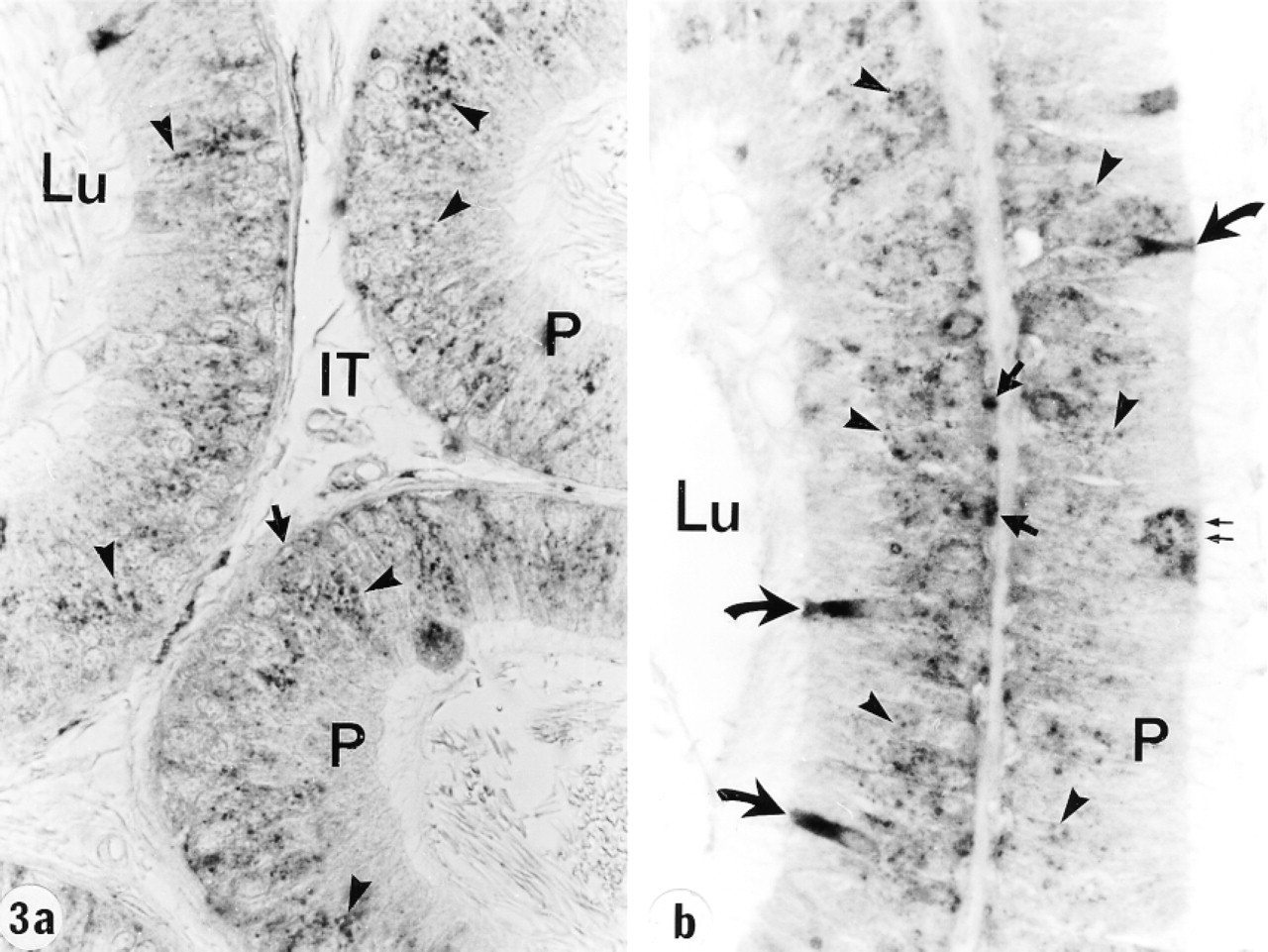
Tubules of the proximal initial segment of the epididymis at low (
Narrow cells of the initial segment and intermediate zone were intensely reactive (Figures 3–6), as were clear cells of the caput, corpus, and cauda regions (Figures 7 and 8). In contrast, basal cells showed a region-specific expression. Many intensely reactive basal cells were noted in the distal initial segment (Figure 5) and proximal intermediate zone (Figure 6), followed by a progressive decrease from the caput to the cauda region (Figures 7, 8, 9a, 9b, and 10).
Electron Microscopy
Gold particles were noted over the lysosomes of those cells that were reactive as seen in the light microscope, examples of which are Sertoli cells of the testis (Figure 11), principal cells of the initial segment (Figure 12) and corpus (Figure 13), epididymidis, and clear cells of the caput region (Figure 14). Not illustrated are nonciliated and Leydig cells. There was no labeling of the coated pits or vesicles, endosomes, and multivesicular bodies (MVBs) of these cells. In general, the pattern of immunogold labeling of lysosomes mimicked that noted for the immunoperoxidase reaction, because corresponding cells and regions that were intensely or weakly reactive in the light microscope revealed many or few gold particles in the electron microscope, respectively, further attesting to the specificity of the reaction because different fixatives and embedding media were utilized in each case.
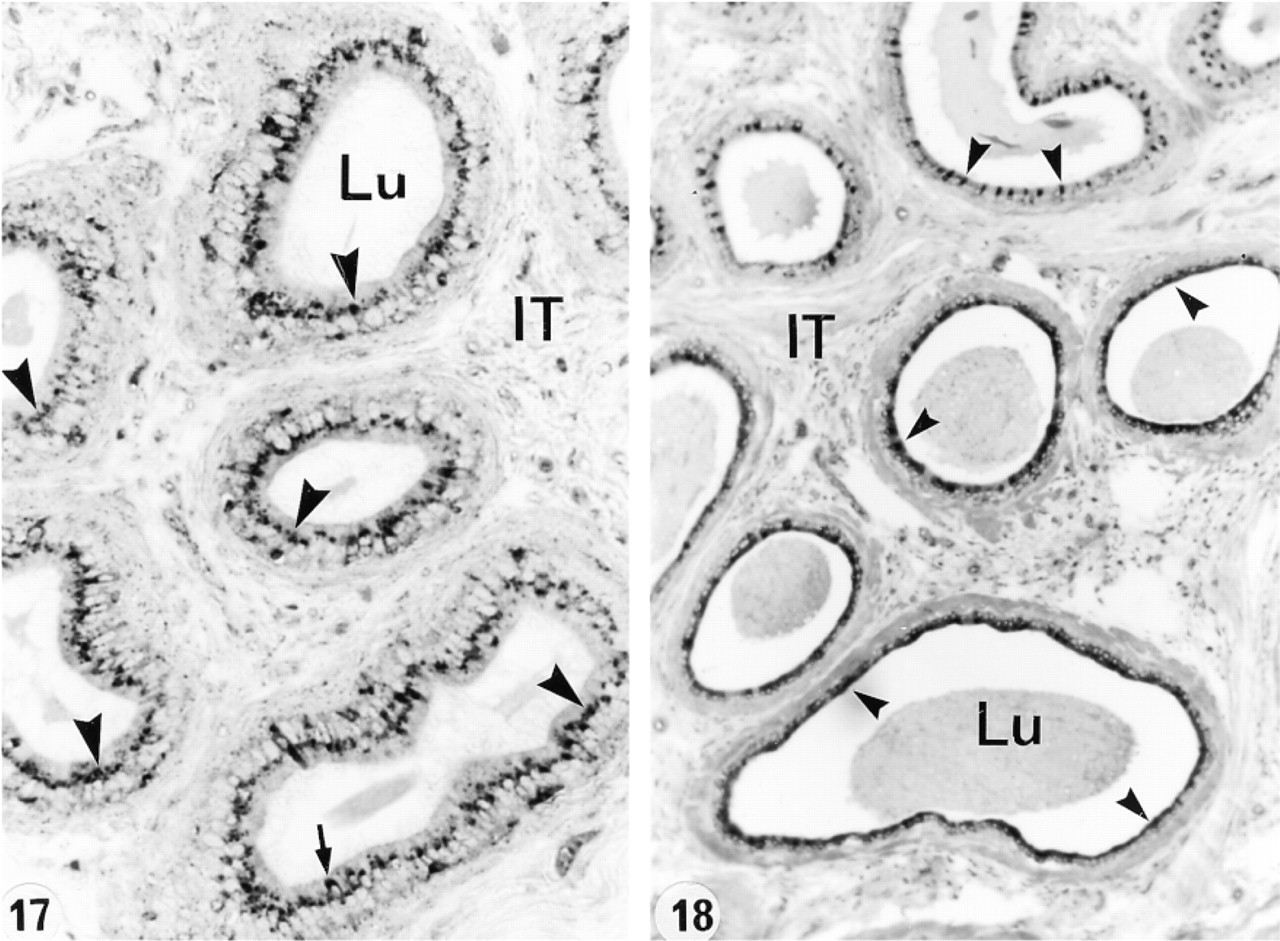
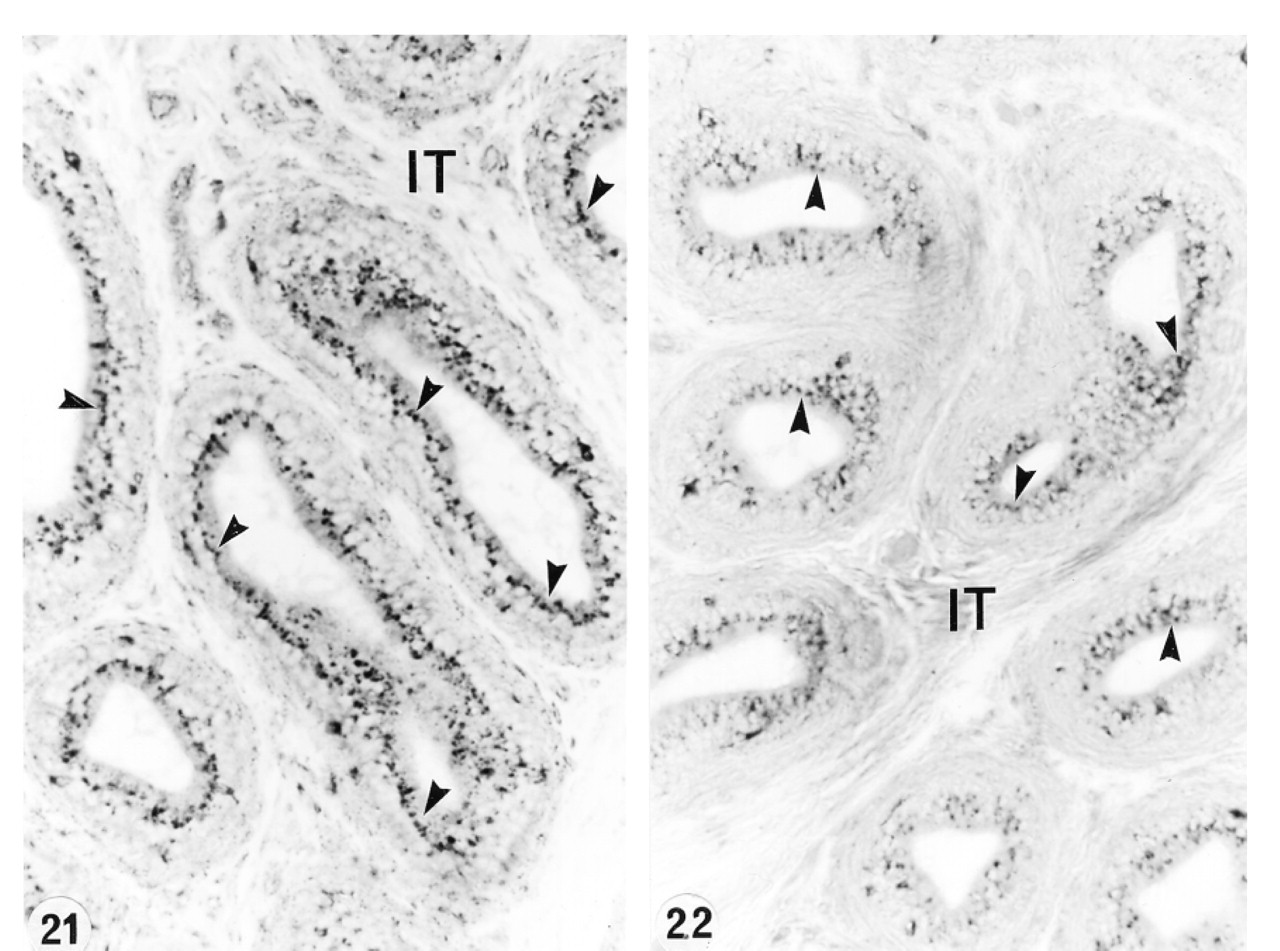
In an attempt to determine whether or not circulating or luminal testicular factors or pituitary factors regulated the expression of cathepsin A in the epididymis, various experimental protocols were performed. After ligation of the efferent ducts for up to 21 days, no difference was noted in the level of expression of cathepsin A in lysosomes of all cell types and in all the different regions (Figures 15 and 16). This was also the case after 21 days of orchidectomy (Figures 17 and 18). Twenty-eight days after hypophysectomy, no difference in cathepsin A expression was noted in Sertoli or Leydig cells of the testis (Figures 19 and 20) or in epithelial cells of the epididymis (Figures 21 and 22).
Use of normal rabbit serum for LM immunostaining showed no staining over cells of the testis or epididymis, and EM immunolabeling revealed only background levels of gold particles over the cells, examples of which can be seen in Veri et al. (1993).
Discussion
In this study, lysosomes of Sertoli cells were reactive for cathepsin A antibody (Table 1), as suggested by Rottier et al. (1998). However, whereas intense expression was seen at most stages of the cycle, there was a notable absence of lysosomal reaction at Stages IX-XII. Other lysosomal enzymes have been localized in lysosomes of Sertoli cells, such as cathepsin D and β-hexosaminidase A (Igdoura et al. 1995; Hall et al. 1996; Hermo et al. 1997), as well as cathepsin L, which was expressed mainly at Stages VI and VII of the cycle (Zabludoff et al. 1990).
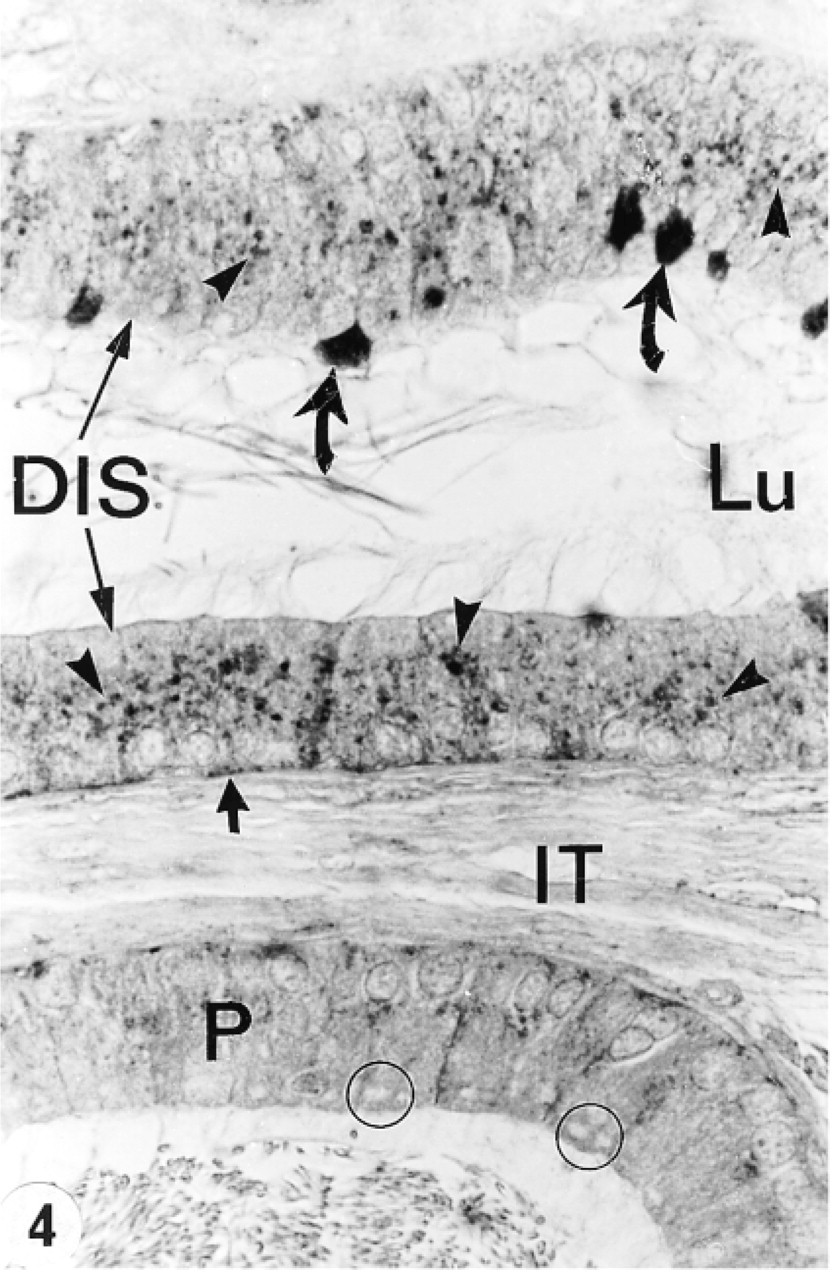
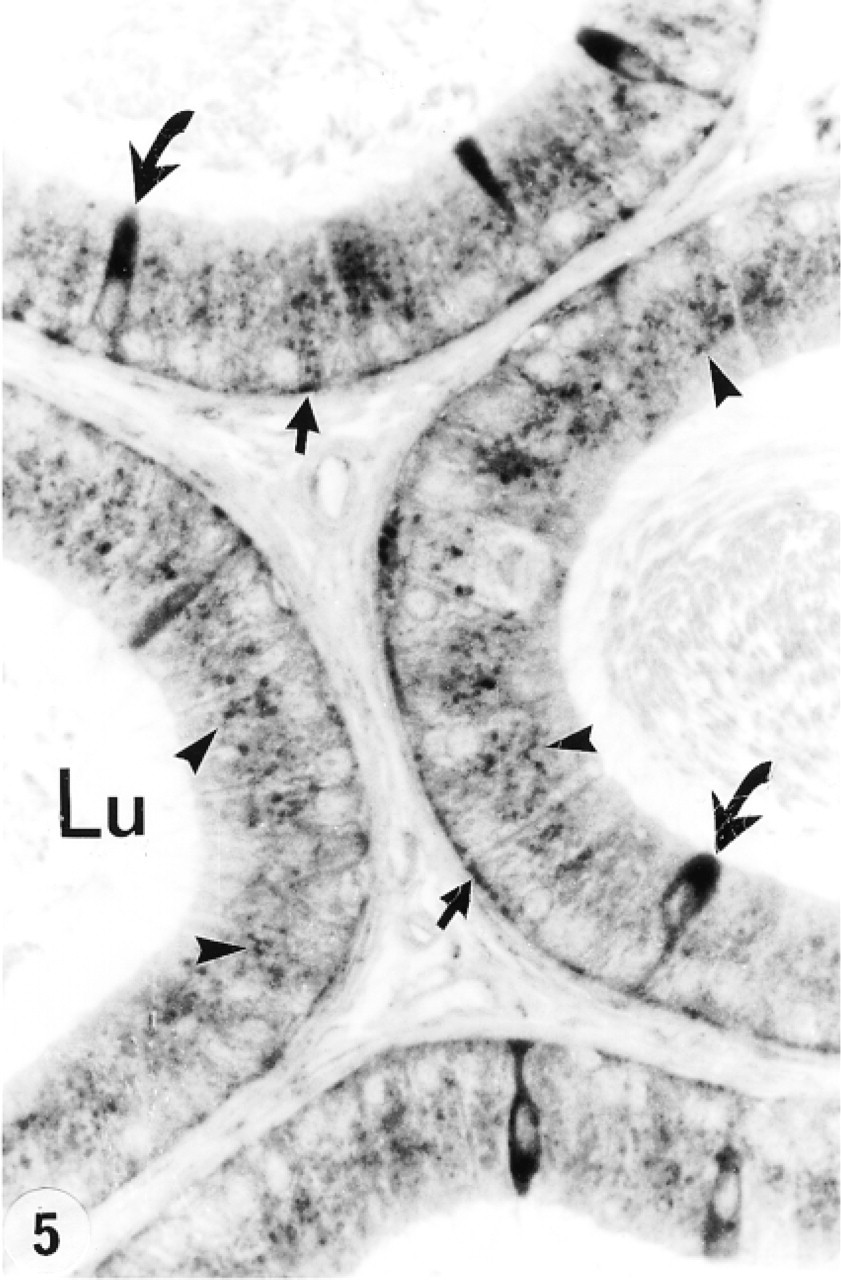
Tubule of the distal initial segment (DIS) and distal intermediate zone (below) of the epididymis immunostained with anti-cathepsin A antibody. Many reactive lysosomes (arrowheads) fill the supranuclear region of principal cells of the DIS. Narrow cells are intensely reactive (curved arrows), as are some basal cells (small arrow). In the distal intermediate zone, few reactive lysosomes are noted in principal cells (P) which, in this region, are characterized by giant apical endosomes (circles). Lu, lumen; IT, intertubular space. Original magnification × 160.
High-power view of the distal initial segment of the epididymis immunostained with anti-cathepsin A antibody. Many reactive lysosomes (arrowheads) are present in the supranuclear region of principal cells. Narrow cells are intensely reactive (curved arrows), as are many basal cells (arrows). Lu, lumen. Original magnification × 128.
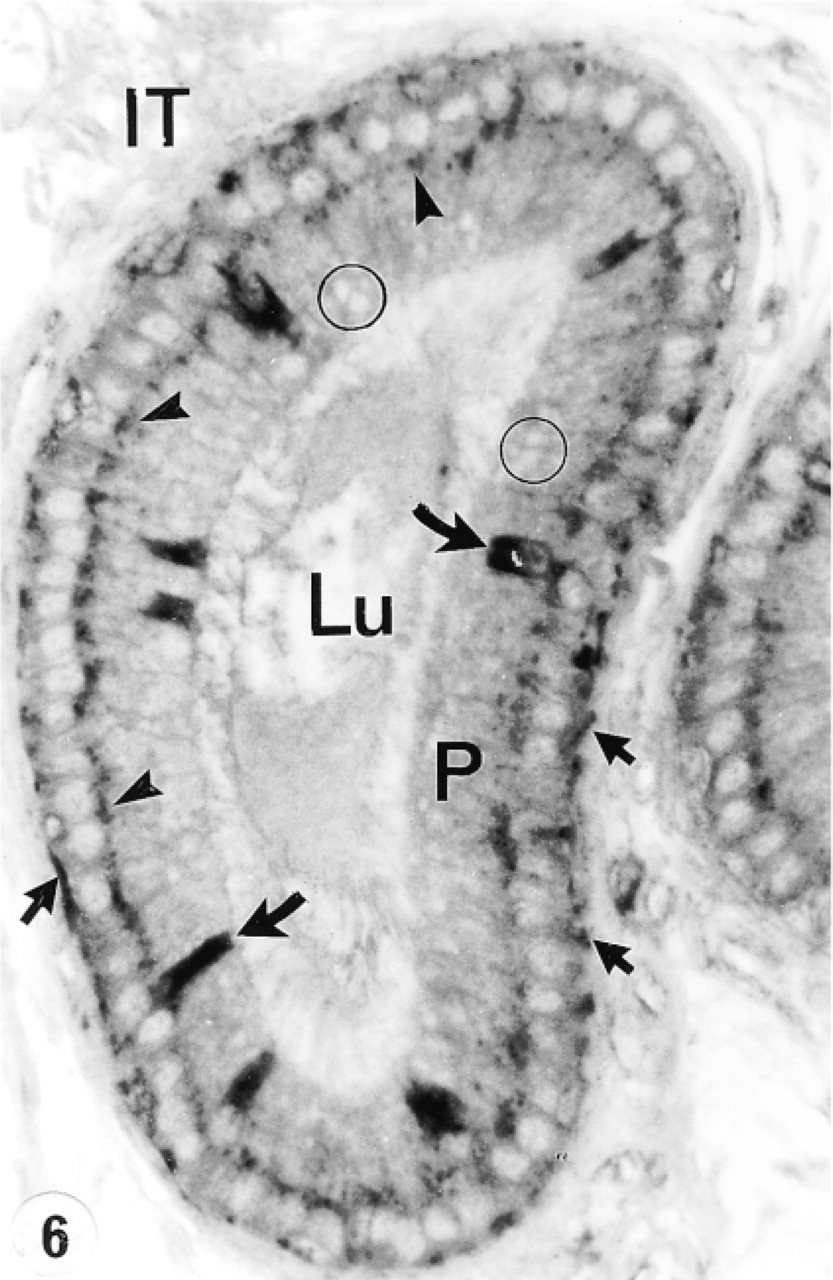
Proximal intermediate zone immunostained with anti-cathepsin A antibody. Principal cells (P) show several intensely reactive supranuclear lysosomes (arrowheads) and unreactive giant endosomes in their apical region (circles). Many basal cells are reactive (arrows), as are narrow cells (curved arrows). Lu, lumen; IT, intertubular space. Original magnification × 160.
Proximal caput epididymidis immunostained with anti-cathepsin A antibody, revealing many reactive lysosomes (arrowheads) in principal cells and some reactive basal cells (small arrows). Clear cells are intensely reactive (large arrows). Lu, lumen. Original magnification × 160.
Sertoli cells of the testis are actively involved in endocytosis, which has been shown to account for the formation of lysosomes, whereby endosomes, formed by fusion of coated vesicles, progressively transform into multivesicular bodies (MVBs) which in turn evolve into lysosomes (Morales et al. 1985; Morales and Clermont 1993). This activity appears to be cyclic because lysosome numbers vary according to the different stages of the cycle of the seminiferous epithelium (Morales et al. 1985; Morales and Clermont 1993; Hermo et al. 1994). Although abundant at most stages, their numbers are significantly reduced at Stages IX-XII, at a time when they are engaged in the degradation of residual bodies emanating from late spermatids at the time of their release into the lumen at Stage VIII of the cycle. The use of tracer studies and acid phosphatase cytochemistry has demonstrated that only the late residual bodies located nearer to the base of the seminiferous epithelium become labeled. These data suggest that the decreased number of Sertoli cell lysosomes during Stages XI-XII is due to their fusion with the late residual bodies near the base of the epithelium (Morales et al. 1985; Morales and Clermont 1993). In the present study, late but not early residual bodies were intensely reactive for anti-cathepsin A antibody, suggesting that lysosomes of Sertoli cells fused with the late residual bodies at Stages IX-XII of the cycle to degrade their contents and demonstrating a role for cathepsin A in this process. Similar to cathepsin A, β-hexosaminidase A was localized to late residual bodies, suggesting a role for this enzyme as well in their degradation (Hermo et al. 1997).
The absence of cathepsin A expression in lysosomes of germ cells (Table 1) is not surprising because several other lysosomal enzymes, such as β-hexosaminidase A and cathepsin B, were not noted in germ cells (Igdoura et al. 1995; Hermo et al. 1997). Cathepsin D and β-hexosaminidase A have been localized in spermatids, but the reaction was noted over the acrosomes of these cells (Srivastava and Ninjoor 1982; Igdoura et al. 1995; Hall et al. 1996).
Lysosomal enzymes have been localized within lysosomes of Leydig cells (Hermo et al. 1992a, 1997; Igdoura et al. 1995). To this list, cathepsin A can now be added, as also noted by others (Rottier et al. 1998; Sohma et al. 1999). These cells have been documented to be active endocytic cells (Hermo et al. 1985a; Hermo and Lalli 1988). The presence of cathepsin A in Leydig cell lysosomes suggests a role for this enzyme in the breakdown of substances derived from endocytosis, and in autophagy, which has been demonstrated in these cells (Tang et al. 1988).
The nonciliated cells of the efferent ducts express cathepsin A (Table 1), and other enzymes (Hermo et al. 1992a, 1997; Igdoura et al. 1995). As with cathepsin D and β-hexosaminidase A, cathepsin A is endogeneously expressed and is not internalized by endocytosis, as confirmed in the present study by electron microscopy which revealed labeling of their lysosomes but not of endosomes or MVBs. This is unlike the case for SGP-1 which not only is endogeneously expressed but is also internalized from the lumen by nonciliated cells as the Sertoli-derived form of SGP-1 (Hermo et al. 1992a, 1995; Igdoura et al. 1993). The presence of cathepsin A and other lysosomal enzymes in nonciliated cells suggests an active role for their lysosomes in the degradation of substances endocytosed from the lumen, a function that has been well-documented for these cells (Hermo and Morales 1984; Hermo et al. 1994; Ilio and Hess 1994). In the present study, ciliated cells although involved in endocytosis (Hermo et al. 1985b) were found to be unreactive for cathepsin A.
Epididymis
In the present study, the lysosomes of principal cells showed a region-specific distribution for cathepsin A along the length of the epididymis with most regions showing many reactive lysosomes except for the intermediate zone (Table 2; Figure 23). Furthermore, in the distal caput region, whereas some principal cells showed many reactive lysosomes, others showed few or none, representing a checkerboard-like staining pattern also noted for expression of various other synthesized proteins (Hermo et al. 1991; Rankin et al. 1992; Veri et al. 1993) (Table 2; Figure 23). In a previous study, we noted that cathepsin A was highly expressed in lysosomes of principal cells of the middle and distal cauda regions as well as along the entire vas deferens which, together with the present data, indicates an important role for this enzyme in their lysosomes (Andonian and Hermo 1999).
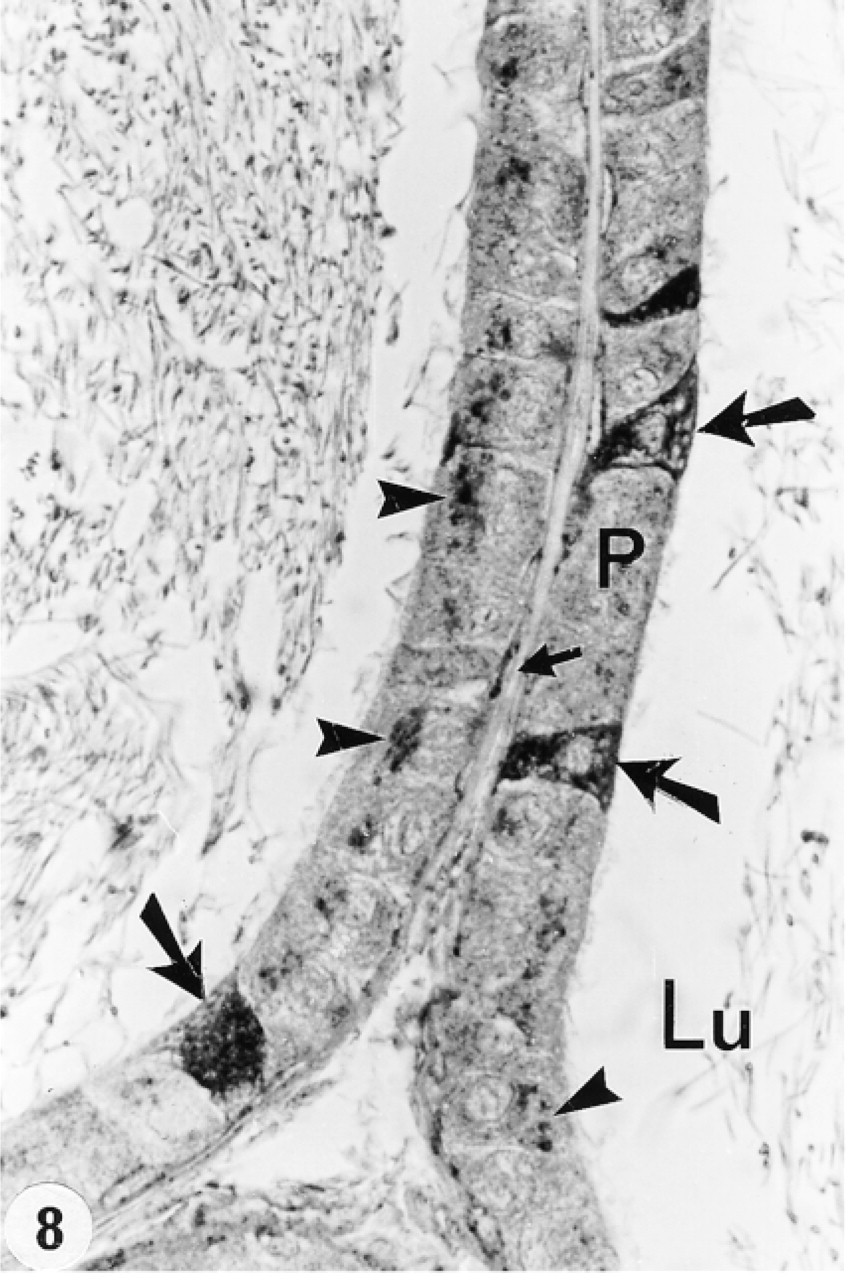
Distal caput epididymidis immunostained with anti-cathepsin A antibody. Some principal cells (P) show many reactive lysosomes (arrowheads), whereas others reveal few or none. Some basal cells are reactive (small arrows), and clear cells are intensely reactive (large arrows). Lu, lumen. Original magnification × 128.
Corpus epididymidis at low (
Cauda epididymidis immunostained with anti-cathepsin A antibody reveals many reactive lysosomes in principal cells (arrowheads). Lu, lumen; IT, intertubular space. Original magnification × 128.
Cathepsin A expression in normal adult testis and efferent ducts a

+, immunostaining present;−, immunostaining absent.
Clear cells are noted for their role in the endocytosis of various substances from the epididymal lumen (Moore and Bedford 1979; Hermo et al. 1988b). These cells found in the caput, corpus, and cauda regions are filled with supranuclear lysosomes and express many lysosomal enzymes in all regions, as is the case in the present study for cathepsin A (Table 2; Figure 23) (Hermo et al. 1992a, 1997; Igdoura et al. 1995). Clear cells of the middle and distal cauda epididymidis and proximal vas deferens were also highly reactive (Andonian and Hermo 1999), suggesting an important role for cathepsin A in clear cells. In contrast, expression of cathepsins D and H and of β-d-glucuronidase was noted to be region-specific, whereas cathepsin B was absent from clear cells (Tomomasa et al. 1994; Igdoura et al. 1995; Abou-Haila et al. 1996).
Narrow cells are involved in the acidification of the epididymal lumen of the initial segment and intermediate zone (Brown et al. 1992; Adamali and Hermo 1996), and their lysosomes express different lysosomal enzymes (Hermo et al. 1992a, 1997; Igdoura et al. 1995; Abou-Haila et al. 1996). The present study noted intense expression of cathepsin A in narrow cells (Table 2; Figure 23). Although these cells have been shown to be involved in endocytosis of fluid-phase tracers (Hermo et al. in press), the specific proteins internalized and acted on by their lysosomes are as yet unknown.
Basal cells are present along the entire length of the epididymis, where they form an incomplete mesh-like barrier at the periphery of the tubule, and by expressing various GSTs may protect sperm from harmful electrophils (Veri et al. 1994; Papp et al. 1995). Basal cells contain lysosomes, with expression of different lysosomal enzymes varying from region to region (Hermo et al. 1992a, 1994, 1997; Igdoura et al. 1995; Abou-Haila et al. 1996). In the present study, cathepsin A was also expressed in a region-specific manner from the initial segment to the proximal cauda region (Table 2; Figure 23), in contrast to the middle and distal cauda epididymidis and entire vas deferens, where basal cells were intensely reactive for cathepsin A and showed no region specificity (Andonian and Hermo 1999). Specific proteins that are endocytosed by basal cells and degraded by their lysosomes are as yet to be identified.
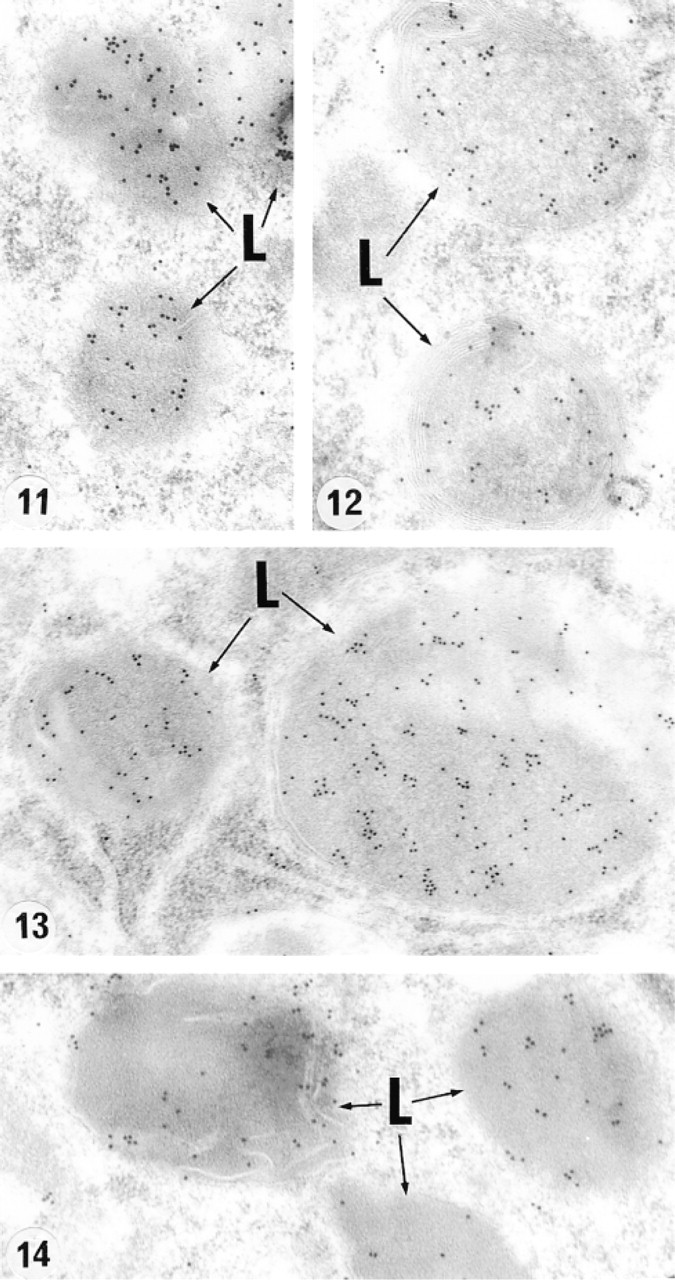
Electron micrographs of lysosomes (L) immunolabeled with anti-cathepsin A antibody of a Sertoli cell (Figure 11), principal cells of the initial segment (Figure 12), and corpus (Figure 13) epididymidis and a clear cell of the caput epididymidis (Figure 14). Note many gold particles located within the matrix of these structures. Original magnifications:
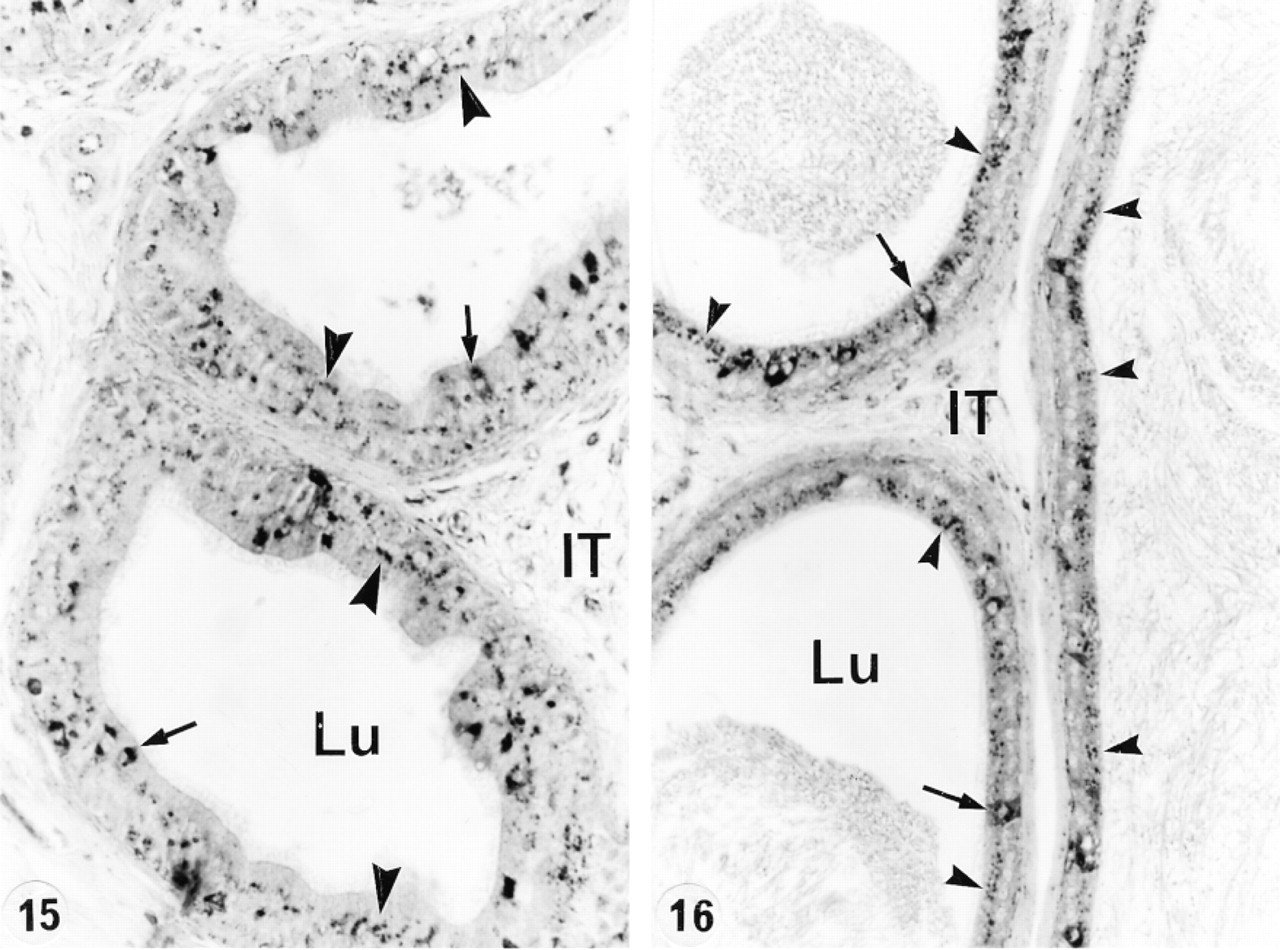
Corpus (Figure 15) and cauda (Figure 16) epididymidis of a 21-day efferent duct-ligated animal immunostained with anti-cathepsin A antibody. Many intensely reactive (arrows) lysosomes are present in principal cells (arrowheads) and clear cells. Lu, lumen; IT, intertubular space. Original magnifications:
In the present study, the cathepsin A-reactive structures in nonciliated, principal, and clear cells of normal animals were confirmed to be lysosomes rather than endosomes or MVBs, on the basis of similarities in their shape, position, and appearance to similar structures confirmed to be lysosomes from earlier studies employing morphology, tracer studies, and acid phosphatase cytochemistry (Hermo and Morales, 1984; Robaire and Hermo 1988; Hermo et al. 1988a,b, 1994). Although it is still unclear how cathepsin A arrives at lysosomes, it does not appear to be endocytosed from the lumen because endosomes and MVBs, early endocytic structures involved in the uptake and recycling of transferrin (Djakiew et al. 1984) and differing in their appearance and position in the cell from lysosomes, were unlabeled. It can be suggested, as from other studies on lysosomal enzymes, that cathepsin A is targeted to lysosomes via the mannose-6-phosphate receptor in the form of small constitutive vesicles derived from the trans-Golgi network of the Golgi apparatus (Griffiths and Simons 1986; Zhou et al. 1996; Farquhar and Hauri 1997).
In nonciliated and various epididymal epithelial cells (Hermo and Morales 1984; Hermo et al. 1988a, 1994) as well as many other cell types, endosomes form by the fusion of early small endocytic vesicles and gradually transform into MVBs, which then evolve into lysosomes by a process described as the maturation model (Murphy 1991; Dunn and Maxfield 1992). It therefore appears that lysosomes would be a good indicator of what the cells are endocytosing from the epididymal lumen, and the absence or diminished expression of a given lysosomal enzyme may reflect the type of substances that are being internalized in any given region of the epididymis. However, a correlation between the types of substances internalized by the epididymis and lysosomal enymes involved has not as yet been performed.
Region-specificity has also been noted for other lysosomal enzymes in the case of nonciliated, principal, clear, and narrow cells (Hermo et al. 1992a, 1997; Igdoura et al. 1995; Abou-Haila et al. 1996; Adamali and Hermo 1996). However, in each case, expression was notably different. Clearly, the variable distribution of lysosomal enzymes in lysosomes of these cells reflects a complex and changing luminal environment whereby these cells internalize substances derived from the lumen as sperm mature and are stored in the epididymis. These substances include the breakdown products of degenerating sperm, cytoplasmic droplets, and specific proteins that are endocytosed in a region-specific manner (Cooper and Hamilton 1977; Lea et al. 1978; Hermo et al. 1988b, 1992b; Flickinger et al. 1988; Rankin et al. 1992; Vierula et al. 1995). The region-specific localization of all these lysosomal enzymes indicates a heterogeneous nature to the functions of these cells, which is consistent with region-specific expression of secretory proteins, GSTs, and other substances, as well as notable differences in the structural appearance of their lysosomes and integral membrane proteins (Robaire and Hermo 1988; Suarez–Quian et al. 1992; Hermo et al. 1994; Papp et al. 1995; Orgebin–Crist 1996).
In the present study, no noticeable difference was observed in the expression of cathepsin A in the different cell types of the epididymis after efferent duct ligation, hypophysectomy, or orchidectomy, with or without testosterone supplementation. These data indicate that cathepsin A is not regulated in these cell types by testicular or pituitary factors. This is in direct contrast to many proteins synthesized by the epididymal epithelial cells that have been shown to be regulated by luminal or circulating testicular factors as well as by pituitary factors (Cornwall and Hann 1995; Robaire and Viger 1995; Papp and Hermo 1996; Orgebin–Crist 1996; Kirchhoff 1999). In addition, our findings show differences in regulation of cathepsin A from other lysosomal enzymes described by those who noted that steroid hormones, such as androgens and estrogens, regulate the expression of several lysosomal enzymes (Gupta and Setty 1995; Abou-Haila et al. 1996). In the efferent ducts, expression of SGP-1 by nonciliated cells was regulated by pituitary factors (Rosenthal et al. 1995), outlining the dramatic differences in regulation of different lysosomal enzymes in the different cell types. Hypophysectomy also had no noticeable effect on the expression of cathepsin A expression in Sertoli or Leydig cells, suggesting that pituitary factors did not regulate its expression in these cell types despite the fact that hormones play an important role in maintaining the structural appearance of Sertoli cells and their synthesis of various proteins (Christensen 1975; Bardin et al. 1988).
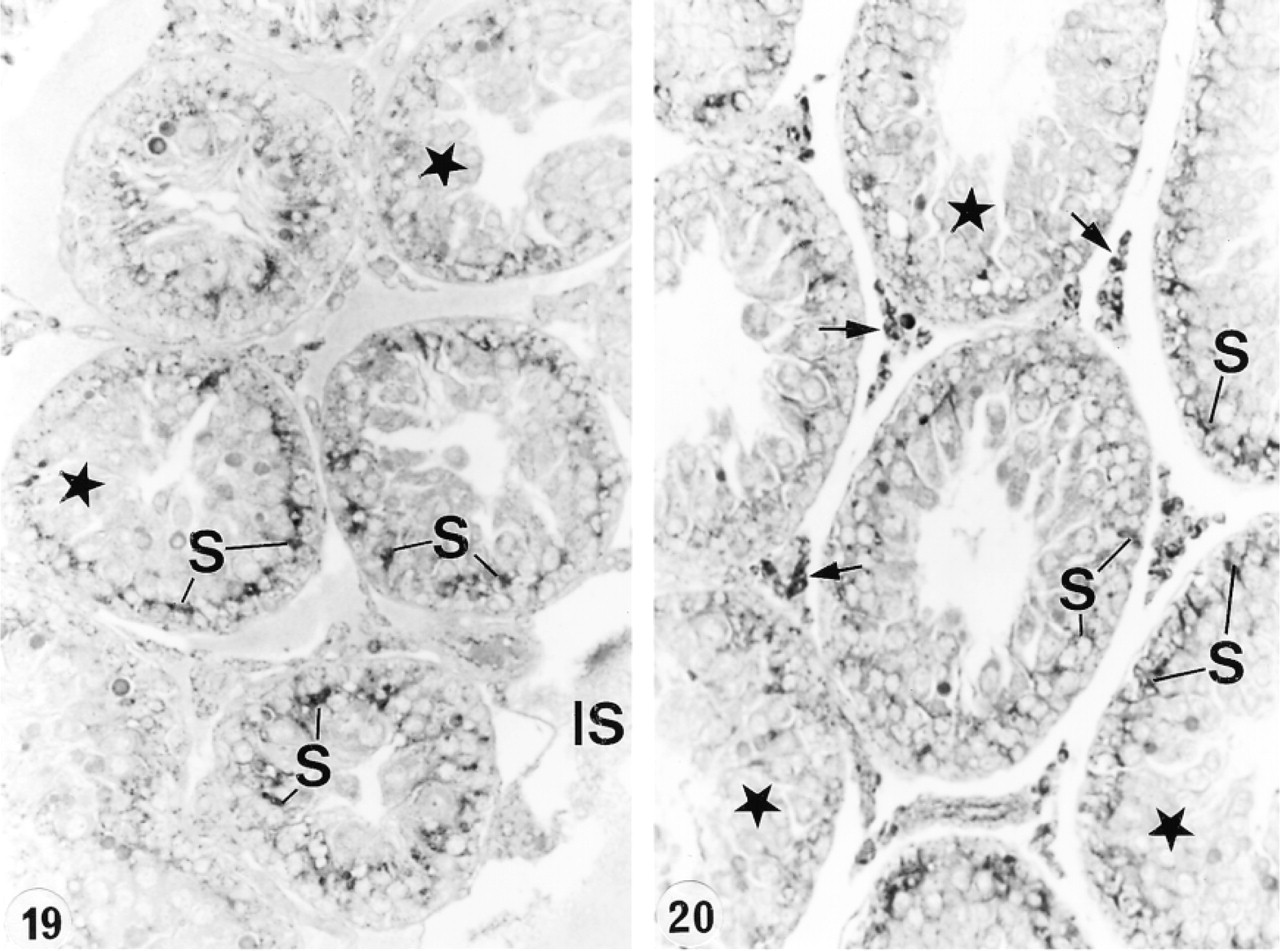
Seminiferous tubules of the testis 28 days after hypophysectomy and immunostained with anti-cathepsin A antibody. A distinct reaction appears over the basal region of the epithelium in association with Sertoli cells (S) and over interstitial cells (arrows). Germ cells do not appear to be stained (stars). IS, interstitial space. Original magnification × 80.
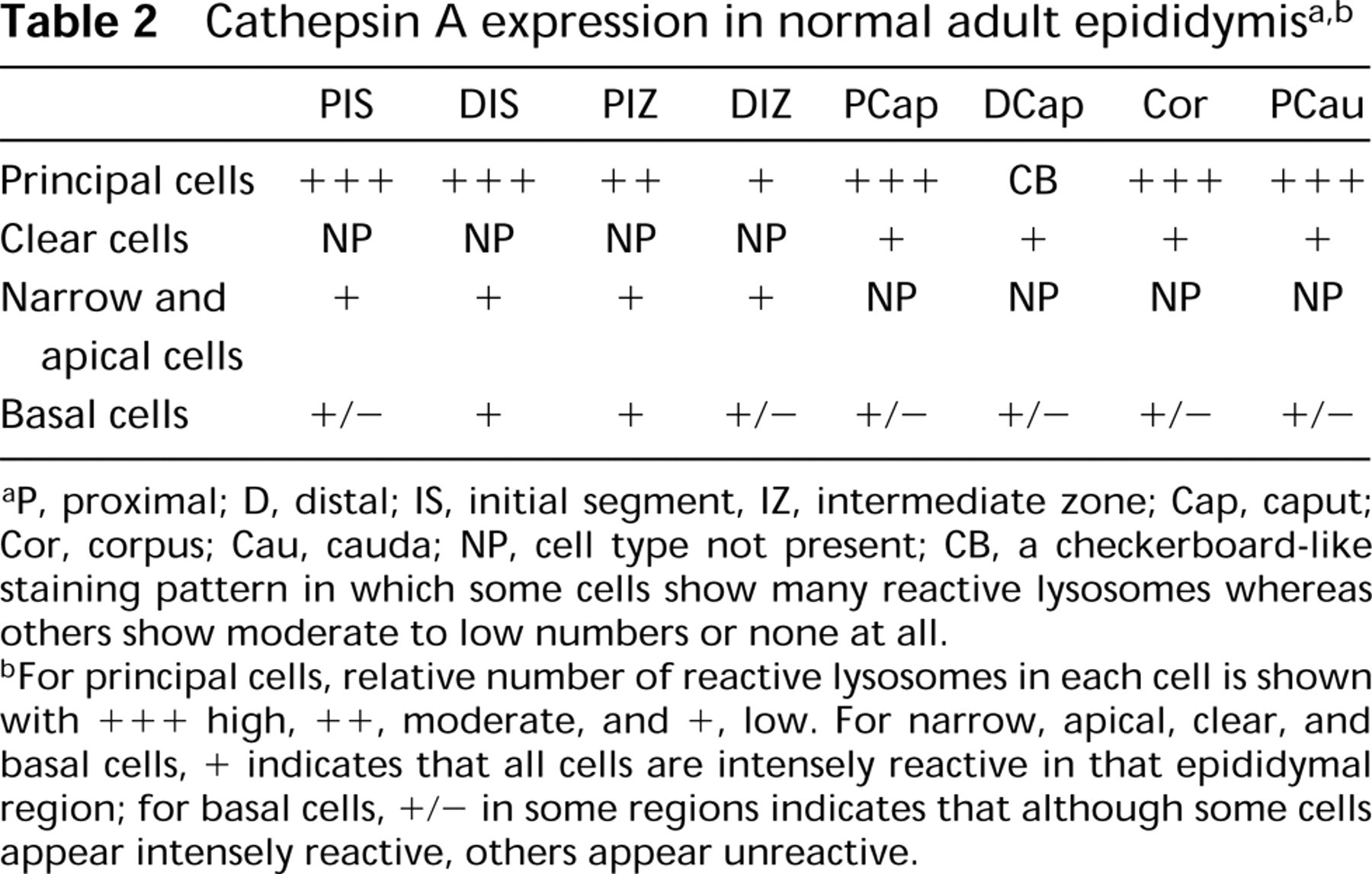
P, proximal; D, distal; IS, initial segment, IZ, intermediate zone; Cap, caput; Cor, corpus; Cau, cauda; NP, cell type not present; CB, a checkerboard-like staining pattern in which some cells show many reactive lysosomes whereas others show moderate to low numbers or none at all.
For principal cells, relative number of reactive lysosomes in each cell is shown with +++ high, ++, moderate, and +, low. For narrow, apical, clear, and basal cells, + indicates that all cells are intensely reactive in that epididymal region; for basal cells, +/− in some regions indicates that although some cells appear intensely reactive, others appear unreactive.
In the present study, electron microscopic analysis on experimental animals was not performed because we assumed that cathepsin A was still localized to lysosomes, as evidenced from the light microscopic analysis of a reaction over punctate supranuclear structures corresponding to the position and shape of lysosomes in cells of control animals.
Genetic lesions in the cathepsin A gene cause severe loss of neuraminidase activity and render β-galactosidase susceptible to rapid intralysosomal proteolysis, resulting in a combined enzyme deficiency that is the basis of the lysosomal storage disorder galactosialidosis. The latter is characterized by the lysosomal storage of sialylated oligosaccharides and glycopeptides in affected tissues (van Pelt et al. 1988a,b,c). In cathepsin A knockout mice, extensive vacuolation of the cytoplasm as seen by LM, possibly corresponding to lysosome accumulation, was reported to occur in all high-expressing cells of the epididymis and in interstitial cells of the testis, but region- or cell type-specific differences in the epididymis as well as EM analysis were not examined (Rottier et al. 1998). The absence of β-hexosaminidase A in mice also showed marked effects on the number, size, and distribution of lysosomes in cells that were affected, indicating that lysosomal storage disorders contribute a great deal to our understanding not only of the contents of lysosomes and role of their individual enzymes but also of the functions of the affected cells (Trasler et al. 1998; Adamali et al. 1999a,b).
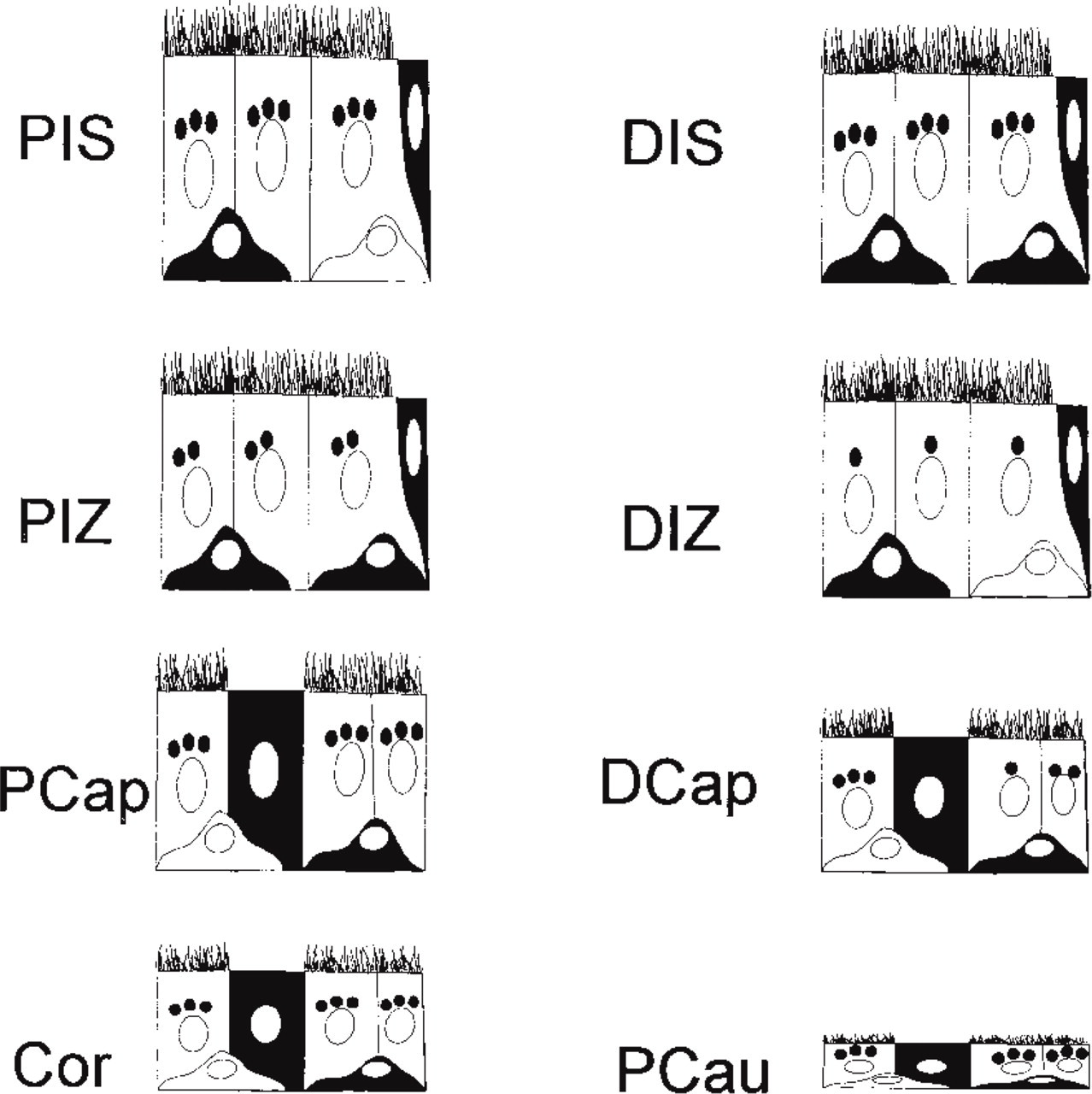
Schematic representation of the staining pattern for cathepsin A in the proximal (P) and distal (D) regions of the initial segment (IS), intermediate zone (IZ), caput (Cap), corpus (Cor), and cauda (Cau) regions of the normal adult epididymis. Principal cells are represented as columnar cells with microvilli and are seen in all epididymal regions. Clear cells, present only in the caput, corpus, and cauda epididymal regions, are represented as columnar cells without microvilli. The hemispherical basal cells are noted in the basal region of the epithelium in all epididymal regions. Narrow cells, found only in the initial segment and intermediate zone, are represented as goblet-shaped cells and are situated in the schematic at the right edge of the epithelial section. The number of reactive lysosomes in principal cells is designated qualitatively as dark dots with three representing high, two moderate and one low numbers. For narrow, clear, and basal cells, the intensity of the reaction is recorded, with dark staining representing an intense reaction over the cell and no staining as the absence of a reaction. The cytoplasmic reaction in these cell types is due to the fact that these cells contain many lysosomes, resulting in uniform staining over this region of the cell. The number of reactive lysosomes in principal cells is high in the PIS, DIS, PCap, Cor, and Pcau, moderate in the PIZ, and low in the DIZ. In the DCap a checkerboard-like staining pattern is noted with cells showing high, moderate, or low numbers of reactive lysosomes. Narrow cells are intensely stained in all regions, as are clear cells. The majority of basal cells are intensely stained in the DIS and PIZ, whereas in all other epididymal regions both intensely reactive and unreactive basal cells are present.
In summary, cathepsin A was localized to Sertoli and Leydig cells but not to germ cells in the testis and to epididymal epithelial cells in a cell- and region-specific manner. In addition, cathepsin A expression does not appear to be regulated by testicular or pituitary factors.
Footnotes
Acknowledgments
Supported by the Medical Research Council of Canada.
We thank Dr Y. Suzuki for the cathepsin A antibody, which he generously provided to us. We thank Jeannie Mui, Mathilda Cheung, P.B. Mukopadhyay, and Jodi Fox for expert technical assistance.