
Abstract
Type XI collagen is a component of the heterotypic collagen fibrils of fetal cartilage and is required to maintain the unusually thin diameter of these fibrils. The mature matrix form of the molecule retains an N-terminal variable region whose structure is modulated by alternative exon splicing that is tissue-specific and developmentally regulated. In the α1(XI) chain, antibodies to two of the peptides, p6b and p8, encoded by the alternatively spliced exons localized these epitopes to the surface of the collagen fibrils and were used to determine the pattern of isoform expression during the development of rat long bones (humerus). Expression of the p6b isoform was restricted to the periphery of the cartilage underlying the perichondrium of the diaphysis, a pattern that appears de novo at embryonic Day (E) 14. P8 isoforms appeared to be associated with early stages of chondrocyte differentiation and were detected throughout prechondrogenic mesenchyme and immature cartilage. After E16, p8 isoforms gradually disappeared from the diaphysis and then from the epiphysis preceding chondrocyte hypertrophy, but were highly evident at the periarticular joint surface, where ongoing chondrogenesis accompanies the formation of articular cartilage. The spatially restricted and differentiation-specific distribution of α1(XI) isoforms is evidence that Type XI collagen participates in skeletal development via a mechanism that may be distinct from regulation of fibrillogenesis.
T
Type XI collagen is a member of the family of fibrillar collagen genes (Ramirez et al. 1990). Although most abundant in cartilage, its expression is not limited to this tissue (Lui et al. 1995a; Yoshioka et al. 1995). It is a heterotrimer composed of three chains, α1, α2, and α3 (Eyre and Wu 1987). The MA613 chain is a product of the COL2A1 gene (Furuto and Miller 1983; Oxford et al. 1994), which produces Type II collagen, the predominant fibrillar collagen in cartilage. Both the α1 and α2 chains are distinct gene products and contain large complex amino terminal domains (see Figure 1) (Yoshioka and Ramirez 1990; Zhidkova et al. 1993). The amino terminal domain of the α2 chain begins with a disulfidebonded amino propeptide (Npp) of about 250 amino acids, originally isolated as an independent cartilage protein called PARP (proline-arginine rich protein) (Neame et al. 1990). The α1 chain contains an amino propeptide homologous to the α2-Npp, followed by a variable region (Zhidkova et al. 1993), and finally by the minor triple helix characteristic of all of the fibrillar collagens.
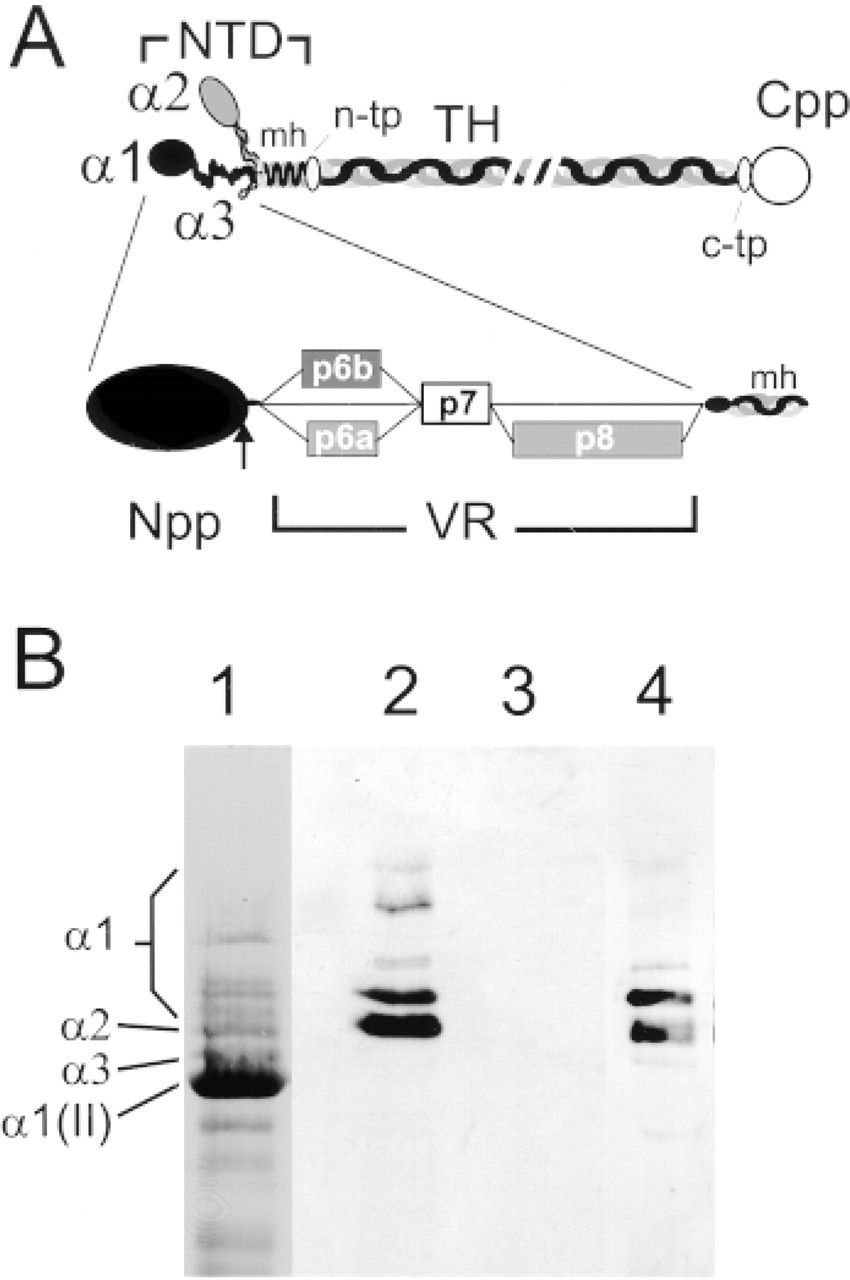
Structure of the amino-terminal domain of Type XI and isoforms of the α1(XI) chain arising by alternative splicing. (
The variable regions of the α1 and α2 chains are not homologous. In both cases, considerable sequence and presumably structural complexity results from alternative splicing of three exons encoding peptides in this region (Oxford et al. 1995; Tsumaki and Kimura 1995; Zhidkova et al. 1995). During cartilage formation and chondrocyte differentiation, complex splicing of the α2 chain rapidly converges to the predominance of a single form lacking all three alternatively spliced exons (Tsumaki and Kimura 1995; Sugimoto et al. 1998) and a highly truncated variable region. In the α1 chain, the variable region is encoded by consecutive exons 6A, 6B, 7, and 8, formerly designated I, II, III, and IV (Zhidkova et al. 1995) or v1a, v1b, c2, and v2 (Oxford et al. 1995). Alternative splicing of exon 6A (peptide p6a), exon 6B (peptide p6b), and exon 8 (peptide p8) generates six possible protein isoforms, p6a+p8, p6b+p8, p8, p6a, p6b, and p0 (containing p7, but lacking p6a, p6b, and p8) (Figure 1A). Mesenchymal cells express the p6a+p8 isoform typical of non-cartilaginous tissues. Differentiation of mesenchymal cells to chondrocytes in micromass culture leads to the expression of five of the isoforms (the p6a isoform is in very low amounts), and p0 and p6b are the most abundant. These isoform-specific peptides can be detected immunochemically both in cell culture and in fetal cartilage tissue (Davies et al. 1998). P6a and p8 are acidic peptides (pI 3.4), whereas p6b is a very basic peptide (pI 11.9) containing clusters of three, four, and five basic residues, mostly lysine (Oxford et al. 1995; Zhidkova et al. 1995).
It is likely that the amino terminal domain, or parts of it, are responsible for the effect of Type XI collagen on limiting fibril diameter (Eikenberry et al. 1992), presumably by steric hindrance of the close packing of Type II collagen molecules in the fibril. The p6a, p6b, and p8 peptides encoded by the alternatively spliced exons may contribute to this function, directly or indirectly, or they may contribute additional functionality to the collagen molecule. The removal of the α2-Npp is relatively rapid (t1/2 1 hr), but less than 50% of the α21-Npp is processed after 20 hr of organ culture of embryonic chick sterna (Thom and Morris 1991). This is extraordinarily slow relative to the rate of matrix production, collagen fibril formation, and tissue growth. Proteolytic processing occurs between the amino propeptide region and the variable region (Zhidkova et al. 1993; Rousseau et al. 1996), such that all of the variable region and minor helix are retained in the mature matrix form of the molecule. The retained variable regions are incorporated into the fibril and the isoforms show distinct patterns of expression in cartilage. For example, the p6a isoform is found predominantly in the proximal rib, the portion that becomes bone, and is scarce or is absent from the distal portion which will become permanent cartilage, even though Type XI collagen is generally distributed in the fetal rat rib cartilage. This suggests a linkage between the expression of the p6b isoform and the complex process of endochondral ossification (Davies et al. 1998). The p6a+p8 and p8 isoforms are prevalent in prechondrogenic mesenchyme and immature cartilage and may be involved in the process of chondrocyte differentiation (Davies et al. 1998). To further examine the relationship between isoform expression and skeletal development, we have investigated the pattern of expression of the p6b- and p8-containing isoforms during the development of fetal rat long bones, primarily the humerus. As in the rib, p6b showed a restricted distribution in the cartilage of the developing humerus. Before primary ossification, p6b was detected only in the diaphysis, primarily adjacent to the perichondrium, and not in the epiphysis. This pattern arose de novo after differentiation of the cartilaginous rudiment and concomitant with the formation of an organized perichondrium as reflected by the staining pattern of Type XII collagen. The p8-containing isoforms were most strongly expressed in areas of newly forming cartilage and disappeared as chondrocyte maturation proceeded. The distinct distribution of isoforms of the α1(XI) chain in the cartilage of developing limbs suggests the potential for unique functional capability beyond the regulation of collagen fibril formation.
Materials and Methods
Antibodies
Antibodies recognizing various epitopes on the collagen α1(XI) chain as well as collagens X and XII have been previously described. These include monoclonal and rabbit polyclonal antibodies raised against a 20-amino-acid sequence within the p8 peptide, a monoclonal antibody raised against the intact p6b peptide (Davies et al. 1998), rabbit polyclonal antibodies raised against the α1(XI) carboxyl telopeptide (c-tp) (Li et al. 1995; Davies et al. 1998), a rabbit polyclonal antibody raised against Type X collagen (Davies et al. 1998), and a rabbit polyclonal antiserum raised against Type XII collagen (Keene et al. 1991). A monoclonal antibody to Type II collagen was purchased from NeoMarkers (Union City, CA). Secondary antibodies linked to FITC or CY3 were obtained from Sigma (St Louis, MO). A rabbit polyclonal antiserum recognizing the p7 peptide encoded by exon 7 was prepared as previously described (Oxford et al. 1994). The full-length p7 peptide, ANIVDDFQDYNYGT-METYQTESPRRVSGSNE(C), was prepared by peptide synthesis in the analytical core facility of Shriners Hospital (Portland, OR). The carboxy terminal cysteine residue was included for coupling to carrier and was not part of the original sequence. Analysis of antibody specificity using immunoblotting of an SDS-PAGE gel was performed as previously described (Davies et al. 1998).
Animals
Timed pregnant rats and pups were purchased from Simonsen (Gilroy, CA). Animals were sacrificed according to the guidelines of the Institutional Review Board of the Oregon Health Sciences University.
Immunohistochemistry
Frozen sections were prepared and analyzed as previously described (Sakai et al. 1986). Acetone-fixed, paraffin-embedded sections (AmeX procedure) were prepared according to the protocol of Sato et al. (1986). Briefly, freshly obtained tissues were rinsed in PBS and placed in acetone at room temperature (RT). After at least 4 hr, the samples were moved to −20C overnight. Samples were then placed successively into fresh solutions of acetone, 4C, 15 min; acetone at RT 15 min; and methyl benzoate, RT, 30 min. Tissue samples were then placed in cassettes and into an automated tissue processor (Citadel 1000; Shandon, Cheshire, UK) for cycling through two changes of xylenes, 30 min each, and two changes of melted paraffin, 1 hr each, the last under vacuum. Six-μm sections were cut on a Reichert-Jung model 820 II microtome (Nussloch, Germany). Sections were deparaffinized in xylene and rehydrated through graded ethanol series before immunohistochemistry.
Unmasking
Sections (frozen or paraffin) were routinely treated with chondroitinase ABC (Sigma), 0.25 ul/ml in PBS for 30 min at RT to remove proteoglycans. As a control, additional unmasking was attempted utilizing 6 M guanidine-HCl buffered in 0.05 M Tris to pH 7.5, or 0.5 M acetic acid, or digestion with the protease ficin (Zymed; San Francisco, CA).
Immunofluorescence Micrographs
Some immunofluorescence images were obtained with a Zeiss Axiophot fluorescence microscope as color slides, which were then digitized. Other images were obtained using a Nikon E800 microscope equipped with a Sensys digital camera (Photometrics; Tucson, AZ) and utilizing Metamorph software (Universal Imaging; West Chester, PA).
Histology
Paraffin sections were rehydrated as above and stained with silver nitrate (von Kossa method) to visualize calcified areas of the tissue and were counterstained with safranin-O to visualize proteoglycans (Clark 1980).
Ultrastructural Localization
Localization of p6b and p8 epitopes by immunoelectron microscopy was performed as previously described (Keene et al. 1995). A solution of 6-nm colloidal gold was obtained from Aurion (Wageningen, The Netherlands) and was coupled to the p6b and p8 MAb according to the manufacturer's instructions.
Results
Isoform-specific Antibodies
Characterization of the expression of the α1(XI) protein isoforms during the cartilaginous phase of long bone development required the use of isoform-specific antibodies. It was not possible to generate antibodies specific to each of the six isoforms because the three peptides modulated by alternative splicing occur in more than one isoform (Figure 1a). The monoclonal antibody specific to the p6b peptide recognizes both the p6b+p8 and the p6b isoform. It is likely that p6b antibody detects primarily the p6b isoform, because the patterns obtained with the p6b and p8 antibody did not significantly overlap at later times of development (see below) and the p6b+p8 isoform is expressed at low levels (Oxford et al. 1995; Davies et al. 1998). The p8 monoclonal and polyclonal antibodies recognize both p6a+p8 and p8 isoforms, which predominate very early in chondrocyte differentiation (Davies et al. 1998). An antibody specific for p6a was not obtained, but the p6a isoform is minimally expressed in cartilage (Davies et al. 1998). It is not possible to make an antibody specific to the p0 isoform. The absence of peptides at two different sites in p0 precludes the use of a junctional peptide. However, this is the most abundant isoform in cartilage (Rousseau et al. 1996; Davies et al. 1998) and is likely to be well represented by the signal obtained with the splicing-independent antibody recognizing the carboxyl telopeptide (c-tp) of the α1(XI) chain. As an additional control for the isoform-independent distribution of α1(IX) chains, a polyclonal antibody specific for the p7 peptide in the variable region was prepared (see Materials and Methods). The p7 peptide, encoded by exon 7, is constitutively expressed and is therefore present in all six isoforms, and is topologically adjacent to both the p6b and p8 peptides. The p7 antiserum specifically recognized α1(XI) chains in an extract of cartilage (Figure 1B), yielding a pattern of immunoblotting identical to that of the previously characterized c-tp antibody. Bands relating to α1(II) or α2(XI) were not recognized, and all blotted bands were collagenase-sensitive.
Localization of the α1(XI) Collagen Chain and Its Isoforms in Fetal Cartilage
E17 and E18 fetal rat humerus were initially examined for the distribution of α1(XI) isoforms. At this stage (Figure 2), the central diaphysis had been invaded by vascular tissue and endochondral ossification was under way. The cartilage (stained with safranin-O) adjacent to the ossification front was hypertrophic and mineralized (stained black with von Kossa stain), while the mineralized bony collar in the periosteum surrounded much of the remaining diaphyseal cartilage (Figure 2C). Well-differentiated perichondrium/periosteum covered the diaphysis. At the epiphysis, the joint cavity was formed and the cartilage appeared relatively uniform. The articular surfaces were covered by immature chondrocytes and mesenchymal cells in a layer defining the developing articular cartilage (Figure 2B).
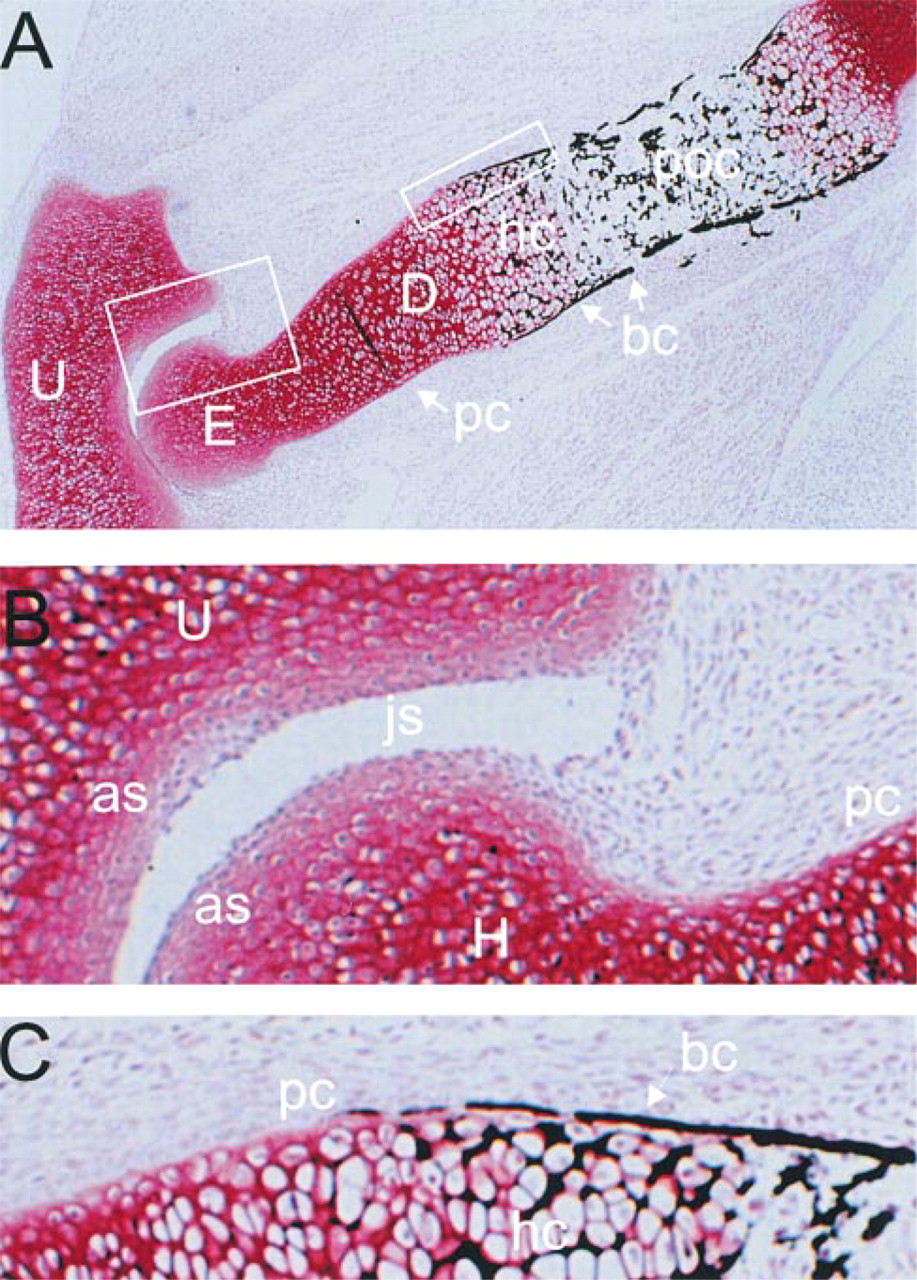
Histology of E18 fetal rat elbow. (
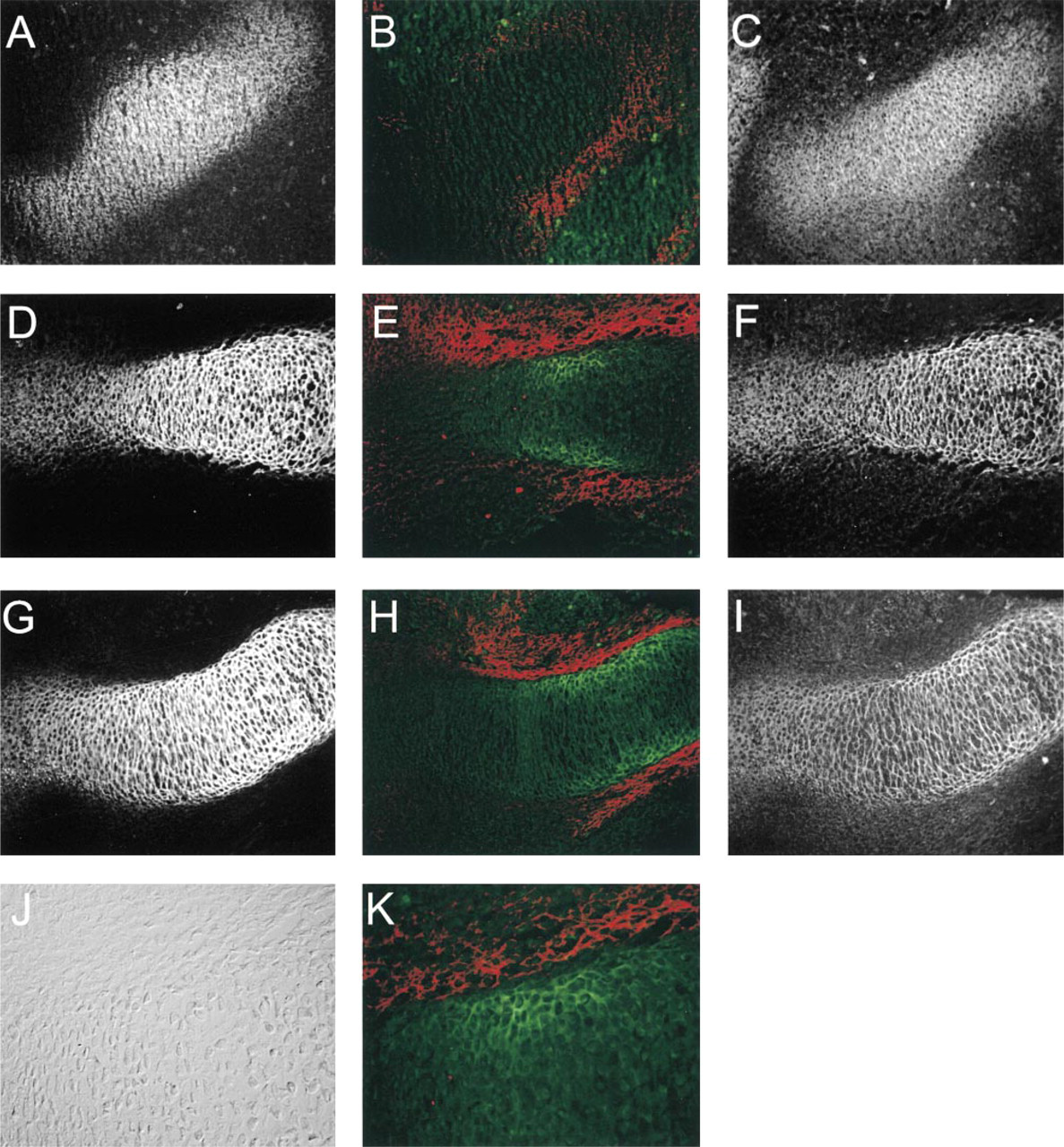
In the distal humerus of E17/E18 rat fetuses, antibodies to Type II collagen defined the cartilaginous zones (Figure 3A). The general distribution of the α1(XI) chain, as determined by the c-tp antibody, was found to overlap that of Type II, except that staining was less intense at the distal third of the epiphysis and was somewhat less in the diaphysis (Figure 3B). The general distribution of the α1(XI) chain was also examined by labeling with the antibody to the p7 peptide (Figure 3C). In this case, the diminished staining of the distal epiphysis was less pronounced, being primarily restricted to the margin adjacent to the joint space. This suggests that the c-tp epitope of the α1(XI) chain is not uniformly available to the antibody. The p8 antibody preferentially labeled the epiphysis (Figures 3D-3F). The staining was intense out to the extreme edge of the joint surface in both the humerus and the ulna, as observed with Type II collagen. Towards the diaphysis, overall staining became weaker and discontinuous; strong, even staining was limited to the lateral margin adjacent to the perichondrium. Staining for p8 was very weak in the hypertrophic zone (compare to Figure 3G). Double labeling with c-tp and p8 antibodies (Figures 3E and 3F) showed that these epitopes co-distributed in the proximal epiphysis and the adjacent diaphysis. However, p8 predominated at the most distal epiphysis and joint surface, whereas c-tp staining predominated in the diaphysis towards the middle of the bone. Higher magnification of this double stain (Figure 3F) showed that p8 surrounded small clusters of cells, whereas c-tp labeled individual cells. This change in p8 staining from the epiphysis to the diaphysis is shown in more detail in Figure 4, comparing in this case p8 and p7 antibodies. Staining with p8 antibodies was general in the epiphysis (Figure 4B), although a few septa between cells labeled much more intensely with p7 than with p8 antibodies (Figure 4C). In the diaphysis, there was incomplete and discontinuous staining with the p8 antibody (Figures 4E and 4F). This suggests that more recently synthesized Type XI collagen did not include p8 isoforms of α1(XI) and that this new matrix and subsequent cell division displaced the existing p8 isoform-containing matrix.
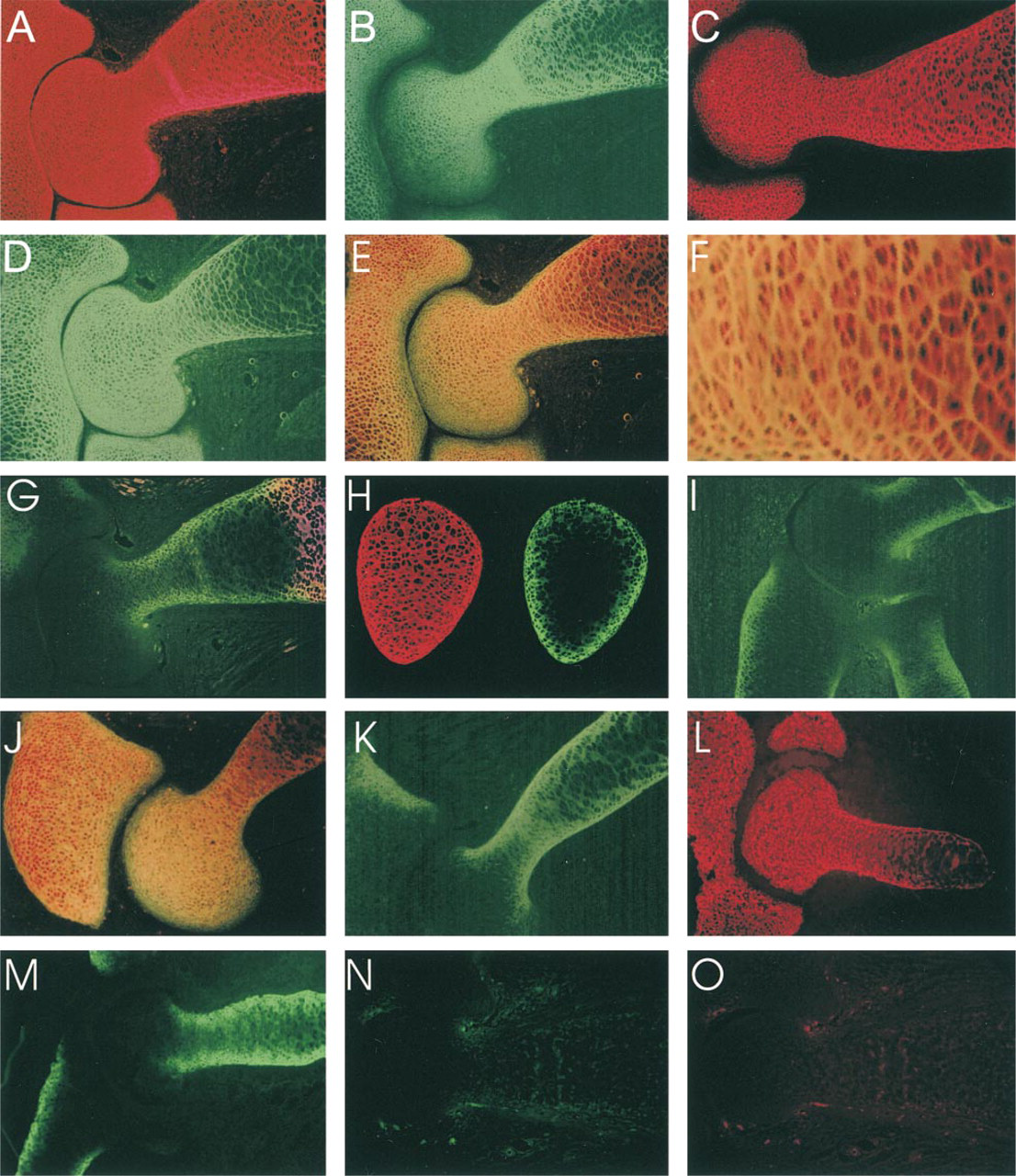
Distribution of p6b- and p8-containing isoforms of the α1(XI) collagen chain in the distal humerus of E18 fetal rat. (
Gradient of p8 isoform distribution from the epiphysis to the diaphysis. E18 humerus labeled with antibody to p8 (green) and isoform-independent antibody to peptide p7 (red). (
More extreme asymmetric labeling of cartilage in the humerus was observed with antibodies to p6b isoforms (Figures 3G-3I). Staining for p6b was most intense at the lateral boundary of the diaphyseal cartilage and became weaker and difficult to detect in the central region farthest away from the perichondrium. Staining was completely absent from the epiphysis. Staining for p6b was not restricted to hypertrophic cartilage, identified by staining with antibodies to Type X collagen, but did overlap slightly at the margin (Figure 3G). Examination of cross-sections of the diaphysis (Figure 3H) confirmed the peripheral staining pattern for p6b isoforms (green) in contrast to the uniform staining observed with the p7 antibody (red). This asymmetric distribution of p6b labeling was also apparent in the ulna and radius (Figure 3I).
The pattern of labeling of p8- and p6b-containing isoforms of the α1(XI) chain at this stage of development, as well as the nonuniform labeling of α1(XI) with the c-tp antibody, was not altered by pretreatment of sections with 6 M guanidine (Figures 2J and 2K) or acetic acid, mild pepsin digestion, or proteolysis with ficin (not shown). Utilization of frozen sections (Figures 2K and 2M) also did not fundamentally alter the staining pattern with antibodies to p6b or p8. Repeated experiments showed that the broader p6b staining reaching the central diaphysis in Figures 2K and 2M probably resulted from the location of the plane of section closer to the perichondrial surface of the humerus. Other contributing factors, such as section thickness (15 μm for frozen, 6 μm for AmeX), better preservation of epitopes in frozen sections, or unmasking of some epitopes by treatment with guanidine may also be involved.
The localizations of p8 and p6b isoforms were refined by comparison to the distribution of Type XII collagen (Figure 5), used here as a marker for non-cartilaginous connective tissues containing primarily Type I collagen. Antibodies to collagen XII labeled the perichondrium. The epiphyseal cartilage underlying the joint surface was also labeled. The p8 staining overlapped with Type XII collagen staining in this distal epiphyseal region, but the matrix corresponding to the layers of cells at the extreme articular surface stained only with collagen XII antibodies (Figure 5A). The p6b and collagen XII antibodies recognized adjacent but almost mutually exclusive domains in the diaphysis (Figures 5B and 5C). Collagen XII antibodies stained the perichondrium, whereas p6b antibodies labeled the underlying cartilage. Nearer to the epiphysis, there appeared to be a very narrow region of overlap at the cartilage-perichondrial interface, but closer to the hypertrophic zone there was virtually no overlap in staining (Figure 5C). This indicated a rather abrupt transition between perichondrium and cartilage in the developing cartilaginous template.
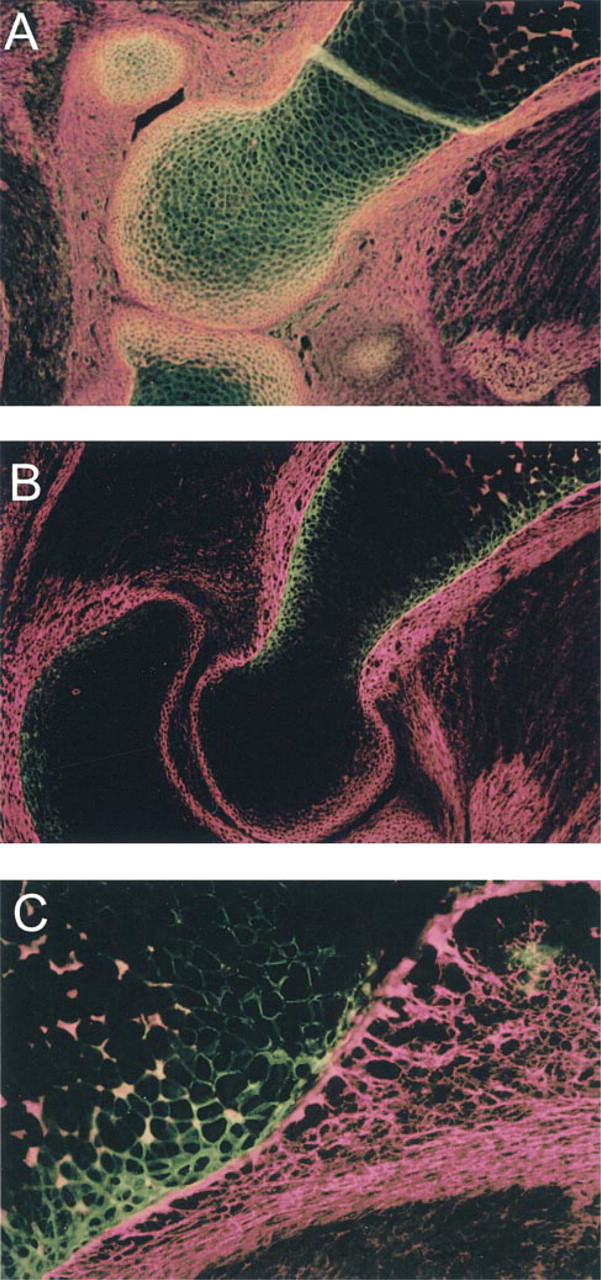
Distribution of p6b and p8 isoforms of α1(XI) relative to collagen XII in E17/E18 fetal rat distal humerus. (
Ultrastructural Localization of p8 and p6b Isoforms
The site of proteolytic processing (Figure 1A) and the matrix staining obtained above with the p6b and p8 antibodies suggested that these peptides in the variable region should be part of the collagen fibrils and available at the fibril surface. This possibility was examined by immunoelectron microscopy using primary antibody-colloidal gold conjugates (Figure 6). The area of p6b labeling shown in Figure 6A was near the end of the bony collar where mineralization was discontinuous (between the hypertrophic zone and the epiphysis; see Figure 2C). The thin collagen fibrils characteristic of fetal cartilage were extensively labeled with p6b antibody. At higher magnification (Figure 6B), the labeling was periodic, with a minimum spacing of about 54 nm corresponding to a periodicity characteristic of the quarter stagger arrangement of collagen molecules within the fibril. The large Type I collagen-containing fibrils and dispersed foci of mineralization of the bony collar were not labeled with p6b antibody. Consistent with the observations by light microcopy, there was no transition or structural gradation between the extracellular matrix of the perichondrium and the underlying cartilage. The thin fibrils of cartilage were juxtaposed to the thick, banded fibrils of perichondrium. Examination of the distal epiphyses with antibodies to p8 showed that these collagen fibrils were also extensively labeled, but it was more difficult to demonstrate periodic labeling (Figure 6C). At the level of light and electron microscopy, the p6b and p8 peptides in their respective α1(XI) isoforms were accessible without need for unmasking, except for the removal of proteoglycans with chondroitinase.
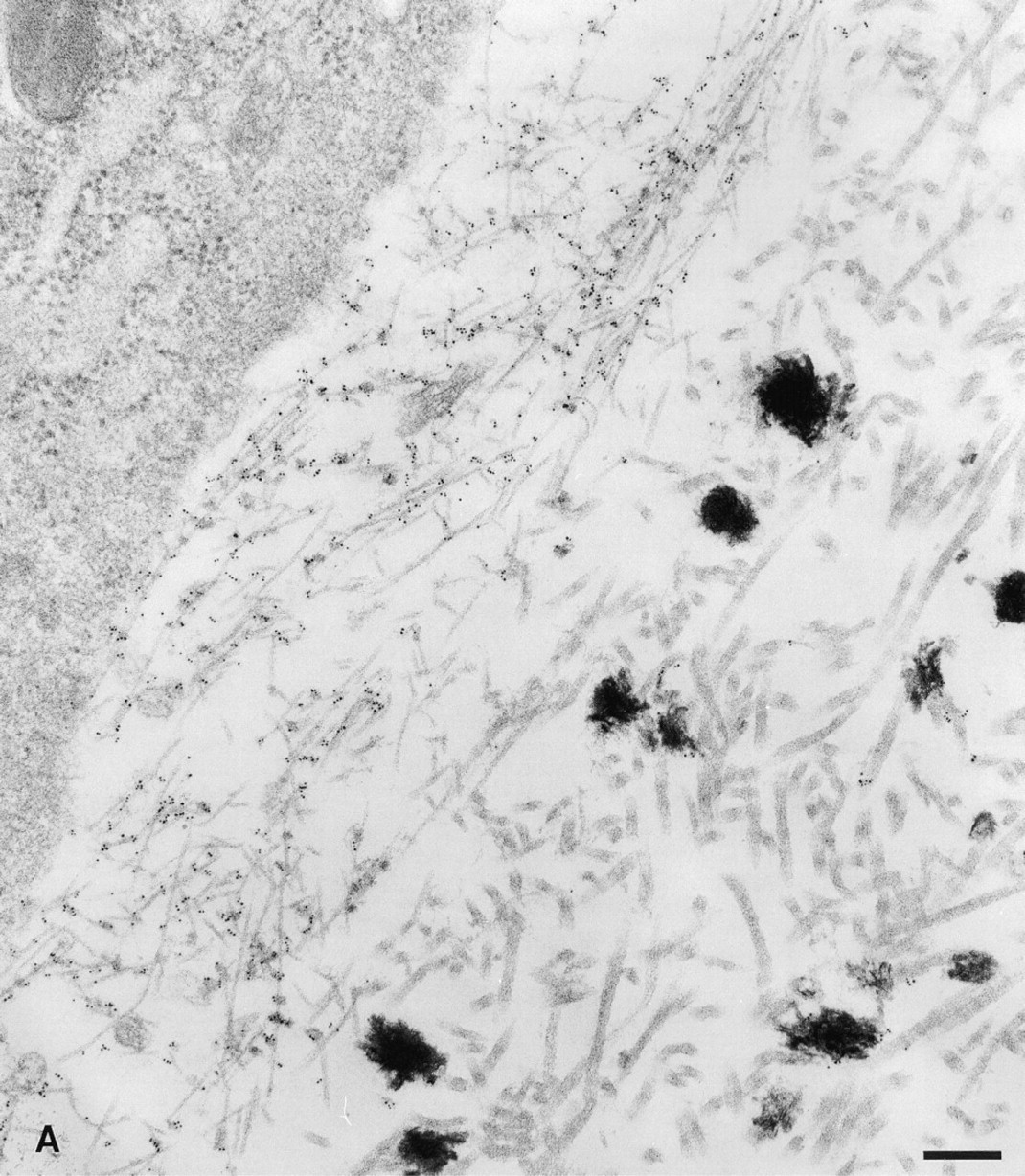
Ultrastructural localization of p6b and p8 peptides to the surface collagen fibrils. Five-nm colloidal gold particles directly conjugated to p6b and p8 primary antibodies were used to localize these isoforms and epitopes in the tissue. (
Developmental Expression of p6b-and p8-containing Isoforms
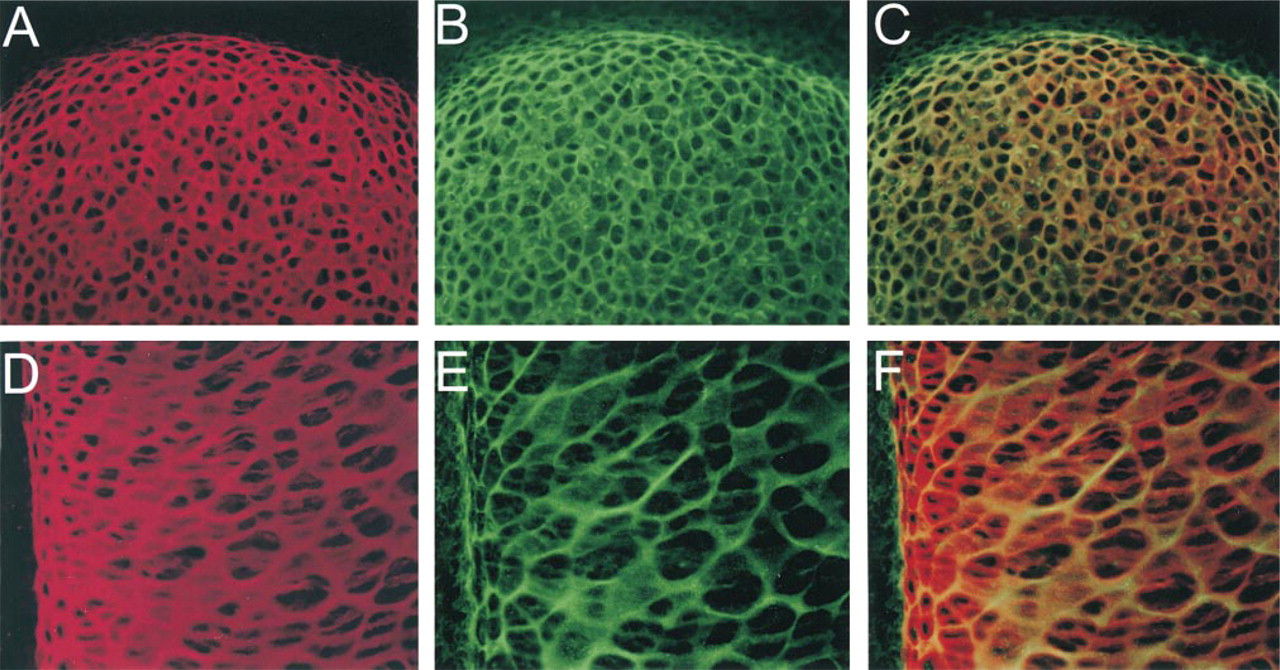
It is possible that the asymmetric and almost mutually exclusive distributions of p6b and p8 arose at the time of cartilage formation. Alternatively, both sets of isoforms could have been uniformly expressed initially and then have become restricted as development and chondrocyte differentiation proceeded. To distinguish between these two possibilities, the distribution of p6b- and p8-containing isoforms was examined at the initial stages of cartilage formation in the rat humerus, E13.5-14.5 (Figure 7). At E13.5 (Figures 7A-7C), the condensing mesenchyme/immature cartilage of the humerus was generally labeled with the p7 antibody (Figure 7A), stronger in the middle and weaker at the ends where differentiation (or chondrogenesis) was still occurring. At this developmental stage, the tissue was highly cellular, with minimal extracellular matrix, and there was no staining for p6b. Diffuse staining for Type XII collagen around the condensation indicated a poorly differentiated perichondrium (Figure 7B). Staining for the p8 isoform was general and resembled that of the p7 antibody (Figure 7C). At E14 (Figures 7D-7F), the extended region of the anlage (diaphysis) contained more differentiated chondrocytes surrounded by matrix that labeled well with the p7 antibody (Figure 7D). The end of the anlage (epiphysis) was stained less intensely and resembled the E13.5 tissue. The perichondrium was well labeled with collagen XII antibodies and had become striated or fibrillar in appearance (Figure 7E), indicating a more advanced stage of differentiation. Staining with the p6b antibody became apparent in a highly focal zone of chondrocytes adjacent to the perichondrium, but not deeper into the cartilage. The staining pattern with the p8 antibody was similar to that of p7, although not as intense (Figure 7F). By E14.5, the region of strong labeling of the p7 epitope in the diaphysis was more extended (Figure 7G). Staining for p6b reached more distally, along with the collagen XII staining of the perichondrium, but was still limited to the region adjacent to the perichondrium (Figure 7H). The collagen XII localization appeared more fibrillar and less diffuse, indicating differentiation of the perichondrium. The staining with the p8 antibody was similar to that on E14 but, in comparison to p7, a more open network was apparent (Figure 7I). This pattern is reminiscent of the p8 staining in the later diaphysis (Figure 3F), although more subtle. The p6b staining at higher magnification is shown in Figures 7J and 7K. Differential interference microscopy revealed that the cells to the left and far right were flattened perpendicularly to the long axis of the forming bone. The cells in the central region appeared rounder, with more matrix between cells, and it was around these cells, adjacent to the perichondrium, that p6b was first detected up to a depth of about five cells.
The distribution of the p6b and p8 isoforms was examined at subsequent stages in the development of the humerus (Figure 8). The α1(XI) chain was generally distributed in the cartilage at E16, as indicated by staining with p7 antibodies (Figure 8A). This pattern was maintained through the regression of cartilage in the diaphysis due to primary endochondral ossification (Figure 8D), the establishment of a metaphyseal growth plate (postnatal Day 3; Figure 8G), and the secondary ossification center in the epiphysis (postnatal Day 7; (Figure 8J). Labeling of the cartilage with the p8 antibody was widespread at E16 (Figure 8B) but rapidly diminished in the diaphysis by E18-E20 (Figure 8E; see also Figures 3D and 4F). Staining of the epiphysis persisted through postnatal Day 7 (Figure 8K). However, by E20 (Figure 8E) the staining was stronger in the developing articular cartilage. By postnatal Day 3 (Figure 8H), the epiphyseal cartilage was only weakly stained, whereas the developing articular cartilage at the joint surface continued to stain strongly, a pattern that was also observed at postnatal Day 7 (Figure 8K). The disappearance of the p8 isoform from the diaphysis and the epiphysis preceded chondrocyte hypertrophy associated with primary and secondary endochondral ossification, as indicated by staining for Type X collagen (Figures 8C-8L).
In contrast, from E16 to postnatal Day 7, the p6b isoform was detected only at the periphery of the diaphyseal cartilage along its entire length, overlapping with Type X staining as the cartilage became hypertrophic, and disappearing only when cartilage became bone (Figures 8C, 8F, 8I, and 8L). Staining for p6b was not detected in association with the secondary ossification center in the epiphysis at postnatal Day 7 (Figure 8L), although it was detectable at the periphery of the well-defined metaphyseal growth plate. Weak, diffuse, and variable staining with the p6b antibody was observed at times in the epiphysis after birth (near the articular surface in Figures 8J and 8L); however, this was an inconsistent finding. By E20, the patterns of immunohistochemical staining for p6b- and p8-containing isoforms were largely segregated (Figure 8H) and remained so (Figures 8H and 8K). This overall pattern of isoform localization during development was also observed in the radius and the ulna (Figures 8E, 8H, and 8K; and data not shown).
Distribution of p6b and p8 Isoforms in Other Skeletal Systems
To determine if the differential staining pattern of α1(XI) isoforms was a general phenomenon, other skeletal elements and cartilage systems were analyzed. Examination of the forepaw of E18 animals revealed a pattern of p6b and p8 staining in the metacarpal bones that was very similar to that of the long bones described above (Figures 9A-9C). The development of metacarpals is similar to that of long bones, except that the distal ends ossify directly from the diaphysis rather than via a secondary ossification center. As before, p6b staining was restricted to the diaphysis, although it was not confined to the periphery. This may be due to the small size of these bones. P8 staining was most pronounced at the ends of the bones, although there was still some staining in the diaphysis. The carpal bones, which ossify much later than the other bones in the hand, stained uniformly with p8 antibody, but p6b staining was not detected, either in the frontal plane shown, which emphasizes the articulating surfaces, or in sagittal section, which revealed the perichondrium at the dorsal and ventral surfaces (not shown). A section through the spine at E18 also revealed differential staining for the p6b isoform (Figure 10B). Compared to the staining with the isoform-independent p7 antibody (Figure 10A), p6b localization was absent from the ends of the neural arch and from the center of the vertebral body. Staining for p6b was observed at the periphery of the vertebral body and throughout the narrow shaft of the neural arch adjacent to the perichondrium (Figure 10C). In this section, the p8 epitope was labeled throughout the vertebral body (Figure 10B) but was stained more strongly at the ends of the neural arch, similar to long bones and digits.
Early developmental expression of Type XI collagen isoforms. Upper forelimbs from E13.5 (
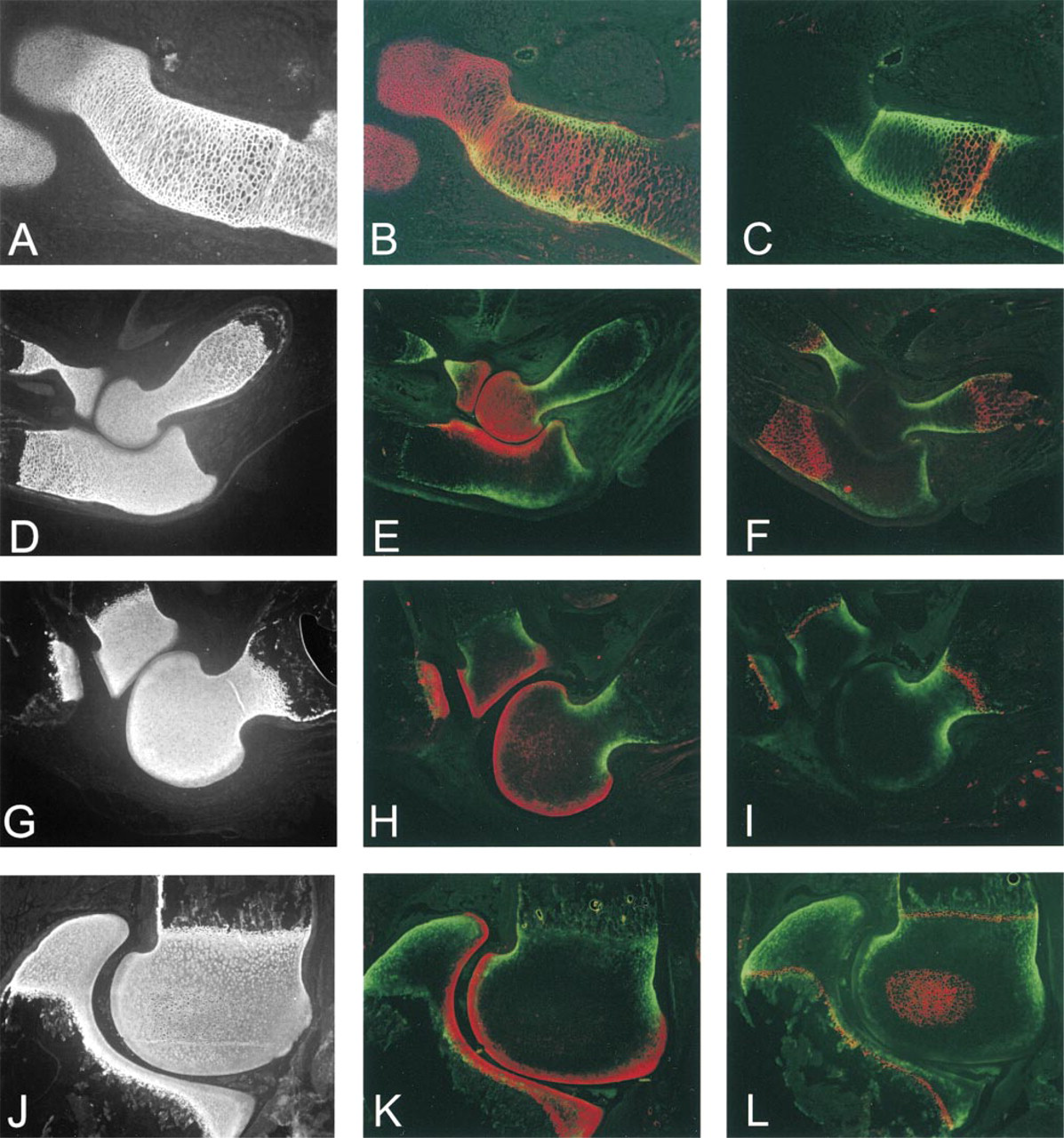
Distribution of p6b and p8 isoforms of α1(XI) during later embryonic and early postnatal development. Upper forelimbs from E16 (
Discussion
During chondrogenesis and endochondral ossification, the extracellular matrix of cartilage serves diverse functions and is produced by chondrocytes according to their state of phenotypic differentiation: chondroblast or immature chondrocyte, mature or prehypertrophic chondrocyte, and hypertrophic chondrocyte. It is not surprising, therefore, that regional heterogeneity of extracellular components would also arise from such differences in support of specific functions or the process of differentiation. The developing articular cartilage matrix produced by mesenchymal cells and immature chondrocytes is enriched in fibromodulin (Archer et al. 1996), tenascin-C (Koyama et al. 1995; Pacifici 1995), and collagen XII and XIV (Watt et al. 1992) (see Figure 5A). Conversely, Type X collagen, as well as bone proteins such as bone sialoprotein and osteopontin, is produced by hypertrophic chondrocytes in the diaphysis and later in the secondary ossification center of the epiphysis (Schmid and Linsenmayer 1985; Bianco et al. 1991, 1993, 1998; Galotto et al. 1994; Gerstenfeld and Shapiro 1996). However, within the bulk fetal cartilage, the differential distribution of isoforms of the α1(XI) collagen chain, as presented here, represents a striking example of regional differences of one protein that is linked both to sequential stages of chondrocyte differentiation and to topological location.
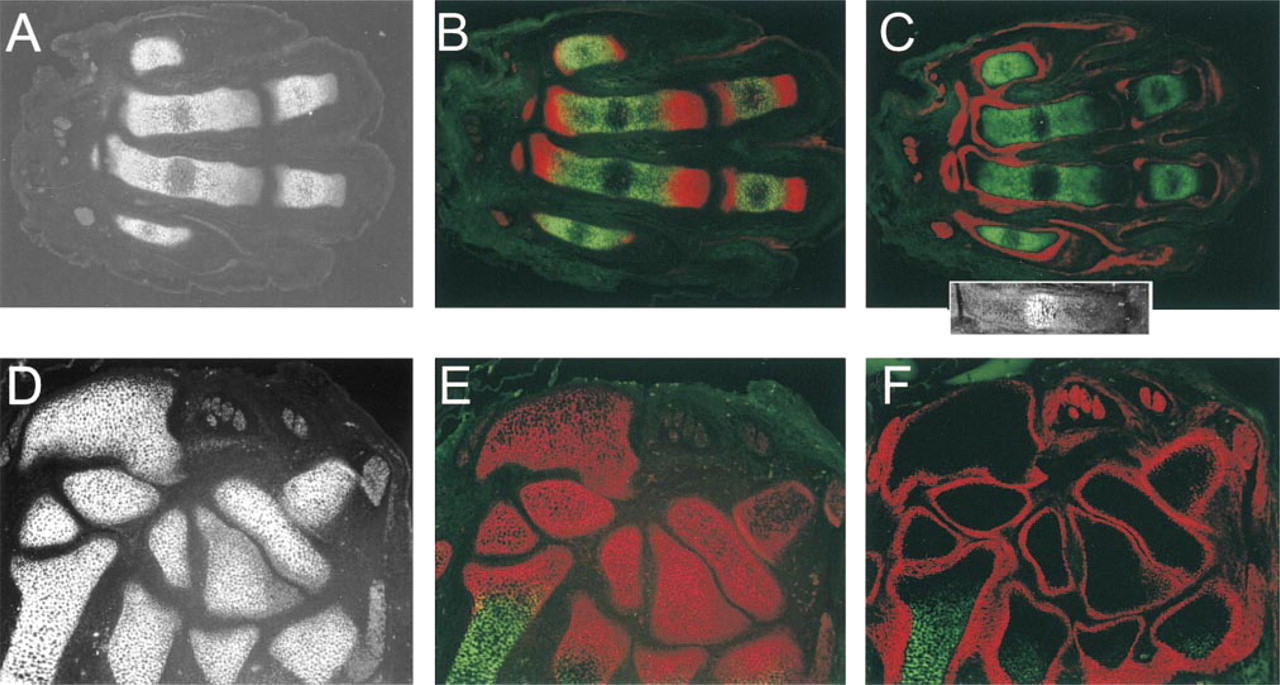
Localization of p6b and p8 isoforms of α1(XI) in fetal rat paw. Sections were taken through the metacarpals and digits (
As depicted schematically in Figure 1, structural studies indicate that the variable region is a distinct domain with a rather extended conformation for all isoforms (unpublished observations). The highly charged composition of the p6a, p6b, and p8 peptides would alter the chemical nature of the variable region according to the isoform and could modulate potential biological activity as well. Lying adjacent to the proteolytic processing site (Figure 1), the peptides could influence the rate of removal of the large N-propeptide and hence fibril morphology. However, despite the differential localization of the p8- and p6b-containing isoforms shown here, no significant difference in fibril diameter was noted in the different regions of developing cartilage (compare Figures 6B and 6C; and unpublished observations). Ultrastructural analysis confirmed that the variable region was an integral part of the collagen fibril because p6b and p8 epitopes were available and were localized at the fibril surface. The functional significance of the isoforms may therefore reside in an ability to mediate interactions between the fibrils and other components of the extracellular matrix.
p8-containing Isoforms Are Associated with Chondrogenesis
The overall pattern of p8 staining reflects the developmental age and stage of the tissue, oldest in the diaphysis and youngest at the margins of the joint. The p8-containing isoforms were found throughout the early differentiating cartilage starting at E13.5, and by 7 days postnatally they were restricted to the area of chondrogenesis at the articular surface, where ongoing differentiation of prechondrogenic mesenchyme supports the formation of articular cartilage (Bland and Ashhurst 1996). In between, p8 staining was observed in a distal to proximal gradient, as p8 disappeared first from the diaphysis and then from the central epiphysis. Removal of p8-containing isoforms from the matrix is not necessarily a prerequisite for chondrocyte hypertrophy because the initial expression of Type X collagen, a well-defined marker for hypertrophic chondrocytes (Schmid and Linsenmayer 1985), was detected at E16 in a cartilage matrix that stains strongly for p8. However, the disappearance of p8 staining from the matrix ahead of chondrocyte hypertrophy, both in the advancing front of primary ossification in the diaphysis and in the secondary ossification at postnatal Day 7, suggests a possible relationship.
Detailed examination of the developmental changes in p8 distribution suggested that such a pattern was unlikely to be sustained at the transcriptional level, although this was not examined directly. It is more likely that p8 isoforms predominantly expressed by early immature chondrocytes persist in the matrix. The localization pattern would therefore derive from the timing and location of turnover or modification of epitope than from synthesis. Turnover could involve degradation of the fibril or proteolytic removal of the variable region from the surface of the fibril. Cessation of p8-containing isoform synthesis with the continued synthesis of other isoforms, predominantly p0, would lead to loss of p8 staining near cells. Synthesis of p0 and cell division displace the peripheral p8-containing matrix, which becomes progressively more difficult to detect, presumably due to turnover. This interpretation is supported by the observation that in the later stages of long bone development, p8 isoforms persist at the articular surface, the only area of active chondrogenesis. This interpretation is also consistent with the observation that in micromass culture, p8 isoforms predominate in prechondrogenic mesenchyme and become a minor component in fully differentiated chondrocytes (Davies et al. 1998). Finally, the distribution of p8 isoforms is analogous to the distribution of the IIA splice form of collagen Type II produced by prechondrogenic mesenchyme and chondroblasts (Sandell et al. 1991, 1994; Ng et al. 1993; Lui et al. 1995b). Whereas expression of the IIA splice form ceases on differentiation, the protein isoform persists in the tissue in a pattern similar to the staining for p8 isoforms of α1(XI) (Oganesian et al. 1997; Zhu et al. 1999).
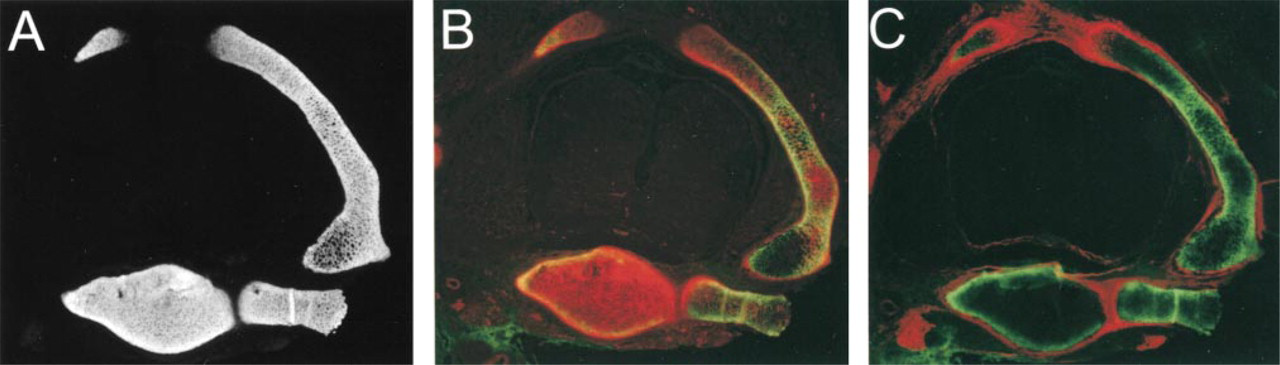
Differential localization of p6b and p8 isoforms of α1(XI) in the vertebrae. Transverse sections were taken through the spine of an E17 rat fetus and immunohistochemically labeled as in Figure 9. (
The function of the p8 isoform in the cartilage of the developing bone is unknown. However, the association of the p8 isoform with areas of chondrogenesis suggests that it may provide an extracellular matrix conducive to chondrocyte differentiation. In this regard, it has recently been shown that the cystine-rich peptide in the amino terminal domain of collagen Type IIA has homology to noggin and can bind to BMP-2, suggesting that modulating BMP/GDF activity could be a function of Type II collagen distinct from its structural role in cartilage (Zhu et al. 1999). There is no homology between the p8 sequence and known signaling elements in skeletal development. Nevertheless, there remains the potential for interaction between p8, as part of the insoluble extracellular matrix, and the soluble growth and differentiation factors that profoundly influence chondrogenesis, chondrocyte proliferation, and hypertrophy, and the development of the epiphysis and joint (Macias et al. 1997; St-Jacques et al. 1999; Storm and Kingsley 1999).
p6b Isoforms Are Associated with Primary Endochondral Ossification
The distribution of the p6b isoform was observed to be highly restricted in the developing long bone. Strongest immunohistochemical staining was adjacent to the perichondrium and diminished towards the interior, eventually becoming undetectable by this method. This pattern is typical of the humerus, radius, and ulna, as well as the femur and tibia (not shown). Ultrastructural analysis clearly showed that the p6b isoform of α1(XI) was a component of the thin collagen fibrils typical of fetal cartilaginous matrix that was indistinguishable from that in deeper unlabeled areas of cartilage. Under the forming bony collar, there was a surprisingly sharp transition to the extracellular matrix of the perichondrium, which is characterized by large, banded collagen fibrils.
Although of different geometry, the vertebral bodies also show preferential localization of p6b at the periphery in cross-section. In the smaller bones, such as the metacarpals and digits, p6b is restricted to the diaphysis but staining was not limited to the periphery similar to the pattern observed in proximal rib (Davies et al. 1998). This is probably due to the narrow diameter of these bones, although other intrinsic differences cannot be ruled out. These observations suggest that the pattern of p6b expression may be a general property of endochondral skeletal development.
The common feature of the p6b distribution was its association with the perichondrium. In the epiphysis, which lacks a differentiated perichondrium, chondrocytes progress through all stages of differentiation and endochondral ossification without deposition of detectable levels of p6b isoform into their matrix. The proximal end of the metacarpal is not a true epiphysis because there is no secondary ossification center. Ossification of this pseudo-epiphysis is a continuation of the primary ossification of the diaphysis. However, p6b-containing isoforms are excluded from the pseudoepiphysis as well. In addition, no p6b was detected in the cuboidal bones of the carpus, whose surfaces are largely articulating. This was true even at the dorsal and ventral surfaces and at late stages of development after the onset of endochondral ossification (unpublished observations). The absence of p6b suggests a developmental relationship between the epiphysis of long bones and cuboidal bones that is distinct from the perichondrium-dependent development of the diaphysis (Rooney and Archer 1992; Long and Linsenmayer 1998).
The restricted distribution of the p6b isoform and its association with the perichondrium arises de novo. At stage E14 in fetal rat (equivalent to about E12.5 in the mouse), p6b was first detected in a group of rounded prehypertrophic chondrocytes at the midpoint of the diaphysis adjacent to the perichondrium, as defined by light microscopy and staining with collagen XII antibodies (Figures 7J and 7K). Before this time the diaphysis contains only flattened proliferative cells oriented perpendicularly to the long axis of the bone (Figures 7A-7C). The expression of p6b also coincides with a change in the appearance of the perichondrium, which shows diffuse staining with collagen XII antibodies at E13.5 and a more striated fibrillar-like staining at E14.
In the development of the long bone, both the cartilage and the perichondrium carry out programs of coordinated differentiation, starting at midpoint of the diaphysis, where the first rounded cells appear, and spreading distally in both directions in a progressive fashion (Rooney and Archer 1992; Bianco et al. 1998). Therefore, later events, such as chondrocyte hypertrophy, formation of the bony collar between the cartilage and the perichondrium, initiation of vascular invasion, and endochondral ossification, all originate in this zone. It is tempting to speculate from the unique localization of p6b that it is involved in the complex developmental crosstalk between the signals of the perichondrium and associated chondrocytes (Chazaud et al. 1996; Lanske et al. 1996; Vortkamp et al. 1996; Serra et al. 1997; von Schroeder and Heersche 1998; St-Jacques et al. 1999). For example, indian hedgehog is produced by these early prehypertrophic cells and its receptor, patched, and downstream response gene, gli3, are produced in the perichondrium in an Ihh-dependent fashion (St-Jacques et al. 1999). It has recently been shown that this Ihh signaling is required for differentiation of the osteoblasts that form the bony collar in the perichondrium. The localization of the p6b peptide to the surface of the collagen fibrils indicates that these peptides would be available for interaction with signaling molecules, such as Ihh, traversing the intercellular space to the perichondrium. Because carpal bones do not express the p6b isoform, it would be interesting to know whether their development is under the control of Ihh.
Alternatively, p6b peptide could mediate interactions between the collagen fibrils and other cartilage matrix macromolecules to strengthen the diaphysis. Early fetal cartilage is highly cellular and lacks the rigidity typically associated with mature cartilage. The highly extended shape of the diaphysis would make it prone to distortion by mechanical stress due to contraction of developing muscle or pressure from position within the uterus. In concert with the mechanical constriction and support of the perichondrium, stabilizing interactions mediated by p6b peptide could enhance the stiffness of the diaphysis. Whether for a structural or a regulatory purpose, p6b and p8 peptides are localized and available at the surface of the collagen fibrils.
The implication of the differential distribution of α1(XI) isoforms in developing cartilage is that there must be a mechanism for cell-specific alternative splicing. This likely involves
Footnotes
Acknowledgements
Supported by grants from the Shriners Hospital for Children (NPM) and the Arthritis Foundation (JTO).
We wish to acknowledge Jay Gambee for the preparation of synthetic peptides, Sara Tufa for technical assistance with electron microscopy, and Dr Kate Gregory for critical reading of the manuscript.