
Abstract
Histamine plays important roles in gastric acid secretion, inflammation, and allergic response. Histamine N-methyltransferase (HMT; EC 2.1.1.8) is crucial to the inactivation of histamine in tissues. In this study we investigated the immunohistochemical localization of this enzyme in guinea pig tissues using a rabbit polyclonal antibody against bovine HMT. The specificity of the antibody for guinea pig HMT was confirmed by Western blotting and the lack of any staining using antiserum preabsorbed with purified HMT. There was strong HMT-like immunoreactivity (HMT-LI) in the epithelial cells in the gastrointestinal tract, especially in the gastric body, duodenum, and jejunum. The columnar epithelium in the gallbladder was also strongly positive. Almost all the myenteric plexus from the stomach to the colon was stained whereas the submucous plexus was not. Other strongly immunoreactive cells included the ciliated cells in the trachea and the transitional epithelium of the bladder. Intermediately immunoreactive cells included islets of Langerhans, epidermal cells of the skin, alveolar cells in the lung, urinary tubules in the kidney, and epithelium of semiferous tubules. HMT-LI was present in specific structures in the guinea pig tissues. The widespread distribution of HMT-LI suggests that histamine has several roles in different tissues.
Keywords
H
Histamine plays an important role in gastric acid secretion (Sanders and Soll 1986), and the therapeutic usefulness of H2 receptor antagonists for treatment of peptic ulcer has been established. Enterochromaffin-like cells present in the oxyntic mucosa and mucosal mast cells have been considered to be major sources of histamine. The histamine released from such stores in turn stimulates H2 receptors present on the parietal cells and activates adenylate cyclase, leading to the cAMP-dependent signal transduction cascade. Histamine plays a permissive role in the secretion of acid from parietal cells in the stomach; the secretion induced by gastrin and muscarinic receptor activation is potentiated by histamine (Sanders and Soll 1986). H2 receptor antagonists efficiently inhibited the acid secretion stimulated through plural receptors. Therefore, histamine is a crucial factor in the oxyntic mucosa. However, the mechanism for its inactivation remained to be determined. Loiselle and Wollin (1993) reported that the inhibition of HMT by a specific inhibitor reduced the uptake of histamine in the rabbit gastric mucosa in vitro and suggested that the uptake of histamine was coupled with its methylation. We found that systemic administration of the HMT inhibitor metoprine increased extracellular levels of histamine in the rat hypothalamus, as measured by an in vivo microdialysis technique (Itoh et al. 1991). Therefore, the activity of HMT may influence the concentrations of extracellular histamine in the tissues. However, little is known about the cellular localization of HMT in tissues.
In the guinea pig trachea, the contractile response to histamine in vitro was enhanced by the addition of an HMT inhibitor to the organ bath, shifting the histamine concentration–effect curve to the left (Ohrui et al. 1992). In mycoplasma-infected hamster (Tamaoki et al. 1998) and nitrogen dioxide-exposed guinea pig (Hoshi et al. 1996), a reduction in HMT activity and hypersensitivity of the trachea to histamine were reported. These findings, again, strongly suggested that HMT activity in the airway is important for inactivation of histamine.
In a previous study, we purified HMT from bovine brain (Nishibori et al. 1991). Recently, we raised a specific antibody against bovine HMT and demonstrated the localization of HMT in the bovine CNS (Nishibori et al. 2000). The antibody crossreacted with HMT from different mammals, including guinea pig. In the present study we investigated the immunohistochemical localization of HMT in guinea pig tissues to better understand the inactivation of histamine in tissues.
Materials and Methods
Animals
We followed the guidelines on animal experimentation of Okayama University Medical School. All procedures were approved by the institutional animal experimentation review committee. Male Hartley guinea pigs weighing 250 g (Shimizu Experimental Animals; Kyoto, Japan) were anesthetized with an IP injection of pentobarbital sodium at a dose of 50 mg/kg and perfused through the left ventricle with 50 ml of ice-cold saline, followed by 500 ml of 10% formalin in 0.01 M PBS. The fixed tissues were dehydrated in a series of ethanol solutions, embedded in paraffin, and cut into serial sections 5–7-μm thick. After the sections had been blocked with 5% normal goat serum and 2% BSA, rabbit anti-HMT antiserum (diluted 1:1500) or preimmune rabbit serum (1:1500) was applied to them for one night at 4C. The sections were rinsed three times with PBS, incubated with biotinylated anti-rabbit IgG goat serum (Vector; Burlingame, CA), washed, and soaked in streptoavidin–peroxidase (Sigma; St Louis, MO). Diaminobenzidine and hematoxylin were the substrate for the enzyme reaction and counterstain, respectively. The preimmune serum did not stain any structure.
Western immunoblotting using a supernatant of the guinea pig tissues showed that anti-bovine HMT antiserum recognized a single band (Figure 1). The apparent size of the band was the same as that of purified guinea pig HMT (Figure 1). This indicates that the antibody is crossreactive for guinea pig HMT.
Production of Antiserum
Bovine brain HMT was purified as described previously (Nishibori et al. 1991). The purified bovine HMT (200 μg) in PBS was emulsified with an equal volume of Freund's complete adjuvant and injected SC into the back of a rabbit. A booster injection was administered 3 weeks later with Freund's incomplete adjuvant and additional injections were administered twice every week. The rabbit was exsanguinated 10 days after the final injection. Western blotting was performed as described previously (Nishibori et al. 1993), with some modifications. The protein-transferred nitrocellulose was blocked with 10% skimmed milk for 1 hr and was then incubated with anti-HMT rabbit serum (1:1500) in 0.1% Tween-20 and 10 mM Tris-buffered saline containing 1% BSA and 5% normal goat serum overnight at 4C. The nitrocellulose was then incubated with biotinylated anti-rabbit IgG–goat IgG (Vector) for 30 min at room temperature and with streptavidin-peroxidase (Sigma) for 30 min. The reaction was developed in the presence of diaminobenzidine, H2O2, and NiCl2.
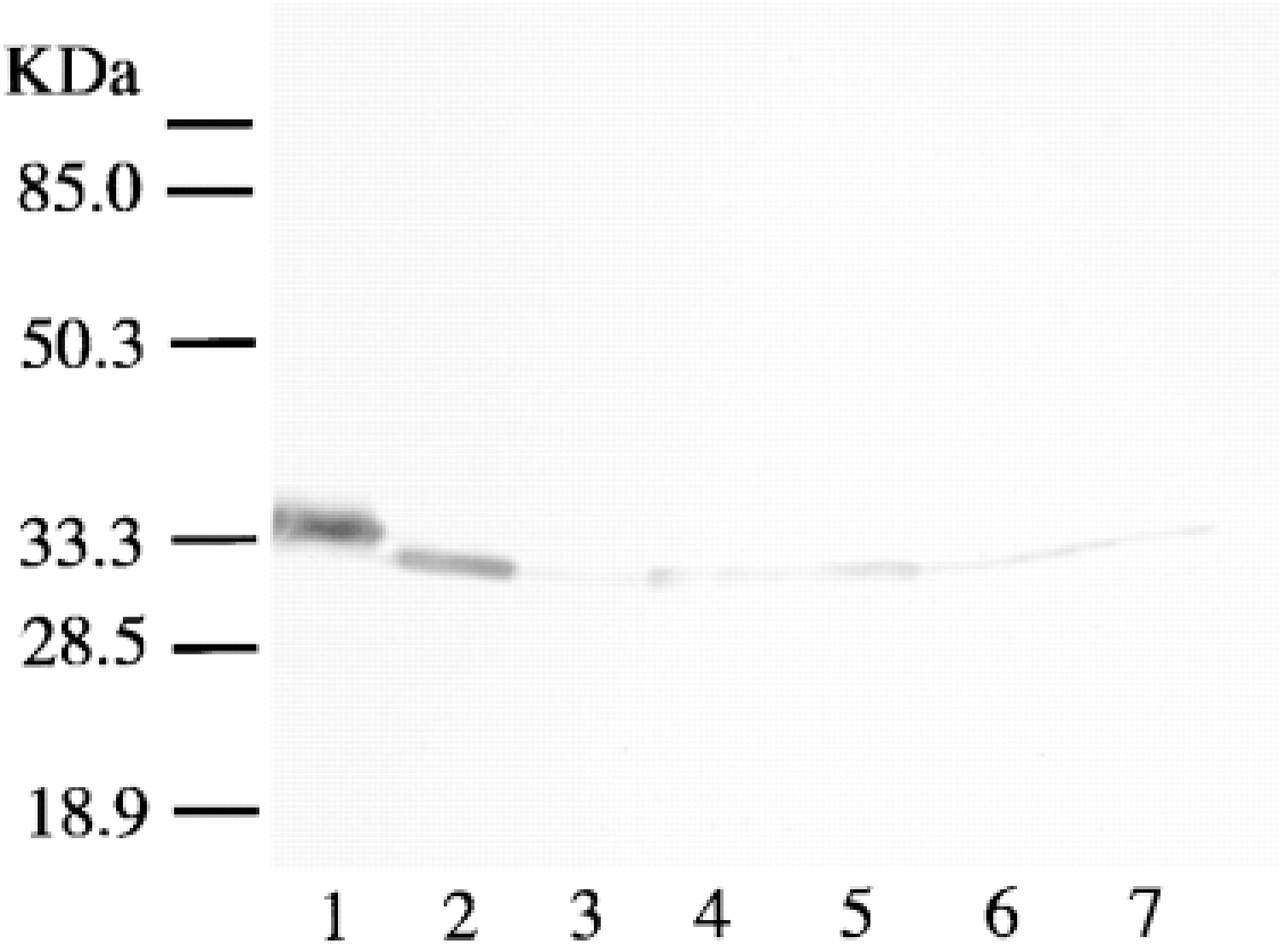
Western immunoblotting of HMT using anti-HMT antiserum. Purified HMT from bovine brain (Lane 1), purified HMT from guinea pig intestine (Lane 2), or supernatant fraction (40 μg protein) of guinea pig tissues (stomach, Lane 3; duodenum, Lane 4; jejunum, Lane 5; ileum, Lane 6; large intestine, Lane 7) was subjected to SDS-PAGE, transferred onto nitrocellulose membrane, and blotted with anti-HMT antiserum (1:1500).
Purification of HMT from Guinea Pig Brain and Intestine
The brain and intestine from 10 male Hartley guinea pigs were homogenized with four volumes of 20 mM Tris-HCl buffer, pH 7.9, containing 1 mM DTT and 1% polyethylene glycol (PEG) (average MW 300). The homogenate was centrifuged at 36,000 × g for 60 min at 4C. To the resultant supernatant, ammonium sulfate was added slowly with gentle stirring to attain 40% saturation, and 20 min later the suspension was centrifuged at 6000 × g for 30 min. The precipitate was discarded and ammonium sulfate was added to the supernatant fraction to attain 75% saturation. The suspension was centrifuged as described above. The precipitate was dissolved in 20 mM Tris-HCl buffer, pH 7.9, containing 1 mM DTT and 1% PEG and dialyzed overnight against the same buffer with three changes of the buffer. The dialyzed sample was applied to a DEAE-cellulose column (2.5 × 12 cm) equilibrated with the buffer and the HMT activity was eluted with 15 mM histamine in the same buffer. The active fractions were pooled and concentrated by an ultrafiltration cell. The concentrate was dialyzed overnight against 10 mM sodium phosphate buffer, pH 7.5, containing 0.15 M NaCl, 1 mM DTT, and 1% PEG, with two changes of the buffer. The dialyzed sample was applied to an Amodiaquine–Sepharose 6B column (5 ml bed volume) equilibrated with 10 mM sodium phosphate buffer, pH 7.5, containing 0.15 M NaCl, 1 mM DTT, and 1% PEG. After extensive washing with 300 ml of the equilibration buffer, HMT activity was eluted with 50 mM histamine dihydrochloride containing 1 mM DTT and 1% PEG. The eluate both from guinea pig brain and from intestine showed a single band on SDS-PAGE with Coomassie Blue or silver staining.
Determination of HMT Activity
The tissues were homogenized with 4 volumes of 50 mM sodium phosphate buffer, pH 7.4, containing 1 mM dithiothreitol and 1% PEG (average MW 300). Centrifugation was conducted at 36,000 × g for 1 hr at 4C, and the resultant supernatants were used to determine the HMT activity in each tissue as previously described (Nishibori et al. 1989). In brief, the reaction mixture (200 μl) was composed of 50 mM sodium phosphate buffer, pH 7.4, containing 50 μM histamine, 50 μM S-adenosylmethionine, 10 μM pargyline, and 20 μl of the enzyme preparation. After a 10-min preincubation, the reaction was performed for 30 min at 37C and was stopped by addition of 50 μl of 1 M NaOH. The telemethylhistamine formed was assayed according to the method of Tsuruta et al. (1981). The HMT activity was expressed as nmol tele-methylhistamine formed/min/mg protein.
Results
Specificity of the Antibody Used
As shown in Figure 1A, anti-bovine HMT antiserum recognized a single band in the samples from different guinea pig tissues, which was the same size as that of the HMT purified from guinea pig intestine and brain. This indicated that the antiserum crossreacted with guinea pig HMT and that the antibody was specific for HMT. The specificity of the antibody was also confirmed in immunohistochemical studies by the lack of staining with preimmune serum (not shown) or immune serum preabsorbed with purified HMT from guinea pig intestine (Figure 2A and 2E). There was no staining with preimmune serum or immune serum preabsorbed with purified HMT in the tissue sections examined in the present study.
Immunohistochemical Staining of HMT-LI in the Alimentary Tract of Guinea Pig
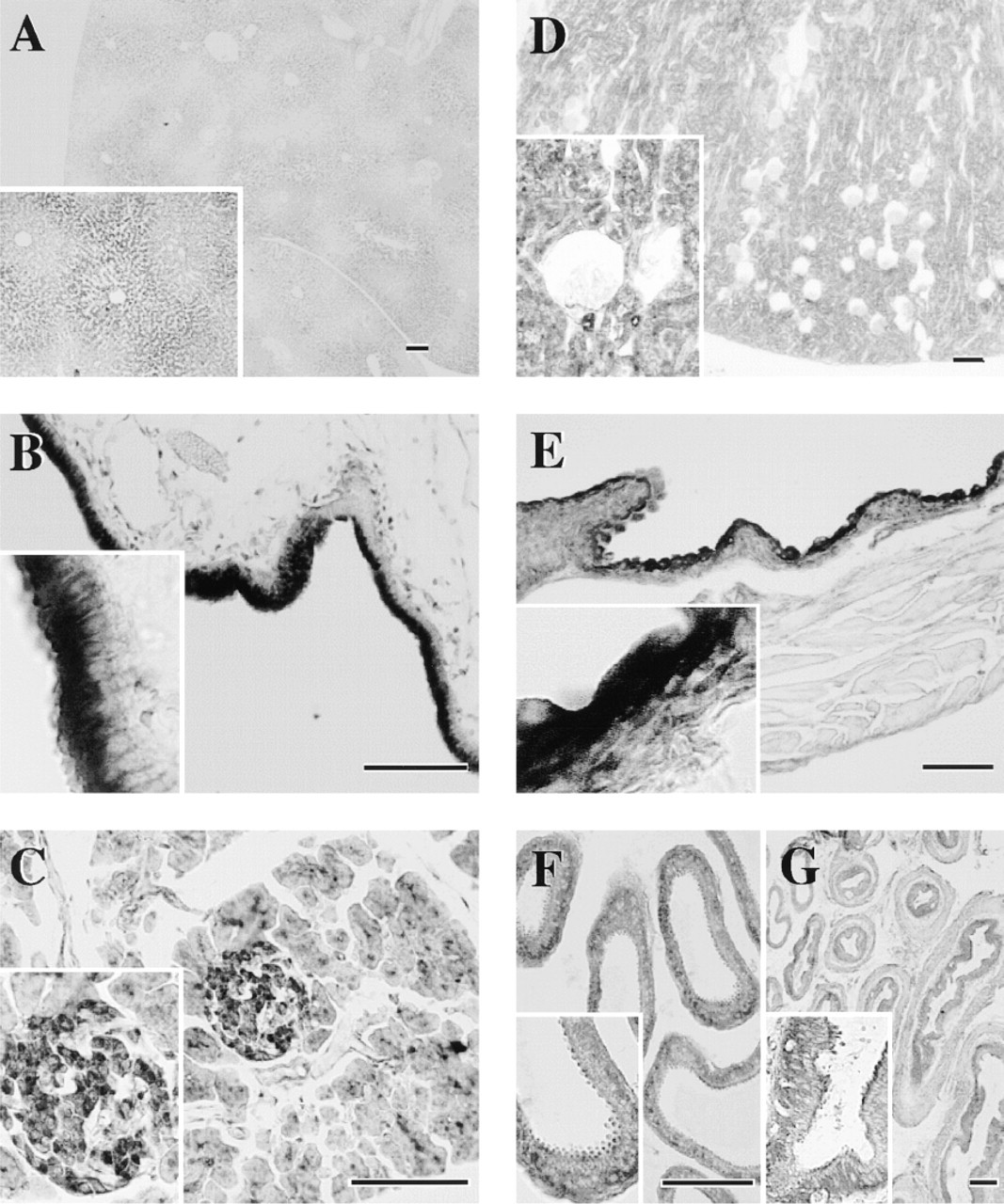
Immunohistochemical studies revealed that HMT-LI was distributed throughout the peripheral guinea pig tissues. In the gastrointestinal tract, the epithelial cell layer from stomach to colon showed HMT-LI (Figures 2–4). In the stomach, the intensity of the staining was higher in the surface mucous cells in the body (Figure 3D). The immunoreactivity in the gastric glands decreased at the glandular base (Figure 3C and 3D). In the cardia and pylorus, the immunoreactive staining in the epithelial cell layer was weaker than that in body glands (Figure 3B and 3E). There were HMT-LI neurons in the myenteric plexus of the stomach (Figure 3F and 3G), duodenum (Figure 2C), jejunum (Figure 2G), ileum (Figure 4B), and colon (Figure 4F). A strong immunoreactive plexus was observed in the intestinal region. Immunoreactive fibers originating from the plexus were also seen in the ileum (Figure 4B, arrowheads). In contrast to the myenteric plexus, the submucous plexus showed no immunoreactivity from stomach to colon. In the liver, the hepatocytes surrounding the central veins were moderately immunoreactive and exhibited a lobular structure. A few lobules were devoid of staining (Figure 5A). In the pancreas, the islets of Langerhans were intermediately immunoreactive and the intensity of the staining differed depending on the type of cell (Figure 5C). The centroacinar cells were moderately stained, whereas the acinar cells were weakly positive (Figure 5C). In the gallbladder, the columnar epithelial layer was strongly immunoreactive (Figure 5B). However, the staining in the submucosa and muscular layer was almost negative.
Immunohistochemical Staining of HMT-LI in the Urinary and Genital Tract of Guinea Pig
In the kidney, all urinary tubule systems, including proximal, Henle, and distal tubules as well as collecting ducts, were intermediately immunoreactive (Figure 5D). On the other hand, renal corpuscles were negative (Figure 5D, inset). In the bladder, the transitional epithelium was strongly immunoreactive (Figure 5E). The smooth muscle cell layer was weakly positive.
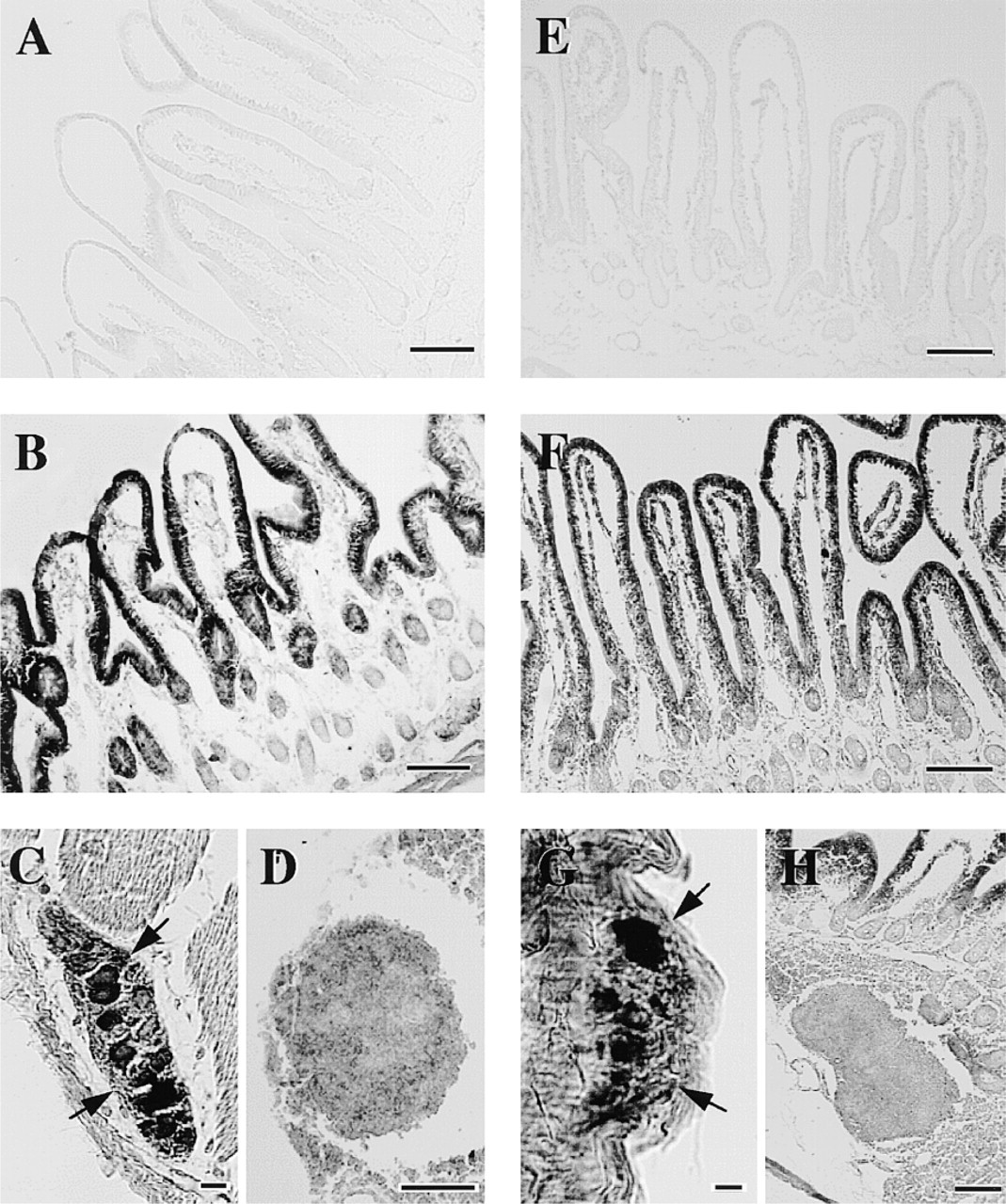
Photomicrographs of sections of guinea pig duodenum (
Immunohistochemical Staining of HMT-LI in the Respiratory Tract of Guinea Pig
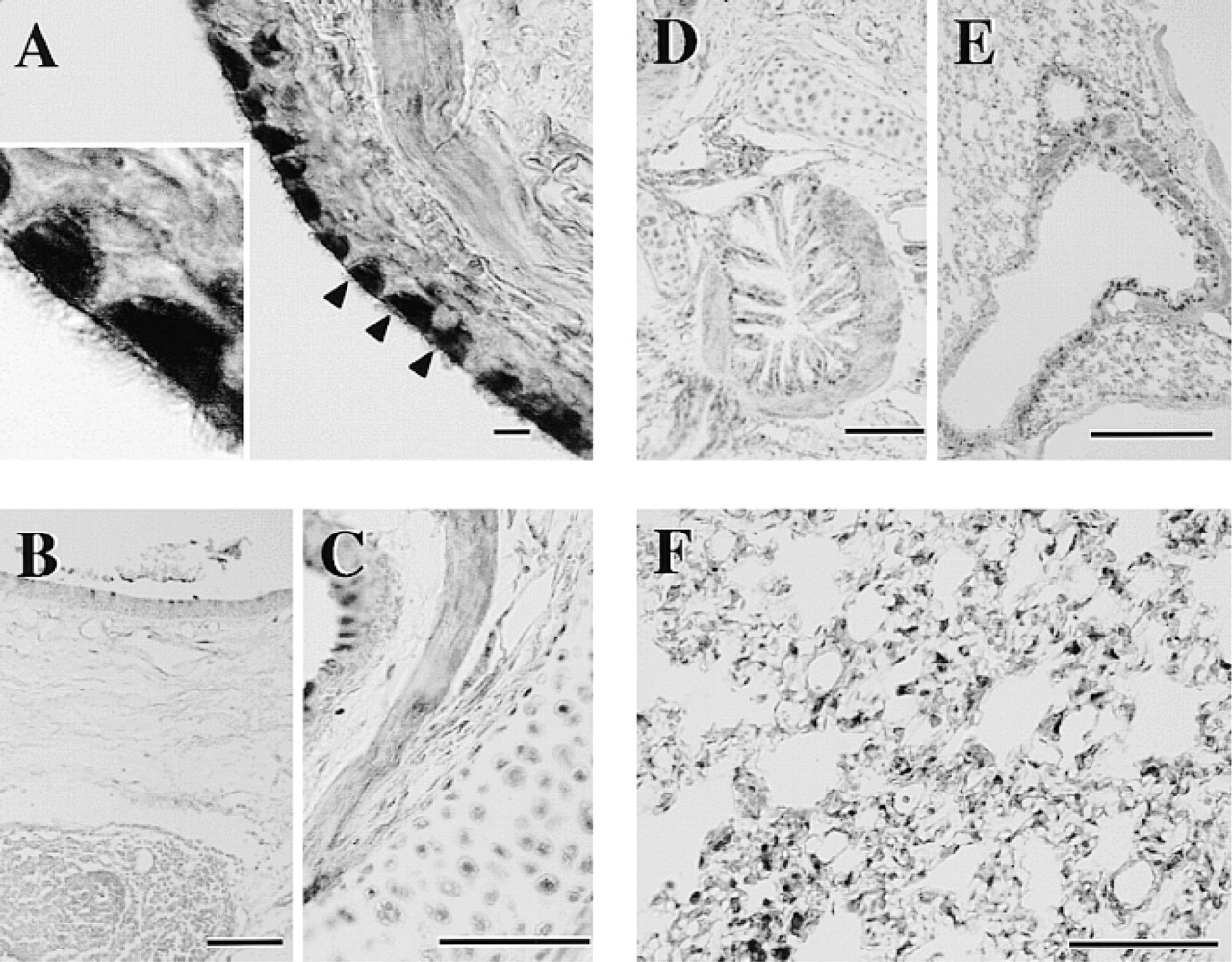
In the respiratory tract, ciliated cells were strongly immunoreactive. The cilia as well as cell bodies were stained homogeneously (Figure 6A). In contrast, goblet cells were devoid of staining. Cartilage cells in the trachea were immunoreactive to some extent (Figure 6C). Tracheal smooth muscle in the posterior portion lacking the cartilage structure was immunoreactive compared with the remaining portions of the ring structure (Figure 6C). In the bronchus, the epithelium, smooth muscle cell layer, and cartilage cells were moderately immunoreactive (Figure 6D). The epithelium and smooth muscle cells in the bronchioles were also moderately stained (Figure 6E). In the lung, irregularly shaped immunoreactive cells were distributed diffusely throughout the alveolar walls (Figure 6F).
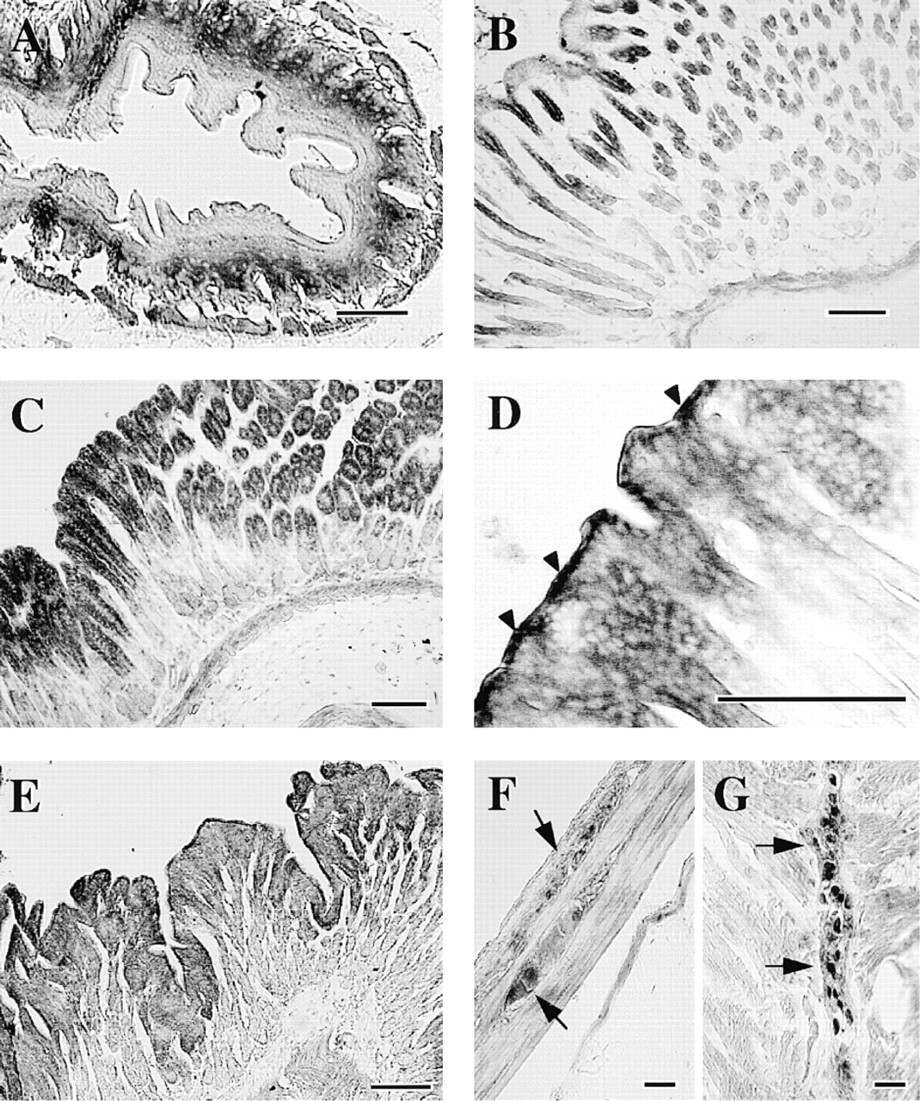
Photomicrographs of sections of guinea pig esophagus (
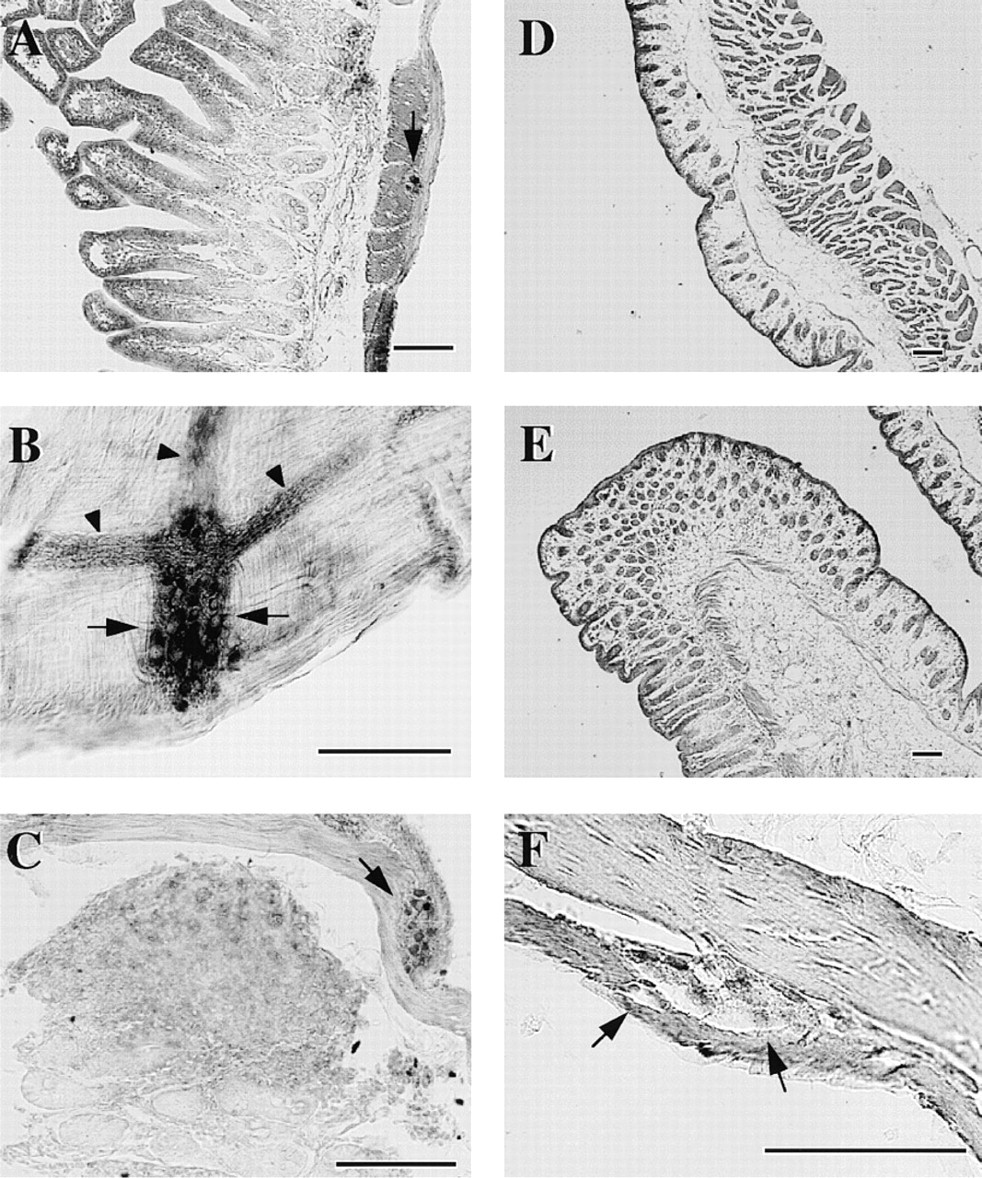
Photomicrographs of sections of guinea pig ileum (
Photomicrographs of sections of guinea pig liver (
Immunohistochemical Staining of HMT-LI in Other Tissues of Guinea Pig
In the skin of nose, ear, and foot pad, the epidermis exhibited intermediate HMT-LI, which distributed with an increasing gradient to the granular layer (Figures 7A, 7C, and 7D). In the hair follicle, the internal root sheath was intermediately immunoreactive and there were a few strongly immunoreactive cells in the external root sheath (Figure 7B). In the tongue, the epithelium was moderately stained (Figure 7E) and the bundles of nerve fibers among the glossal muscle were weakly stained (Figure 7F). The myocardium of the heart was stained moderately in both the atrium and the ventricle (not shown). The tissue distribution of HMT–like immunoreactivity and HMT activities is summarized in Tables 1 and 2.
Photomicrographs of sections of guinea pig trachea (
Discussion
We concluded that the anti-bovine HMT polyclonal antibody recognized the guinea pig HMT specifically because the antibody recognized a single band on Western blotting, using different tissue extracts, which was the same size as purified HMT from guinea pig intestine as well as from brain. Immunohistochemical staining also supported the specificity of the antibody because the antiserum preabsorbed with purified guinea pig as well as preimmune serum did not stain any structure in the tissues examined.
The gastrointestinal tract has the highest HMT activity. Consistent with this finding, the epithelial cell layer of the gastrointestinal tract was strongly immunoreactive for HMT. In general, the HMT-LI in the mucosa distributed along a gradient from the luminal side to the deeper mucosa propria. Specific cells in the mucosal epithelium were not stained. This distribution suggested a role for epithelial HMT in the inactivation of histamine from food.
Endogenous histamine is stored in enterochromaffin-like cells in the gastric mucosa and in enterochromaffin cells in the intestinal mucosa (Panula et al. 1985). The role of histamine released from enterochromaffin-like cells has been well characterized with respect to the acid secretion in the oxyntic gastric gland (Lorenz et al. 1983; Sanders and Soll 1986; Loiselle and Wollin 1993). The acid-producing cells are parietal cells that are present in the deeper and inner portion of the glandular body. The enterochromaffin-like cells are close to but do not make contact with the parietal cells. Therefore, the histamine released from enterochromaffin-like cells appears to diffuse to some extent until reaching the target parietal cells. Loiselle and Wollin (1993) reported that the histamine uptake in the rabbit oxyntic mucosa was coupled with histamine methylation and that the inhibition of HMT by metoprine markedly reduced the net uptake of histamine. Those authors suggested an interaction between the uptake and the methylation of histamine. A similar effect of HMT inhibition on extracellular levels of histamine in the brain was found with an in vivo microdialysis technique (Itoh et al. 1991; Adachi et al. 1992). Therefore, it is possible that the activity of HMT is one of the factors that determines the effective concentrations of histamine in the brain and gastric mucosa. The relatively diffuse localization of HMT in the gastric mucosa corresponds to the disposition of enterochromaffin-like cells and parietal cells, and suggests that the histamine released from enterochromaffin-like cells is taken up and catabolized in neighboring cells, contributing to the removal of histamine from interstitial fluid. The localization of HMT in the gastrointestinal tract was similar from stomach to colon, although the staining intensity was greater in the stomach and upper intestine. In particular, the luminal side of the gastric epithelium was strongly positive. This suggests that epithelial HMT also contributes to the inactivation of histamine from food.
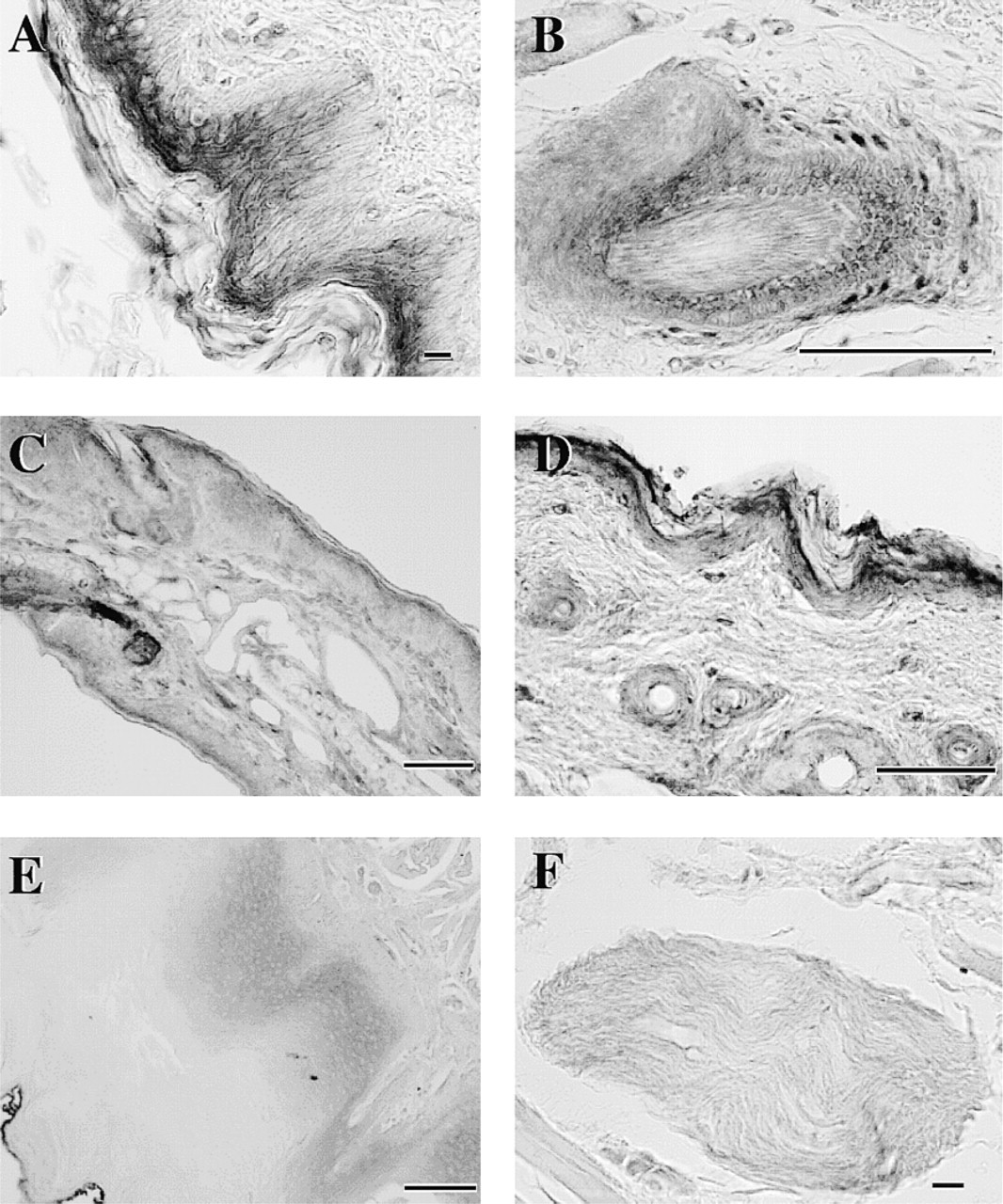
Photomicrographs of sections of guinea pig tongue (
The smooth muscle cell layer of the gastrointestinal tract was intermediately immunoreactive for HMT. The immunoreactivity was relatively high in the ileum, where histamine-induced contraction through the stimulation of H1 receptors has been used for biological assay of histamine. Interestingly, almost all the neurons in the myenteric plexus were HMT-immunoreactive from the stomach to colon. The neurons in the duodenum and jejunum were strongly positive compared with those in other gastrointestinal regions. Panula et al. (1985) found histamine-immunoreactive nerve fibers in the rat and guinea pig gut. They observed histamine-immunoreactive cells in the submucous ganglia after the loading of
Distribution of HMT-like immunoreactivity and HMT activities in gastrointestinal tissues of the guinea pig

aThe intensity of the staining was classified into five categories: –, negative; ±, weak; +, moderate; ++, intermediate; + + +, strong.
bThe results are the mean ± SEM of four experiments.
Distribution of HMT-like immunoreactivity and HMT activities in different guinea pig tissues

aThe intensity of the staining was classified into five categories: –, negative; ±, weak; +, moderate; ++, intermediate; +++, strong.
bThe results are the mean ± SEM of four experiments.
The other strongly immunoreactive cells for HMT were ciliated cells in the trachea. It was reported that the contractile response of guinea pig trachea to exogenous histamine was enhanced by inhibition of HMT and by removal of the tracheal epithelial layer (Ohrui et al. 1992; Yamauchi et al. 1994), clearly indicating the importance of epithelial HMT for the catabolism of histamine in the trachea. An in situ hybridization study suggested epithelial expression of HMT (Ohrui et al. 1992). However, it is apparent from the present immunohistochemical study that ciliated cells rather than goblet cells contribute to the HMT activity in the trachea. Viral (Nakazawa et al. 1994) or mycoplasma infection (Tamaoki et al. 1998) was reported to reduce HMT activity in the airway, suggesting lesioning of ciliated cells on infection. Moreover, histamine has been suggested to be involved in goblet cell secretion (Irokawa et al. 1997; Tamaoki et al. 1997), which was strongly influenced by the activity of HMT (Tamaoki et al. 1997), probably in the neighboring ciliated cells. Therefore, the supersensitivity of airway smooth muscle and mucous secretion to histamine, exogenously administered or endogenously released, appeared to be due in part to the lesioning of ciliated cells and the resulting reduced ability to inactivate histamine. Further work is necessary to test this notion.
The epithelial cells in the gallbladder and the bladder were strongly immunoreactive for HMT. The sensitivity of gallbladder contraction to histamine correlated well with the severity of cystitis (Lennon et al. 1984). In the interstitial cystitis, higher histamine levels were detected in the washings of bladder lumen from patients (Lundeberg et al. 1993). This is consistent with the case in the airway. Thus, HMT may be involved in the physiological control of histamine action in many kinds of tissues, and the reduced activity of HMT resulting from inflammation caused by different stimuli appears to lead to a marked change in the sensitivity of the target cells to histamine. In the rat kidney, HMT-LI was detected only in proximal tubules (Fukuda et al. 1986). In contrast, all urinary tubules were stained in the guinea pig kidney, suggesting a species difference in the distribution of HMT. Moreover, the vascular walls, which were intermediately immunoreactive in the bovine CNS (Nishibori et al. 2000), were not stained in the guinea pig tissues.
In this study we showed the localization of HMT in specific cells in the peripheral tissues. Concerning the alternative pathway of histamine catabolism, involving diamine oxidase, few immunohistochemical data are available. Although Schwelberger et al. (1998, 1999) reported the presence of DAO in the intestine and proximal tubules in pig kidney, the immunohistochemical localization in other tissues remains unclear. Beaven (1982) suggested that the localization of DAO was restricted to the intestine, kidney, thymus, and placenta. The relative importance of the two pathways for histamine inactivation remains to be elucidated.