
Abstract
Staphylococcus aureus is a common human pathogen involved in non-bronchial diseases and in genetic and acquired bronchial diseases. In this study, we applied an immunolabeling approach for in vivo and in vitro detection of S. aureus, based on the affinity of staphylococcal protein A (SpA) for the Fc region of immunoglobulins, especially IgG. Most strains of S. aureus, including clinical strains, can be detected with this labeling technique. The approach can be used for detection and localization with transmission electron microscopy or light-fluorescence microscopy of S. aureus in infected tissues such as human bronchial tissue from cystic fibrosis (CF) patients. The methodology can also be applied to cell culture models with the aim of characterizing bacterial adherence to epithelial cells in backscattered electron imaging with scanning electron microscopy. Application to the study of S. aureus adherence to airway epithelium showed that the bacteria did not adhere in vivo to intact airway epithelium. In contrast, bacteria adhered to the basolateral plasma membrane of columnar cells, to basal cells, to the basement membrane and were identified beneath the lamina propria when the epithelium was injured and remodeled, or in vitro when the epithelial cells were dedifferentiated.
Staphylococcus aureus is a common human pathogen associated with various infections, particularly those involving the respiratory tract. S. aureus is the first pathogen to appear in cystic fibrosis (CF) respiratory infection and is also involved in acquired respiratory diseases (e.g., chronic bronchitis and nosocomial infections). S. aureus binds to many host matrix, plasma, and tissue proteins, such as fibronectin (Vercellotti et al. 1984), laminin (Lopes et al. 1985), collagen (Switalski et al. 1989), elastin (Park et al. 1991), and fibrinogen (Cheung and Fischetti 1990). For almost all of these components, a corresponding bacterial adhesin has been identified. These bacteria-binding proteins mediate the adherence of S. aureus, and it has been suggested that they can contribute to the colonization of host tissues in a number of human pathologies, such as endocarditis (fibronectin; Kuypers and Proctor 1989) and septic arthritis (collagen; Patti et al. 1994b; Foster and McDevitt 1994). However, cellular receptors for S. aureus binding to airway epithelium have yet to be identified.
Krivan et al. (1988) showed that purified asialylated gangliosides (asialo GM1) could bind S. aureus. Recently, Ulrich et al. (1998) showed that S. aureus adheres primarily to the mucous components of CF and non-CF airway epithelium, and that this bacterium does not bind to mucous-depleted ciliated cells. However, areas of CF tissues examined by these authors did not exhibit any sign of remodeling or damaged epithelial structures, as usually observed in CF (Brézillon et al. 1997). In bronchial obstructive diseases (CF or non-CF), bacteria not only produce virulence factors that affect mucous synthesis and ciliary beating but also induce airway epithelial damage. This injured and repairing epithelium may then offer new sites of adherence for opportunistic pathogens (Plotkowski et al. 1991). However, the adherence of S. aureus to airway epithelium, in relation to its remodeling state, has never been previously investigated. Because the in vivo or in vitro study of S. aureus adherence to airway epithelium requires immunocytochemical identification of the bacteria, the present study was conducted with the aim of developing a suitable immunolabeling approach that could be applied to the detection of S. aureus in CF infected lung tissues and in cell culture models. This CF tissue provides a prime example of in vivo S. aureus respiratory infection. The methodology may be applicable to either immunofluorescence observation with light microscopy or immunogold detection with scanning electron microscopy in backscattered electron imaging or transmission electron microscopy in tissues or cell cultures.
Materials and Methods
Human Airway Tissues
Chronically infected lungs were obtained from six CF patients (five males and one female; four homozygous for the AF508 mutation, two with unknown genotypes; mean age 23.5 ± 10.3 years) before surgical lung transplantation. Sputum bacteriology showed that these CF patients were chronically colonized by S. aureus or P. aeruginosa isolated as a unique infecting pathogen. Tissue fragments were taken at lobar and segmental levels and were cryofixed in optimal cutting temperature (OCT) medium (Tissue Tek; Miles, Elkhart, IN).
Human Airway Epithelial Cell Culture
Nasal polyps from non-CF patients were used as human airway tissue for the cell culture study in 24-well plates. The polyps were cut into small explants (2 mm2) and seeded onto a Type I collagen matrix in a defined RPMI 1640 culture medium (Seromed Polylabo; Strasbourg, France) supplemented with insulin (1 μg/ml; Sigma Chemical, St Louis, MO), apo-transferrin (1 μg/ml; Serva, Heidelberg, Germany), epidermal growth factor (10 ng/ml; Serva), hydrocortisone (0.5 μg/ml; Sigma), retinoic acid (10 ng/ml; Sigma), penicillin (100 U/ml) and streptomycin (100 μg/ml; Sigma) at 37C in air with 5% CO2 (Chevillard et al. 1991). After 3–4 days of culture, explants were surrounded by an outgrowth resulting from both cell migration and proliferation. Well-differentiated and non-differentiated airway epithelial cells (in the center and at the periphery of the outgrowth, respectively) could be identified.
Bacteria
The Cowan III strain (ATCC, Rockville, MD; #12600) and the protein A (SpA)-deficient strain of S. aureus (DU5875 strain; a generous gift from Pr. T.J. Foster, Department of Microbiology, Trinity College, Dublin, Ireland) were maintained at −20C in trypticase soy broth (TSB; Institut Pasteur, Paris, France) containing 20% v/v glycerol. For adherence assays, bacteria were cultured overnight in TSB at 37C, harvested by centrifugation, and resuspended in RPMI 1640 culture medium containing 20 mM HEPES (N-2-hydroxyethylpiperazine-N′-2-ethanesulfonic acid). The bacterial suspension was adjusted to a final concentration of 107 colony forming units (CFU)/ml. Bacterial self-aggregation was limited by extruding ex tempo the bacterial suspension through a fine needle just before contact with the airway epithelial cell cultures.
Bacterial Adherence
On Day 3 or 4, antibiotics were removed from primary human airway epithelial cell cultures by repeated rinsing with RPMI 1640 culture medium. A 200-μl aliquot of bacterial suspension was added to the cultures for 1 hr at 37C. To remove nonadherent bacteria, cultures were rinsed three times with 0.5 ml phosphate-buffered saline (PBS; 0.1 M, pH 7.2) and were prepared for immunofluorescence microscopy or for scanning or transmission electron microscopy.
Immunodetection of S. aureus
The immunodetection of S. aureus was based on the high affinity of SpA, a cell wall-associated protein of S. aureus, for Type G immunoglobulins (IgG) through binding to the Fc fragment (Forsgren and Sjöquist 1966). The immunolabeling procedure for S. aureus is shown in Figure 1.
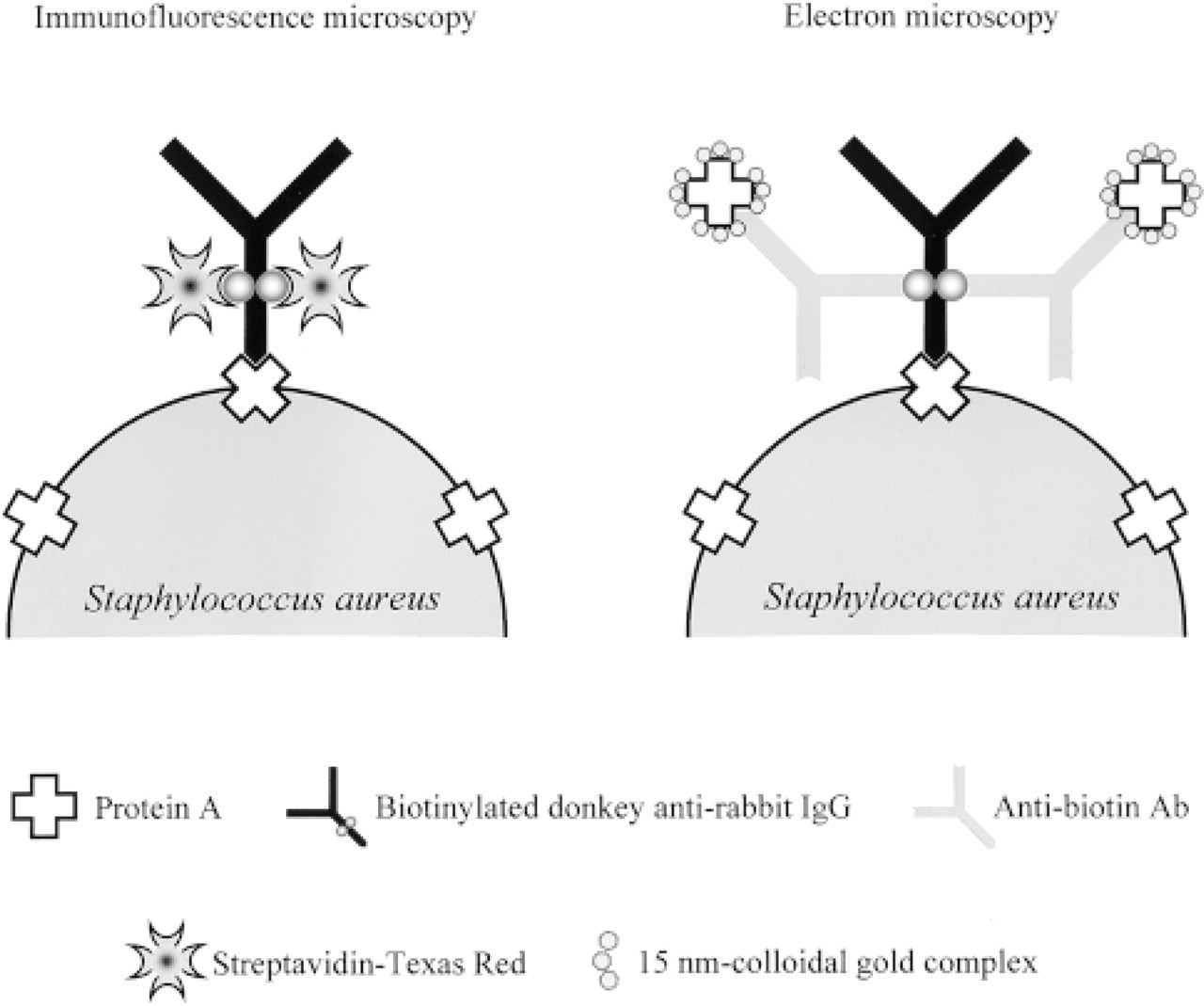
Principle of the succesive steps used for the immunological detection of S. aureus in light and in electron microscopy. The labeling approach is based on the detection of the most characteristic protein produced by S. aureus, SpA, a cell wall-associated protein, which binds in a non-antigenic but specific way to the Fc region of IgG. For the immunofluorescence protocol, cell wall-associated SpA molecules were detected with biotinylated donkey anti-rabbit IgG and then with streptavidin-Texas Red. For the immunogold protocol, cell wall-associated SpA molecules were detected with biotinylated donkey anti-rabbit IgG, with a goat anti-biotin antibody, and then with SpA-15-nm gold.
Immunofluorescent detection of S. aureus. For immunofluorescence, cultures and tissues were embedded in OCT, immersed in liquid nitrogen, and stored at −80C until cryosectioning. Five μm-thick cryosections deposited on gelatincoated slides were fixed in pre-cooled methanol (-20C) for 10 min, rinsed in PBS, and preincubated with 3% PBS-bovine serum albumin (BSA). Sections were first exposed to biotinylated donkey anti-rabbit (DAR) IgG (1:50 in 1% PBS-BSA; Amersham, Les Ulis, France) for 60 min. After incubation with 3% PBS-BSA, SpA-biotinylated IgG complexes were detected by streptavidin-Texas Red (1:50 in 1% PBS-BSA for 60 min; Amersham).
When a double immunolabeling (detection of both bacteria and epithelial antigens) protocol was performed after the labeling of bacteria as described above, sections were further incubated for 60 min with pure nonimmune goat serum (NGS; Sigma) reconstituted in distilled water according to the manufacturer's recommendations. This step saturated SpA molecules present at the surface of the bacteria before the second immunolabeling step. Sections were then incubated with primary mouse monoclonal antibody (MAb) against cytokeratin-13 (CK13; 1:600 in 1% PBS-BSA for 60 min; Sigma), a typical airway epithelial basal cell marker (Dupuit et al. 1995) and then with anti-mouse F(ab′)2 fractions coupled with digoxigenin (1:50 in 1% PBS-BSA for 60 min; Boehringer Mannheim Biochemica, Mannheim, Germany). Sections were finally incubated with anti-digoxigenin Fab fractions coupled with fluorescein isothyocyanate (FITC; 1:50 in 1% PBS-BSA for 30 min; Boehringer Mannheim). Negative controls for S. aureus detection were performed by (a) incubating the sections with nonbiotinylated DAR IgG (1:50 in 1% PBS-BSA for 1 hr; Beckman Coulter, Villepinte, France) in replacement of the biotinylated DAR IgG, to streptavidin-Texas Red and to NGS, or (b) by first exposing the sections to NGS before the labeling of the bacteria with biotinylated DAR IgG and streptavidin-Texas Red. In the second control step (negative for cellular antigen detection), the primary antibody was replaced by murine nonimmune IgG2aK fractions (Sigma). To control the specificity of S. aureus labeling, CF bronchial tissues from a patient infected only with P. aeruginosa were processed as followed. Sections were first incubated with biotinylated sheep anti-mouse IgG (1:50 in 1% PBS-BSA for 1 hr; Boehringer Mannheim), with streptavidin-Texas Red and with NGS as described above. Sections were secondly treated with a mixture of P. aeruginosa rabbit anti-sera (PMA, PMC, PME, and PMF; 1:10 in PBS-BSA 1% for 1 hr; Sanofi Diagnostics Pasteur, Marnes-La Coquette, France), with antirabbit F(ab′)2 fractions coupled with digoxigenin (1:50 in PBS-BSA 1% for 1 hr; Boehringer Mannheim), and then with anti-digoxigenin Fab fraction coupled with FITC (1:50 in PBS-BSA 1% for 30 min).
The sections were finally counterstained with Harris hematoxylin, mounted in Citifluor antifading solution (Agar Scientific; Stansted, UK), and observed with an Axiophot microscope (Zeiss; Oberkochen, Germany) with epifluorescence and Nomarski differential interference illumination.
Immunoelectron Microscopic Detection of S. aureus. For scanning electron microscopy (SEM) or transmission electron microscopy (TEM), cell cultures were fixed in 4% PBS-paraformaldehyde for 60 min at 4C. The cultures were then rinsed twice in PBS and in 1% PBS-BSA. Specimens were first incubated with biotinylated DAR IgG (1:50 in 1% PBS-BSA for 60 min) and then, to saturate SpA molecules present at the surface of bacteria, with pure NGS (for 60 min). Specimens were finally exposed to goat anti-biotin antibody (1:200 in 1% PBS-BSA for 60 min; Sigma) and to SpA-15-nm or SpA-40-nm gold particle complexes (for 60 min) prepared as previously described by Frens (1973), and postfixed overnight in 2.5% PBS-glutaraldehyde.
Negative controls were performed by first incubating specimens with pure NGS (for 60 min) or with non-biotinylated DAR IgG before incubation with goat anti-biotin antibody and SpA-15-nm or −40-nm gold particle complexes. Free SpA (Sigma; ref. P3838) used for gold particle complexes had a binding capacity of 7–14 mg of human IgG per mg solid.
For SEM, cultures were dehydrated through graded ethanol concentrations, critical point-dried with CO2, affixed onto stubs, coated with carbon (20 nm), and observed with a Philips XL30 scanning electron microscope operating at 10 kV with secondary electron imaging (SEI). In addition, immunogold-stained bacteria were identified with backscattered electron imaging (BEI) at 25 kV. Secondary and back-scattered electron signals could also be mixed.
For TEM, the cultures were dehydrated through graded concentrations of ethanol and embedded in agar resin 100 (Agar Scientific; Orsay, France). Ultrathin sections were stained with uranyl acetate and lead citrate and then observed with a Hitachi 300 transmission electron microscope operating at 75 kV.
Results
S. aureus Immunodetection with Light Microscopy
S. aureus that had adhered to the apex of airway epithelial cells or was present at the contact of basolateral membranes of airway epithelial cells could be easily detected in cell cultures (Figure 2A). Labeling of S. aureus, in parallel with the detection of basal cells using anti-CK13 antibody, allowed specific detection of all bacteria in the absence of any fluorescent background (Figure 2B). Negative control slides for S. aureus detection, using non-biotinylated donkey antirabbit IgG (Figures 2C and 2D), as well as double control experiments negative for S. aureus (preincubation with NGS) and epithelial cell marker immunodetection, showed no immunolabeling (Figures 2E and 2F). No Texas Red labeling was observed when immunodetection of S. aureus was performed on cell cultures exposed to the SpA-deficient strain (Figures 2G and 2H).
In all infected bronchial tissues from five different S. aureus-infected CF patients (Figure 3), bacteria could be detected in the sputum covering pseudostratified bronchial epithelial areas (Figures 3A and 3B). When the epithelium was pseudostratified (Figures 3C and 3D), S. aureus was detected on the basal side of the CK13 basal epithelial cells, beneath the basement membrane, and in the lamina propria. This shows that the bacteria had gained access to these sites of adherence after epithelial barrier integrity breakdown. In patchy areas of injury, bacteria were found adhered to CK13-positive airway epithelial cells, identified as basal cells, and beneath the basement membrane in the lamina propria (Figures 3E and 3F). No FITC or Texas Red labeling was observed in negative controls for S. aureus or for CK13 basal cell immunodetection (not shown). In P. aeruginosa-infected CF bronchi, the specificity of S. aureus labeling was assessed by double labeling for P. aeruginosa (in green with FITC) and S. aureus (in red with Texas Red). Using double exposure (Figures 3G and 3H), only P. aeruginosa was detected.
S. aureus Immunodetection with Electron Microscopy
In TEM, pre-embedding immunogold labeling of S. aureus was found over the entire surface of the bacteria (Figure 4). Many 15-nm colloidal particles were present at the surface of the bacteria that adhered to airway epithelial cells (Figure 4A) and to the collagen matrix on which cells had grown (Figure 4B). In contrast, bacteria were sparsely labeled with SpA-40-nm gold particle complexes (Figure 4C). Very few 40-nm gold particles were present at the surface of the bacteria, and some bacteria were not labeled (Figure 4D). In control experiments performed with preincubation with NGS before incubation with goat anti-biotin antibody and SpA-15-nm gold complex, no gold particles could be detected on bacteria (Figure 4E). Similarly, when immunodetection of S. aureus was performed in cell cultures exposed to the SpA-deficient strain, no bacteria were labeled with SpA-15-nm colloidal gold particles (Figure 4F). This pre-embedding labeling did not generate any background, inclusive of the collagen matrix or in contact with cellular membranes.
For the SEM experiments, the efficiency of labeling, characterized by the superimposition of the SEI and BEI views of bacteria, was analyzed as a function of the diameter of gold particles. Whereas the entire surface of the bacteria observed with SEI was labeled with SpA-15-nm colloidal gold particles (Figures 5A and 5B), labeling with BEI was heterogeneous, of low intensity, and with a speckled distribution of SpA-40-nm colloidal gold particles (Figures 5C and 5D). Apart from the gold particle diameter, optimal dilutions of IgG and anti-biotin antibody were adjusted to generate the best signal:noise ratio, which was characterized by the superimposition of bacteria observed with SEI with the respective labeled surface observed with BEI.
S. aureus Adherence to Airway Epithelial Cells
We further investigated, using SEM, the adherence of S. aureus to airway epithelial cells in vitro as a function of their state of differentiation. This could be easily reproduced by using the primary explant-outgrowth culture model (Chevillard et al. 1991), which permitted an analysis of the adherence of S. aureus to the native airway tissue (the explant) and to airway epithelial cells with different degrees of differentiation (the outgrowth). Whereas no S. aureus was found to adhere to differentiated ciliated airway epithelial cells in the absence of mucus, S. aureus adhered to mucous filaments (Figure 6A) present at the surface of airway epithelial cells in the outgrowth. At the periphery of the outgrowth, S. aureus adhered mainly to poorly differentiated airway epithelial cells and to the Type I collagen matrix onto which explants had been seeded (Figure 6B). These observations using SEM illustrated the difficulty in identifying isolated S. aureus from mucous granules (Figures 6A and 6B) present in the cell cultures, and underscored the necessity of coupling classical observations with the aid of SEI for immunodetection of S. aureus with BEI.
Imaging of bacteria adhered to airway epithelial cells in cell cultures was performed with the SEI and BEI configurations of the SEM. Under standard observation with SEI, the segregation of bacteria from mucous granules or microdebris present in the preparations was difficult, especially for isolated bacteria (Figure 6C). In contrast, labeled isolated bacteria were easily distinguished from mucous secretory granules and from other objects (Figure 6D).
Discussion
In the present work, an immunolabeling methodology was applied to the detection of S. aureus. The immunolabeling methodology involved is applicable to in vivo and in vitro study of the interaction of S. aureus with various tissues and particularly with airway epithelium. This method permits the immunodetection of the bacteria with optical and electron microscopic techniques in infected human bronchial tissues and in primary cell cultures.
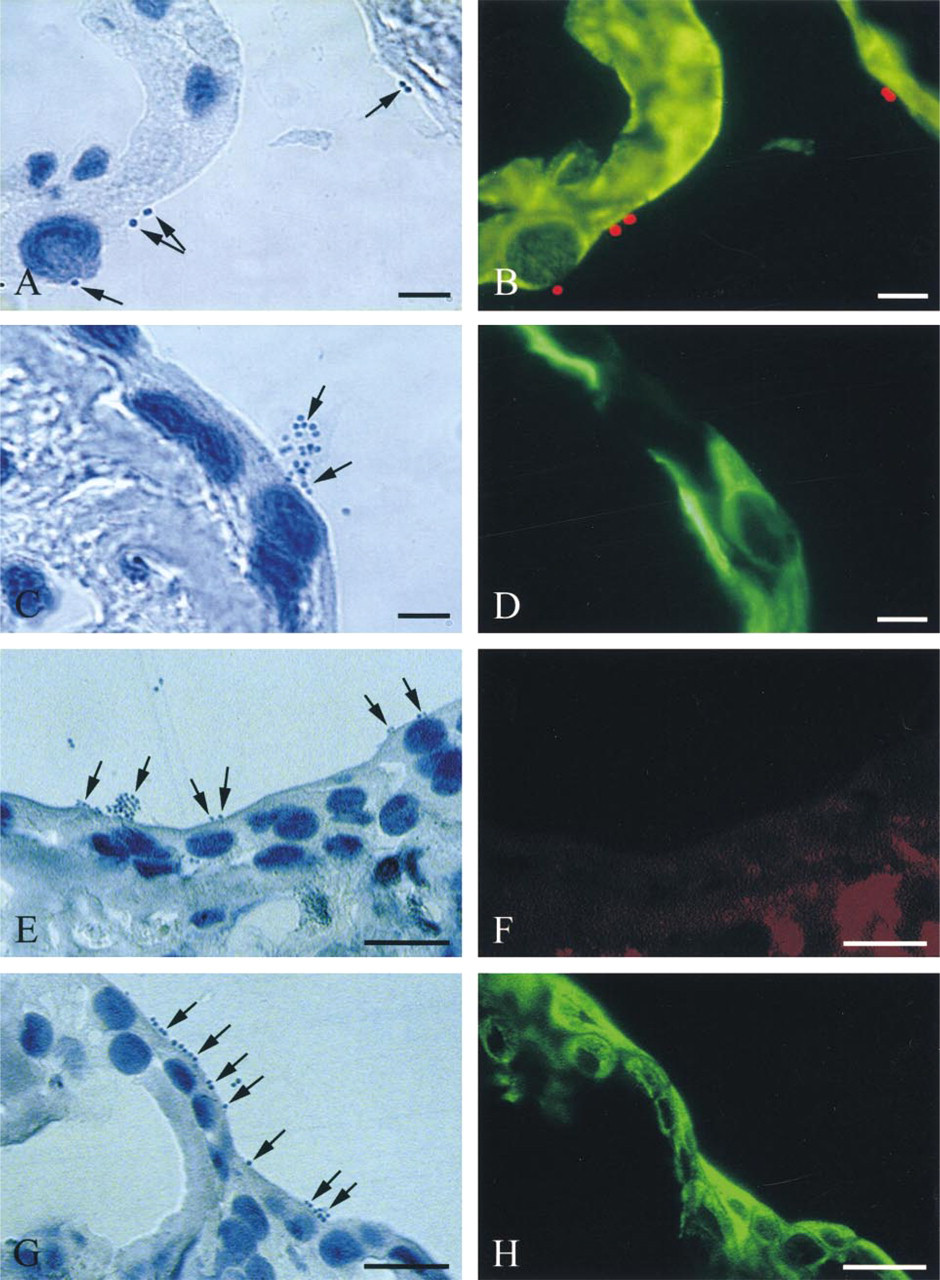
In vitro S. aureus immunodetection using light microscopy. Cowan III (
The labeling approach is based on the detection of the most characteristic protein produced by S. aureus, i.e., protein A (SpA). SpA is a cell wall-associated protein that binds in a nonantigenic but specific way to the Fc region of IgG from several species (Forsgren and Forsum 1970). SpA displays a high affinity for IgG but also, to a lesser extent, for IgA, IgE, and IgM molecules (Goding 1978). This 42-kD protein is produced by most strains of S. aureus (Seki et al. 1998) and is either covalently attached to the cell wall of the bacteria or secreted. Typical SpA consists of five IgG binding domains whose amino acid sequences are slightly different (Uhlen et al. 1984). SpA has been shown to display a great heterogeneity in the sequence and number of IgG binding domains (Sakurada et al. 1994).
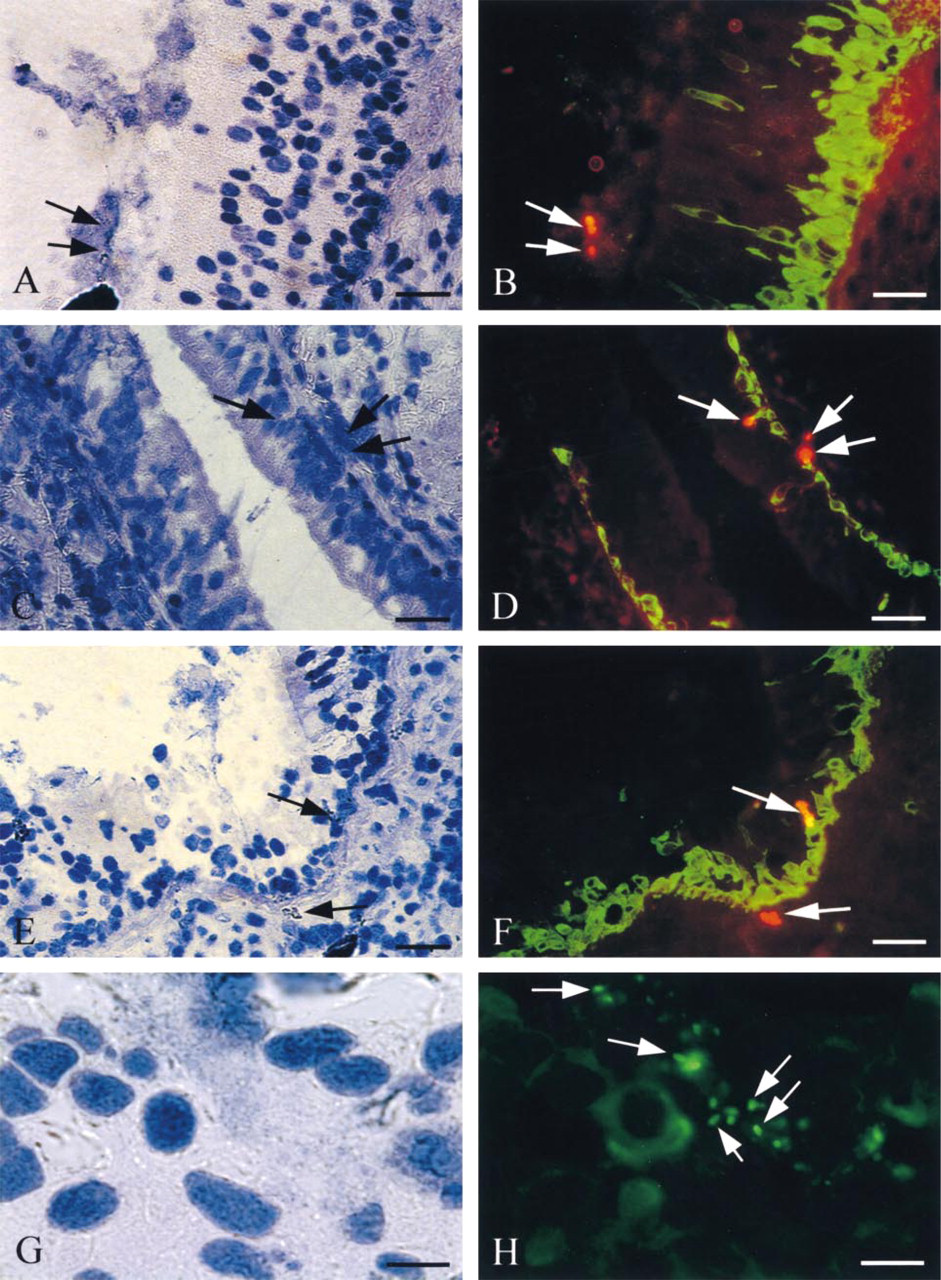
In vivo S. aureus immunodetection using light microscopy. Whereas bacteria were not easily identified in CF tissues due to the presence of mucous plugs, with the SpA labeling approach, S. aureus was easily detected in vivo in the bronchi of chronically infected CF patients, in parallel with CK13, a basal cell marker. S. aureus was observed in the mucous layer present at the surface of the pseudostratifed epithelium (
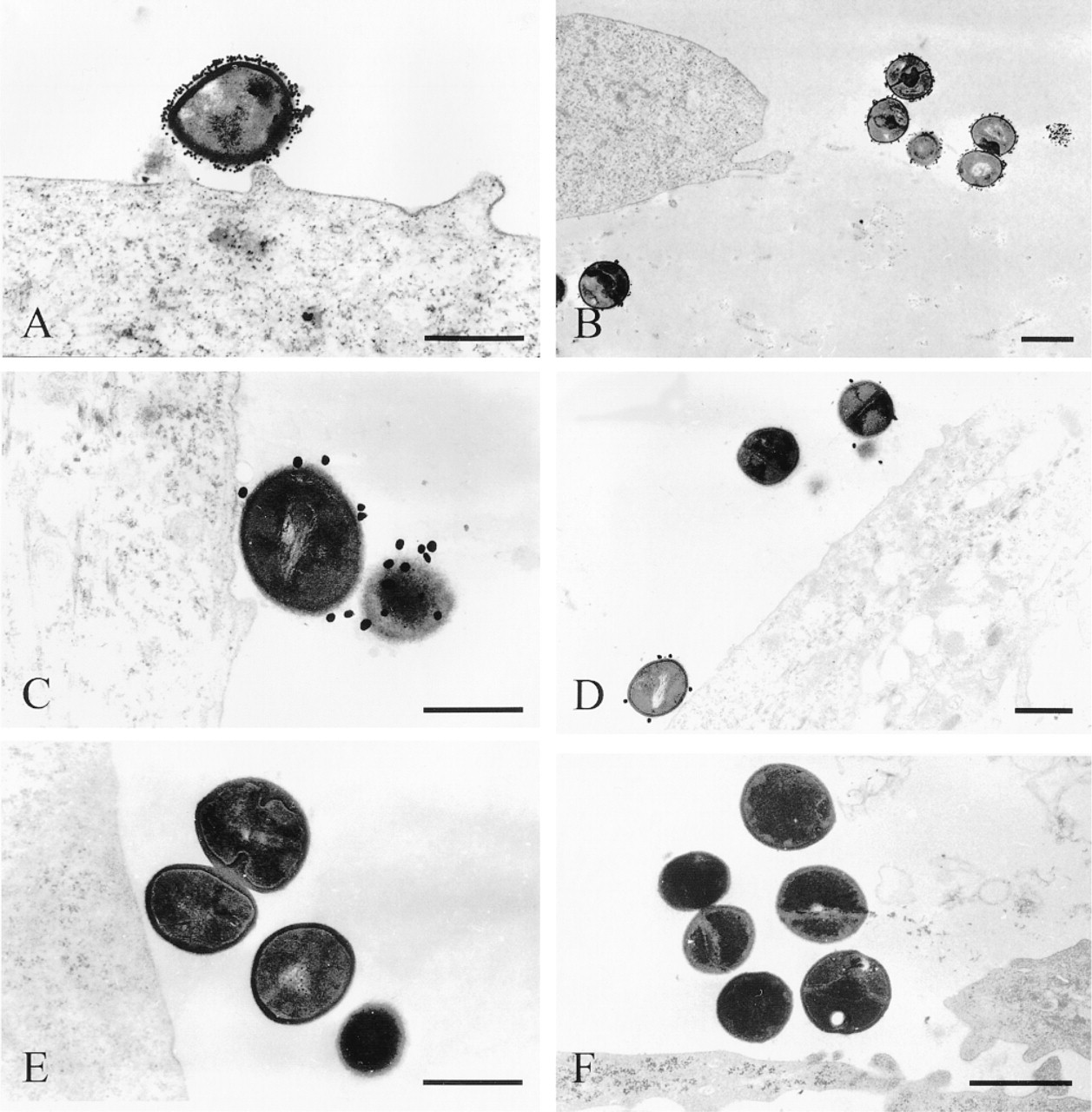
S. aureus immunodetection in TEM. Immunogold labeling of S. aureus with SpA-15-nm gold particle complexes is distributed over the entire surface of the cocci, whether the bacteria adhered to the surface of epithelial cells (
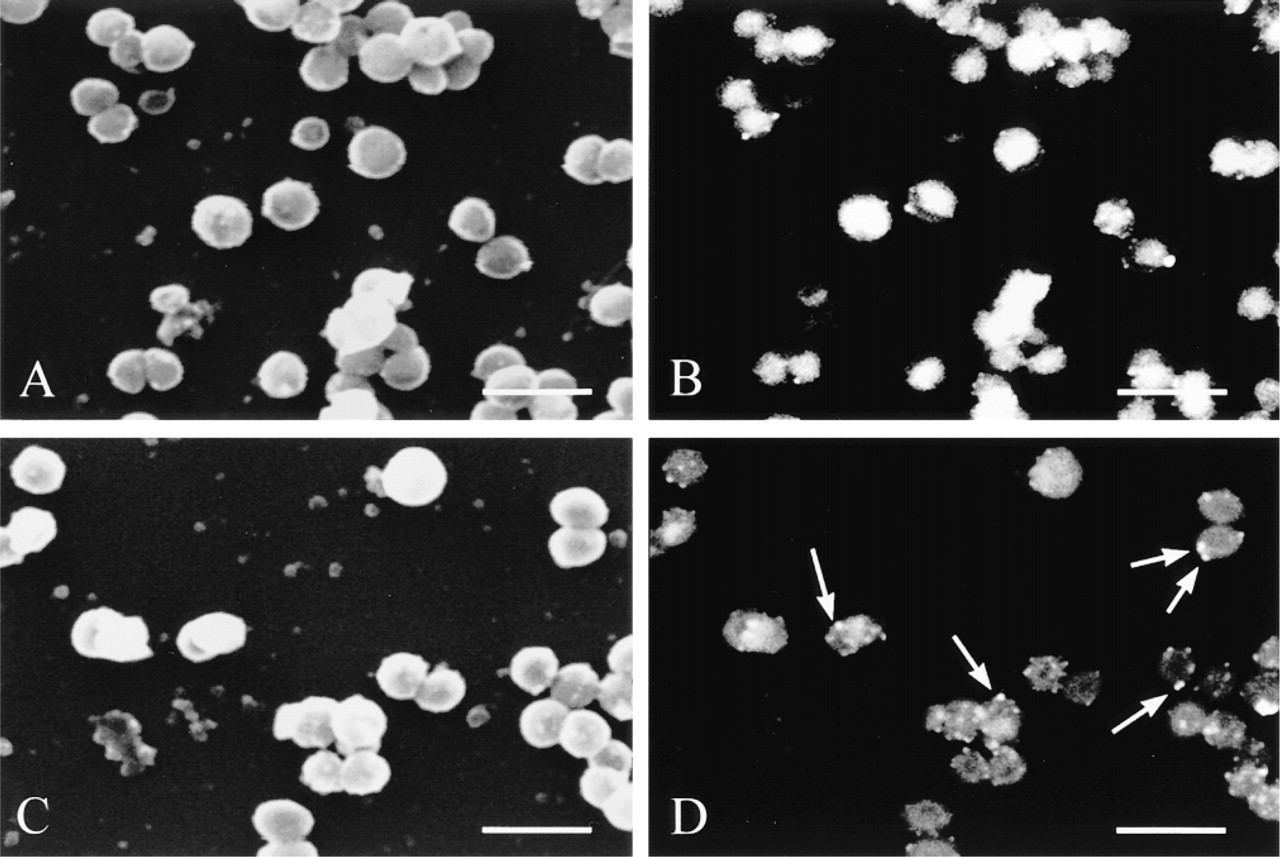
S. aureus immunodetection in scanning electron microscopy: comparison between 15-nm and 40-nm gold particles. Immunogold labeling of S. aureus in SEM is more homogeneous and intensive using 15-nm gold particles (
Because of the specific interaction of SpA with IgG, the signal detection reported here is highly specific to S. aureus. Moreover, clinical strains devoid of protein A have been shown to be uncommon, because it has been reported that among 190 clinical strains of S. aureus, only one strain was lacking the spa gene (Seki et al. 1998). Another study (Van Belkum et al. 1997) reported that, even though 85% of clinical strains of S. aureus isolated from patients with nasal colonization expressed SpA, the SpA-deficient strains were not less virulent than the SpA-producing strains. In addition, S. aureus IgG binding activity has been shown to be mediated not only by SpA but also by a polypeptide Sbi, which is clearly distinguishable from SpA (Zhang et al. 1998). The sequence homology between spa and sbi genes is limited to the IgG binding activity-encoding domain(s). No homology has been found within the promoter sequences, suggesting that the genes are regulated in different ways. This suggests that S. aureus strains that are unable to bind immunoglobulins are rare, especially in vivo where IgG binding is important to escape the immune system of the host (Patti et al. 1994a). The labeling methodology described here may permit detection of almost all the clinical strains that infect patients.
Because SpA (and, to a lesser extent, Sbi) binds not only IgG but also IgA, IgE, and IgM molecules, the difficulty in the present experiments was to saturate all SpA molecules before detection of other antigens in the preparations. This was overcome by incubating specimens first with biotinylated IgG and second with nonimmune goat serum. This procedure ensures that vacant SpA molecules present at the surface of bacteria, and possibly incompletely saturated during the immunodetection of S. aureus, do not trap immunoglobulins that will be further used for the immunolabeling of epithelial cell markers in the double labeling procedure. The labeling methodology applied in the present study is different from that used by Ulrich et al. (1998), who labeled bacteria with antibodies raised against one component of the S. aureus cell wall (techoic acid). Although this latter method is a suitable and classical approach used in the detection of S. aureus, bacteria are labeled amid a significant contaminating background signal, perhaps generated by the use of total antiserum or by the level of expression of this bacterial antigen in the bacterial cell wall. The main advantage of the immunolabeling methodology applied here is that it does not generate any background signal, even with a double immunolabeling procedure. Moreover, the amplification procedure enabled us to detect bacteria that poorly express SpA, rather than a one step-protocol based on a flagged IgG.
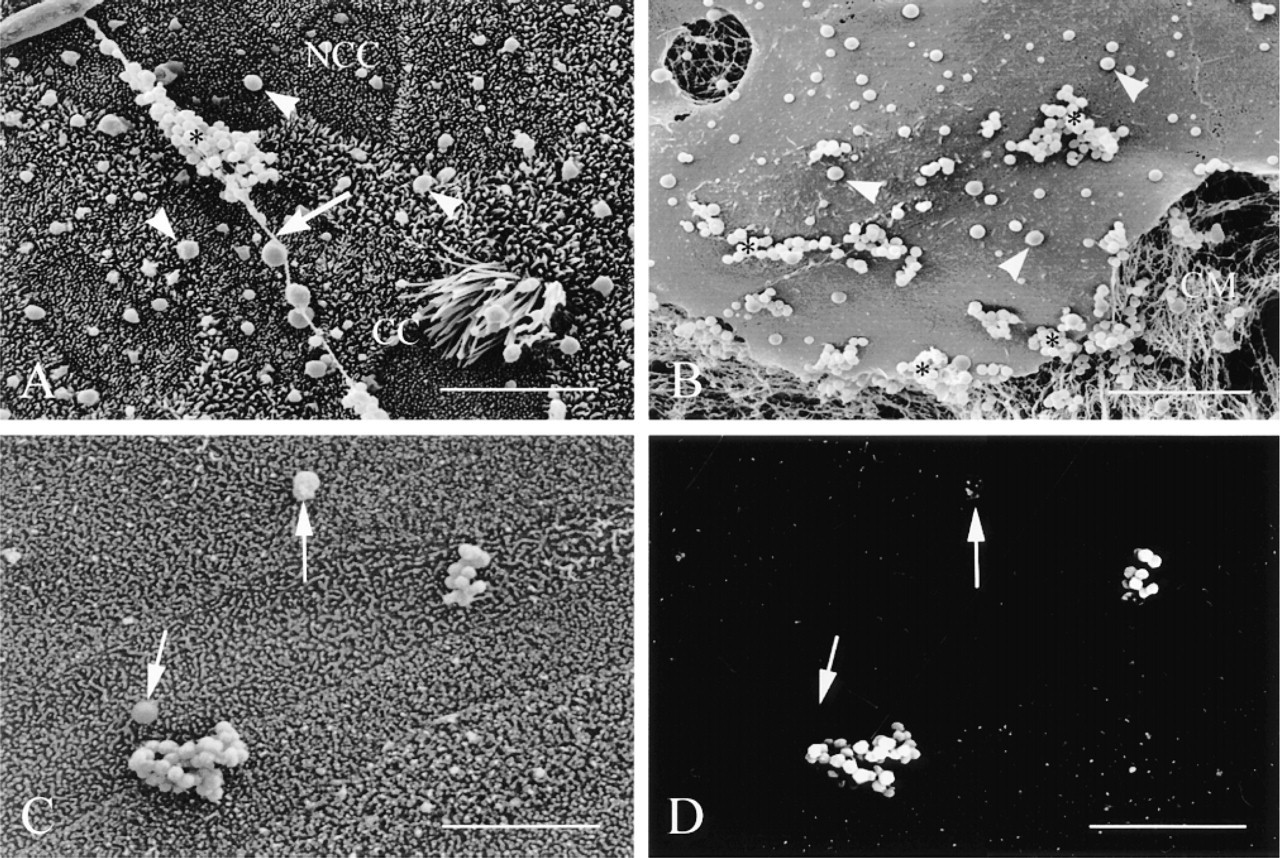
S. aureus adherence to non-CF human airway epithelium in vitro. SEM views show S. aureus (asterisks) adherent to mucous filaments (arrows) present at the surface of airway epithelial cells in the outgrowth where ciliated cells (CC) and non-ciliated cells (NCC) are present (
Using TEM, it could be shown that colloidal gold complexes were localized over the entire bacterial membrane surface, without any background signal arising in the collagen matrix or the epithelial cell membrane. To confirm that immunolabeling of S. aureus via SpA is a specific process, we also used an SpA-deficient strain of S. aureus. No labeling of the SpA-deficient strain was observed by light microscopy or electron microscopy. One could have expected a weak labeling of bacteria due to expression of the Sbi protein. However, the parent strain 8325-4 used to obtain this SpA-deficient strain has been reported to have undetectable levels of Sbi even though the sbi gene could be detected by Southern blot analysis (Zhang et al. 1998).
The identification of S. aureus with the aid of SEM is difficult because of morphological and size similarities of S. aureus with mucous granules or microdebris. The immunodetection of S. aureus with BEI coupled with SEI observations represents a useful tool for studying the mechanisms involved in S. aureus adherence to airway epithelial cells. This could be particularly helpful in the identification of bacterial adhesins and epithelial receptors using quantitative methods with computer-aided on-line image analysis techniques. We observed that immunodetection of S. aureus with BEI was optimal for SpA-15-nm gold compared to SpA-40-nm colloidal gold particles, the SpA-5-nm colloidal gold particles giving no more benefit for S. aureus detection with BEI observation (personal data). These results agree with previous reports showing that for the 15-nm gold particle size, 60 molecules of SpA are present at the surface of each gold particle (De Roe et al. 1987) and that this number is directly proportional to the average diameter of the particle. Moreover, for large gold particles (>30 nm), the gold complex becomes less stable and a less efficient probe for BEI (Frens 1972).
This cytochemical methodology based on staphylococcal protein A can be applied either in light microscopy or in electron microscopy, but also for immunodetection of the bacteria in many staphylococcal pathologies including bronchial diseases. In the present study, the application of this methodology to the detection of S. aureus in infected tissues was validated with the study of S. aureus interaction with airway epithelium. Mechanisms of colonization of airway epithelium by S. aureus remain unknown. S. aureus is capable of adhering to purified components of epithelial cell membranes, such as the binding sequence Gal-Nacβ1-4Gal present in the glycolipid asialo GM1 (Krivan et al. 1988). However, in situ studies with intact cells and bacteria are lacking. Recently, Ulrich et al. (1998) reported that S. aureus adherence to airway epithelium is negligible compared to adherence to mucus. In fact, although the CF patients in this latter study were chronically infected with S. aureus for several years, the surface airway epithelium was intact and exhibited a normal pseudostratified organization without any remodeled or injured areas. This discrepancy could be explained by the fact that our CF patients were at a late phase of disease compared to the CF patients included in the study of Ulrich et al. (1998), who were younger at the time of lobectomy. In the present work and in previous studies (Baltimore et al. 1989; de Bentzmann et al. in press), it has been shown that infected CF airway surface epithelium may exhibit intact as well as remodeled and damaged epithelial areas. The extent of each kind of epithelium, preserved or injured, varied from one pulmonary lobe to the other, but both were equally encountered. Intact pseudostratified epithelium provides a tight barrier system whose function is to prevent bacterial invasion. In intact areas of infected CF airway surface epithelium, we observed that bacteria were trapped by mucus and kept away from the epithelial cells, as previously reported by Ulrich et al. (1998). In the remodeled areas of the same tissue, we could detect bacteria at the contact of basolateral membranes of columnar airway epithelial cells in the epithelial layer and with basal cells, as well as beneath the basement membrane in the lamina propria underlying the airway epithelium. Microabscesses of S. aureus beneath the airway epithelium could represent a protective mechanism by which bacteria escape host-defense mechanisms. This tropism for injured and remodeled airway epithelium was also observed in vitro. Whereas no bacteria were found to adhere to ciliated cells in the absence of mucus, S. aureus mainly adhered to poorly differentiated epithelial cells and basal cells, as well as to basolateral membranes of columnar airway epithelial cells. This suggests that our culture model is a good model for studying bacterial adherence as a function of the remodeling and differentiation status of airway epithelium.
In the present study, we have applied an immunolabeling methodology based on the affinity of SpA for IgG, which is applicable to either immunofluorescence observation using light microscopy or immunogold detection with electron microscopic techniques for immunodetection of the bacteria in many staphylococcal pathologies, including bronchial diseases. The study of S. aureus adherence to airway epithelium, which is the first step involved in the pathogenesis of staphylococcal lung diseases, implies the identification of bacterial adhesins and epithelial receptors. This will be further achieved by using human primary airway cell culture models, with immuofluorescence and quantitative methods as well as computer-aided on-line image analysis techniques coupled with BEI and SEI in SEM.
Footnotes
Acknowledgements
Supported by a grant from DRET/DGA no. 96.34.053.00.470 and partially funded by the Association Française de Lutte contre la Mucoviscidose (AFLM). E. Mongodin is a doctoral fellow of the Ministère de la Défense - DRET/DGA.
We thank Pr T.J. Foster for providing us with the SpA-deficient strain of S. aureus. We are grateful to Pr Klossek (CHU J. Bernard; Poitiers, France) for providing human nasal polyp tissues and to Pr Couetil (Hopital Broussais; Paris, France) for CF lung tissues.