
Abstract
The Dlx-2 (distal-less gene) homeoprotein transcription factor controls early tooth development but has not been studied during the late stages of biomineralization. Transgenic mice containing a Dlx-2/LacZ reporter construct were used to map the Dlx-2 expression pattern in cementoblasts, the dental cells most closely related to bone cells and therefore suggested to be uniquely positioned osteoblasts. During initial root formation, marked expression of Dlx-2 was evident in molar and incisor root epithelium, whereas dental papilla and follicle were negative. Dlx-2 was expressed in this epithelium from the apical loop to the area of its disruption. During acellular cementum formation in both incisors and molars, Dlx-2 expression was observed in the majority of differentiated cementoblasts from the apical region to the erupting zones. During cellular cementum formation, the presence of which characterizes growth-limited molars, Dlx-2 expression was restricted to the innermost cementoblasts and entrapped cementocytes. These data further support the hypothesis of a complex origin and fate of cementum-forming cells, as previously suggested by the expression patterns of a set of mesenchymal and epithelial markers, notably ameloblastin as shown here. Dlx-2 expression might constitute a landmark of cementoblast subpopulations of epithelial origin.
R
Two types of cementum have been identified, the distribution of which varies depending on the tooth type. The acellular cementum covers the buccal aspect (root analogue) of the incisor surface and the coronal and the middle portions of the molar root surface; the cellular cementum covers the apical portion and furcation of the molar root surface. Acellular and cellular cementums differ mainly by the matrix organization of the collagen fibers and the absence/presence of cells included in the cementum matrix (Bosshardt and Schroeder 1996).
The determination of putative factors involved in the fate of root epithelial cells and the consecutive cementoblastic cell differentiation (Davideau et al. 1995; Ten Cate 1997; Terling et al. 1998) is largely based on experiments concerning tooth crown formation (for review see Thesleff and Sharpe 1997). Homeobox genes encode transcription factors that play an important role during tooth morphogenesis, as shown for Msx and Dlx genes (Thesleff and Sharpe 1997). Dlx homeobox genes constitute a subfamily of divergent homeobox genes related to the Drosophila distal-less (Dll) gene (Stock et al. 1996). In mice, six Dlx genes have been identified and their expression pattern involves distinct and overlapping regions of developing craniofacial structures, especially teeth (Qiu et al. 1997; Weiss et al. 1998). The loss of Dlx-1 and Dlx-2 gene function in null mutant mice results in profound alterations of orofacial skeletal structures, including the absence of maxillary molar development (Thomas et al. 1997). These studies have established the key role of Dlx genes in early tooth patterning (Sharpe 1995; Thomas and Sharpe 1998; Weiss et al. 1994, 1998). The role of Dlx genes in the later stages of dental tissue formation has been recently highlighted by the identification of a DLX3 mutation associated with the tricho-dento-osseous syndrome (Price et al. 1998). This syndrome is principally characterized by dental root and enamel defects (Price et al. 1998). Furthermore, the strong expression of Dlx-2 in dental epithelium during mouse amelogenesis (unpublished observations) suggests that the Dlx gene may be involved in the control of ameloblast differentiation (Thomas et al. 1995).
To investigate the potential role of Dlx gene during root formation, the expression of the Dlx-2 gene was studied in Dlx-2/LacZ transgenic mice (unpublished data).
Materials and Methods
Investigation of Dlx-2/LacZ Transgenic Mice
Transgenic mice were produced using a Dlx-2/LacZ reporter construct containing 3.7 kb upstream of the translation start site and thus using the endogenous Dlx-2 promoter (unpublished data) in B6CBA mice (Haston, UK). Crossing a single line, confirmed by Southern analysis, with wild-type mice produced heterozygotes and wild-types used in this study. Thirteen-, 21-, and 45-day-old mice were used (n =70). Mandibles of wild-type and heterozygous Dlx-2/LacZ mice were dissected in 0.1 M PBS (Sigma; La Verpillière, France), pH 7.4, and then fixed in 4% paraformaldehyde (Sigma) in PBS for 2 hr at 4C. The samples were rinsed for 2 hr in PBS at 4C and stained for β-galactosidase activity. The staining was performed overnight at 37C in PBS containing 4 mM of both ferri- and ferrocyanic acid (Sigma), 2 mM of magnesium chloride (Sigma), 0.04% of X-Gal (Euromedex; Strasbourg, France) stock at 50 μg/μl in dimethylsulfoxide (Sigma), and 0.02% of Nonidet-P40 (Sigma). The samples were again fixed for 24 hr at 4C with the same fixative described above and rinsed in PBS for 4 hr at 4C. Samples were then decalcified for 4 weeks at 4C in PBS with 4.13% disodium ethylenediamine-tetraacetic acid (Sigma) and 0.2% paraformaldehyde (Sigma), pH 7.4. Samples were dehydrated through ascending concentrations of ethanol and cleared with toluene before wax embedding. Serial paraffin sections (8 μm) were cut (Microtome Leica RM 2145; Rueil-Malmaison, France) in sagittal and frontal planes, placed on 50 mg/ml poly-
Immunohistochemistry
Immunohistochemistry with polyclonal antibodies against rat ameloblastin (gift of Paul Krebsbach, NIDCR, Bethesda, MD), was performed on 21-day-old Dlx-2/LacZ transgenic mice wax sections prepared as for histological analyses. Those polyclonal antibodies were produced against a purified recombinant ameloblastin as described in Lee et al. (1996).
Primary and secondary antibodies, as well as conjugated peroxidase, were diluted in a solution of 0.05% Tris-NaCl, 2.5% NaCl, pH 7.4, containing 1% normal goat serum (NGS; Amersham). Washes were performed in 0.05% Tris-NaCl solution, pH 7.6, containing 1% bovine serum albumin (Sigma). Endogenous peroxidase in the tissues was inactivated by treatment with 3% H2O2 for 15 min. The tissue was then blocked with 10% NGS for 24 hr at 4C. The primary antibody (1:1000 dilution) was incubated for 18 hr at 4C, biotinylated secondary antibody (1:800; Sigma) for 30 min, and streptavidin-peroxidase (1:300; Vector, Compiegne, France) for 30 min. Diaminobenzidine was used as the peroxidase substrate (Sigma). The sections were counterstained or not with eosin 0.5%, dehydrated, and mounted with DePeX resin. Micrographs were taken with a Zeiss Orthoplan microscope.
Results
In mouse mandible, two different tooth types are present, i.e., the incisor and the three molars. Incisors correspond to continuously growing organs that exhibit the entire sequence of root development throughout mouse life (Figure 1), whereas molars (Figure 2) correspond to a limited growing model.
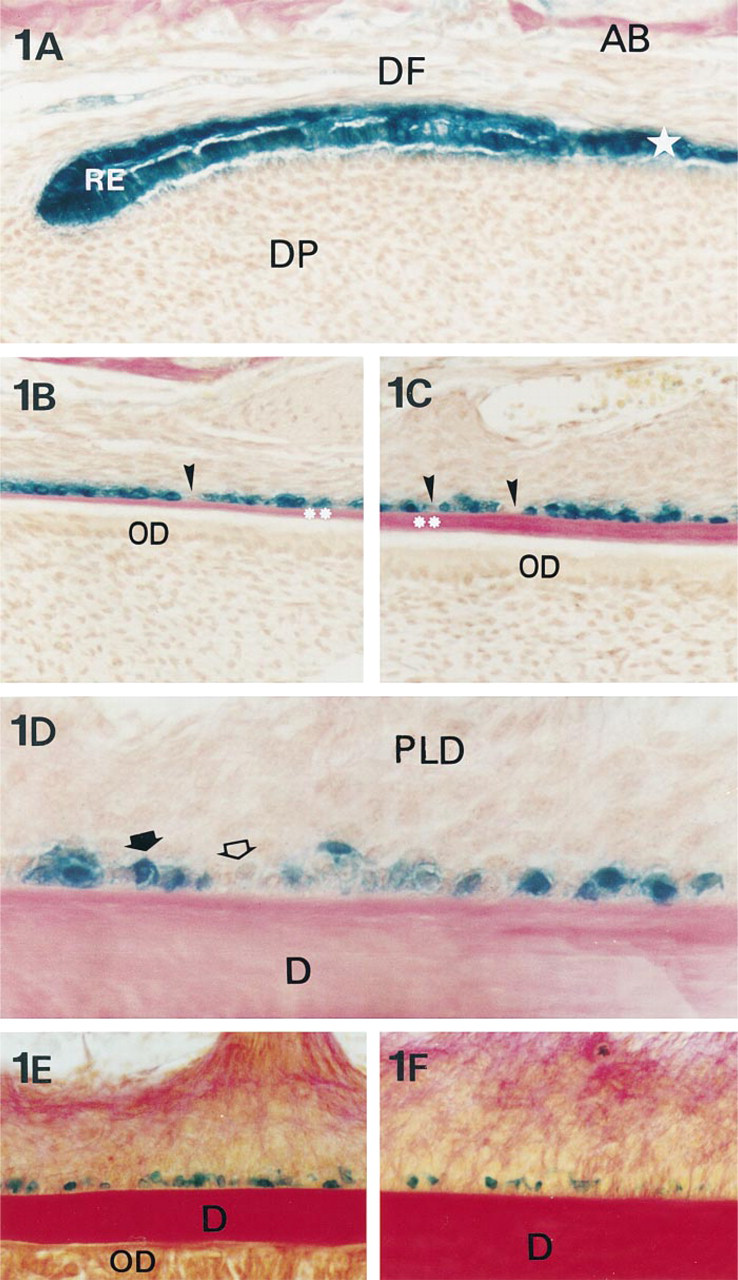
Expression of Dlx-2 in mice incisors during acellular cementum formation. β-Galactosidase histoenzymology in mandibular root incisor sections. In 13-day-old mice, from the apical (
In the “apical” part of the incisor, the epithelial root sheath is continuous and the root odontoblasts differentiate (Figure 1A). During the deposition of the first dentin layer by odontoblasts, the epithelial root sheath began to disrupt and the presumptive cementoblasts became progressively prominent along the root surface as the first layer of cementum matrix formed (Figures 1B and 1C). At the same time, the periodontal ligament fibroblasts flattened and became progressively angled to the root surface, and periodontal collagen bundles inserted in the cementum matrix (Figure 1D). From 13 to 45 days, the periodontal ligament underwent full development, becoming wider and more fibrous (Figures 1D-1F).
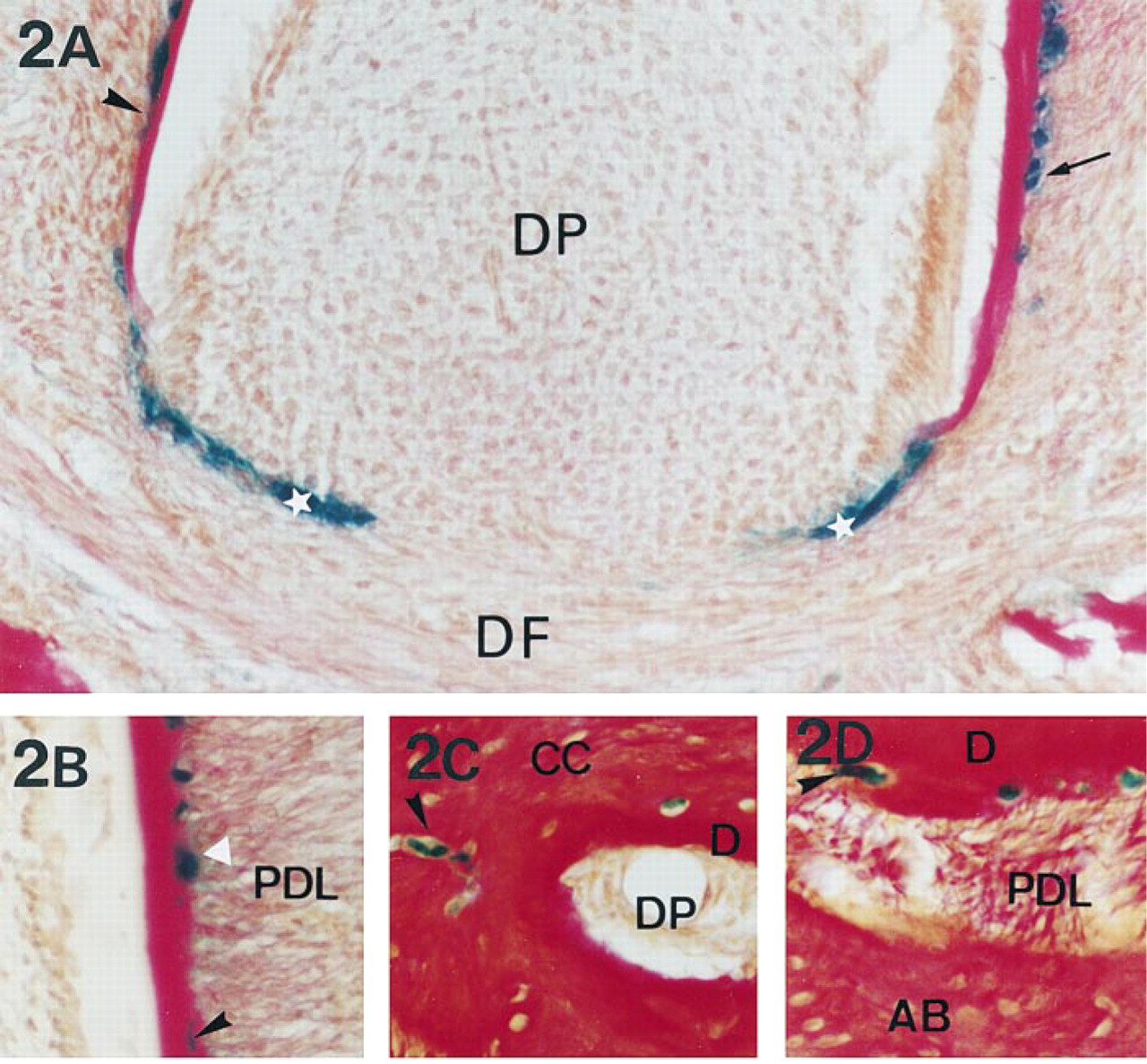
Expression of Dlx-2 in 13- to 45-day-old mouse molars during acellular and cellular cementum formation. β-Galactosidase histoenzymology in mandibular molar sections concerning the root apical region. (
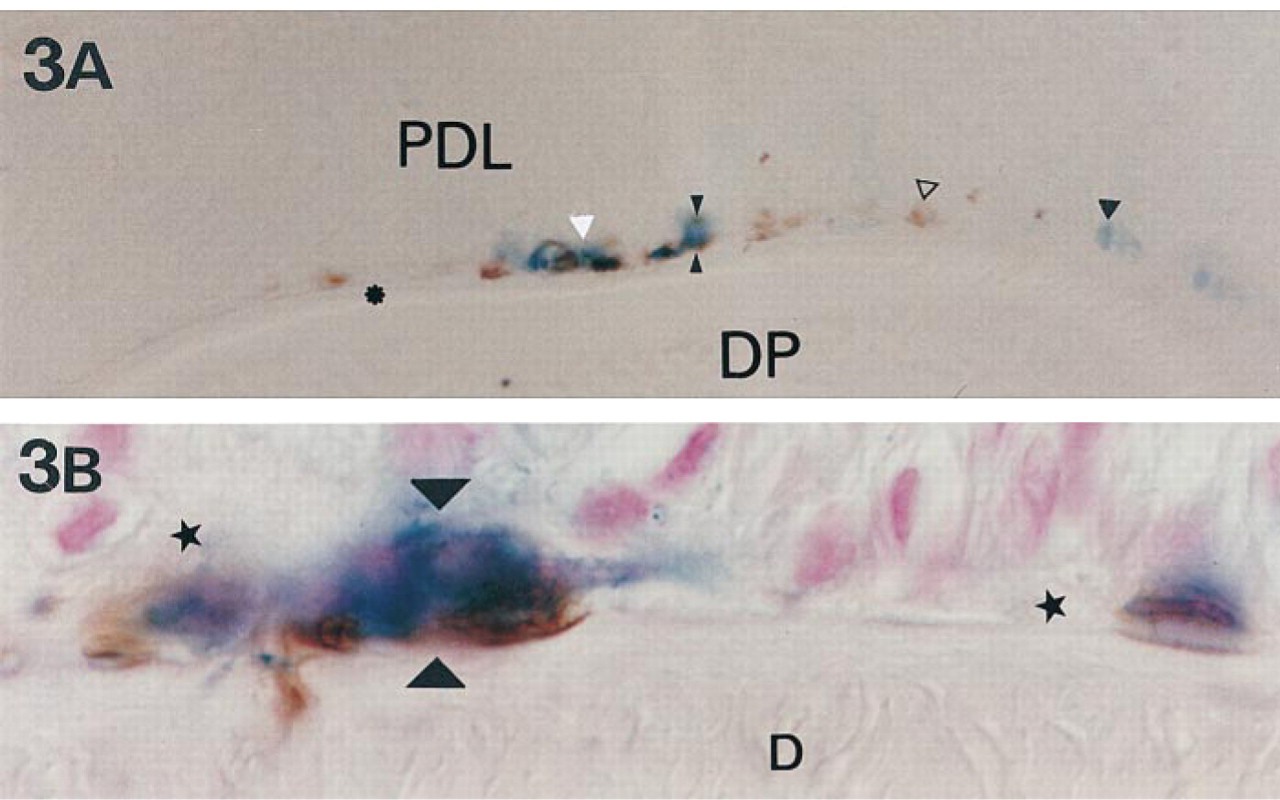
Expression of Dlx-2 and ameloblastin in 21-day-old mouse molars during acellular and cellular cementum formation. (
In incisors from 13- to 45-day-old mice, strong Dlx-2 expression was observed in the epithelial root sheath, whereas dental follicle and papilla cells were not labeled (Figure 1A). No β-galactosidase staining was observed in wild-type animals (not shown). During root sheath disruption, Dlx-2 was still expressed in epithelial cells lining the root dentin (Figure 1B), whereas unlabeled cells progressively appeared on the root surface (Figure 1C). During acellular cementum matrix deposition, strong Dlx-2 labeling was observed in the major part of the cementoblasts associated with the maturing periodontal ligament of 13-day-old mouse incisors (Figure 1D). In 45-day-old mouse incisors, Dlx-2 was expressed in cementoblasts located between the dense fiber bundles inserting in the cementum matrix (Figures 1E and 1F).
In pre-erupting first molars of 13-day-old mice, the root was progressively formed in relation to the lengthening of root sheath from the cervical loop (Figure 2A). At this stage of root formation, the cementum deposited against root dentin was of the acellular type. In erupting first molars of 21-day-old mice, two thirds of the root length was formed. At this stage, the major part of the cementum deposited was of the acellular type, but the first formation of cellular cementum was observed at the apical root portion and was characterized by the inclusion of cells in the cementum matrix (Figure 2B). In erupted first molars of 45-day-old mice, “final” root length was achieved and the root sheath had disappeared. At this stage, the formed cementum was mostly cellular. The thickness of cellular cementum progressively increased with age and was maximal at the apex (Figure 2C). Furthermore, the cellular cementum spread coronally and was deposited on previously formed acellular cementum layers (Figure 2D).
As observed in incisors, Dlx-2 was strongly expressed in the molar root sheath (Figure 2A). In preerupting and erupting molars, Dlx-2 labeling was observed in a limited number of cementoblasts lining acellular cementum (Figures 2A and 2B). Their number decreased progressively with age, as shown by the few Dlx-2-labeled cementoblasts detected on the root surface of 45-day-old mice erupting molars (Figures 2C and 2D). During cellular cementum formation, Dlx-2 expression was observed only in some cementocytes of the initial cellular cementum deposited at the root apex (Figures 2B and 2C). Dlx-2 labeling was also detected more coronally in a few cementocytes included in the first layer of cellular cementum deposited on acellular cementum (Figure 2D).
Immunolabeling of ameloblastin in transgenic mice enabled us to detect the cells that co-expressed Dlx-2 and ameloblastin (Figure 3). During the early stages of molar root sheath dislocation (Figure 3A), the cells lining the root surface were either labeled for both Dlx-2 and ameloblastin or exclusively for Dlx-2 or ameloblastin. During initial deposition of the cellular cementum above acellular cementum (Figure 3B), successive steps of cell embedding inside the cementum matrix could be analyzed. Some Dlx-2-labeled cementoblasts and cementocytes also appeared to contain ameloblastin.
Discussion
This report is the first description of the expression of a homeobox gene in dental root tissues. Our data suggest that the Dlx-2 homeobox gene may be involved in the control of cementum formation. Moreover, the continuous presence of Dlx-2 in odontogenic cells from tooth initiation (Thomas et al. 1997) and morphogenesis (Thomas et al. 1995) to the latest stages of dental tissue formation, as shown here in root tissues, illustrated the fact that the role of homeobox genes is not restricted to the early stages of tooth organogenesis.
During root formation of molars and continuously erupting incisors, Dlx-2 was strongly expressed in the epithelial root sheath from the apical loop to the disrupting area. The apical loop of crown dental epithelium is also characterized by intense expression of Dlx-2 (Thomas et al. 1995). The homologous expression in both crown and root epithelial cells has been previously shown for receptors of EGF (Davideau et al. 1995), and growth hormone (Young 1995). These data show that crown and root epithelial cells initially share some common regulatory mechanisms, as confirmed by their similar capacity to induce crown and root odontoblast differentiation (Thomas and Kollar 1989). The specific role of Dlx-2 during the differentiation of mineralized tissue-forming cells remains unknown. However, other homeobox genes, such as Dlx-5 and Msx-2, are involved in the regulation of osteocalcin gene expression of differentiating osteoblasts (Newberry et al. 1997; Ryoo et al. 1997). These data suggest that Dlx-2 may be involved in the control of root epithelial cell differentiation, as shown for neuronal cell differentiation (Ding et al. 1997).
During radicular cementum formation, cementoblasts in a similar manner to osteoblasts and odontoblasts, when overtly differentiated, are involved in matrix deposition in cooperation with periodontal ligament cells (Bronckers et al. 1994; Davideau et al. 1998). The modalities of cementum formation vary depending on tooth type. The incisor contains an acellular cementum which is continuously deposited during the eruption process. In contrast, the roots of molars contain two cementum types. The first one deposited is acellular. The second one is cellular, and progressively increases in thickness towards the root apex. During acellular cementum formation, the persistence of Dlx-2 expression in cementoblasts confirms that some of these cells display root epithelium cell characteristics. This was also shown by the presence in root-lining cells of several epithelial markers, such as E-cadherin (Terling et al. 1998), and an enamel protein, the amelin/ameloblastin/sheathlin studied at the mRNA level (Fong et al. 1996) and protein level (this study). Ameloblastin was used and clearly evidenced in this study rather than the main enamel protein amelogenin, which appeared to be detected lining the root surface only by electron microscopy (Bosshardt and Nanci 1997). This observed patchwork/lattice of Dlx-2 positive cells along the root surface may correspond to the different origins of cementoblasts: one group would derive from root epithelium and the other from the ectomesenchyme of the follicular sac (Cho and Garant 1988; Bosshardt and Schroeder 1996). The mesenchymal transition in odontogenic epithelium is supported by studies reporting the expression of some mesenchymal markers in dental epithelial cells (Bosshardt and Nanci 1997; Chen et al. 1998) and, more precisely, the expression of bone sialoprotein and osteopontin in epithelial cells lining the acellular fibrillar cementum in the cervical portion of the tooth (Bosshardt and Nanci 1997). Epithelial and ectomesenchymal root-lining cells would interact during cementum formation (Alatli et al. 1996). An alternative would be that Dlx-2 and ameloblastin down-regulation is one landmark of a progressive epithelio-mesenchymal conversion of root epithelium-derived cementoblasts (Thomas 1995), as shown by the co-expression of keratin and vimentin in cementoblasts in vivo (Webb et al. 1996) and in vitro (Thomas 1995). The present study also shows that the selective expression patterns of ameloblastin and Dlx-2 may follow divergent signaling pathways, illustrating the complexity of cell fate in the epithelial root sheath during cementogenesis. The fact that the proportion of lining cells expressing ameloblastin and/or Dlx-2 appeared limited may reflect a progressive loss of epithelial features of involved cells during the progression of root formation.
During cellular cementum formation in molars, Dlx-2 and ameloblastin were expressed only in root epithelium and in some cementocytes of the first deposited layers of cellular cementum. A similar expression pattern has been previously observed for amelin mRNA (Fong et al. 1996), suggesting that some root epithelial cells were included passively in cementum matrix (Thomas 1995; Bosshardt and Schroeder 1996). Furthermore, the absence of Dlx-2/ameloblastin expression in cementoblasts lining the more external cellular cementum layers and the major part of cementocytes suggests that the later stages of cellular cementum deposition do not involve the Dlx-2/ameloblastin epithelial pathways as occurs during acellular cementum formation. An alternative would be that acellular and cellular cementum-forming cementoblasts do not have the same origin.
The expression pattern of Dlx-2 during root formation illustrates the complex origin and fate of cementum-forming cells. Comparative analysis of the homeoprotein Dlx-2, ameloblastin, and previously published matrix protein expression patterns suggests that root morphogenesis and cementogenesis involve epithelio-mesenchymal cooperation from the initial to the later stages of dental root formation.
Footnotes
Acknowledgments
Supported by grants from the EA2380 laboratory, University Paris VII. Travel grants were also obtained from EEC (COST B8 action).