
Abstract
Fibroblasts are the principal mesenchymal cell type found within the connective tissues of all organs. Once thought to play a passive role in tissue remodeling, fibroblasts have now emerged as a key player in regulating structural immunity and modulating the reparative injury response. A recent surge in single-cell RNA sequencing studies has advanced our understanding of the biology of fibroblasts, highlighting their cellular diversity and organization across health and diseased conditions at an unprecedented resolution. In this review, we discuss up-to-date literature on fibroblast subpopulations identified from 2 distinct barrier tissues: oral mucosa and skin. We focus on the transcriptomic signatures that distinguish subsets of fibroblasts in homeostasis and perturbed conditions (i.e., wound healing or chronic inflammatory diseases), and we link them to mechanistic studies that provide functional insights. A deeper understanding of fibroblast diversity and its functional significance may uncover tissue-specific roles in regeneration and immunomodulation, which will be crucial for the development of precision therapy that directly targets fibroblast subsets.
Keywords
Introduction
Fibroblasts were first described in the 19th century as spindle-shaped cells of the connective tissue. Perhaps owing to this designation based on morphology, there had been ambiguity to whether fibroblasts should be truly classified as a single-cell type. We now know that morphology alone is inadequate and does not convey the functional significance of fibroblasts. Fibroblasts are commonly identified by the positive cell surface protein expression of pan-fibroblast markers such as CD140A and/or CD90 and the lack of leukocytic, endothelial, epithelial, and erythrocyte lineage markers (CD45-, CD31-, CD326-, Ter119-, respectively). However, the aforementioned criterion is not an absolute definition of fibroblasts, since it also identifies multipotent skeletal/mesenchymal stromal cell subsets in the long bones (Ambrosi et al. 2019). Thus, single-cell RNA sequencing (scRNA-seq) studies on fibroblasts typically select for additional extracellular matrix genes, such as Dcn, Col1a2, Col3a1, and more. Besides their role in extracellular matrix deposition, fibroblasts influence the immune landscape during wound healing, cancer, and chronic inflammatory disorders (He et al. 2020; Correa-Gallegos et al. 2021). These diverse functions of fibroblasts across and within the organs implicate the existence of fibroblast subpopulations that can directly be linked to pathologic outcomes, which is less explored in the oral biology field.
The oral cavity is subject to repeated trauma from mastication and microbial insult. Despite this perpetual injury, oral mucosae heal exceptionally fast and with minimal scar formation (Iglesias-Bartolome et al. 2018). Moreover, the damage induced by periodontal pathogens in periodontitis is typically gradual and rarely results in sepsis. The resilient nature of the oral mucosa had been attributed to the neural crest developmental origin of oral fibroblasts (Rinkevich et al. 2015). In contrast, skin fibroblasts have multiple developmental origins: paraxial mesoderm gives rise to dorsal skin fibroblasts and lateral plate mesoderm to ventral trunk and limb skin, and, uniquely, facial skin fibroblasts are derived from the neural crest. Facial fibroblasts exhibit higher proliferative capacity and contribute to an improved wound-healing profile as compared with other skin regions (Usansky et al. 2021), which is linked to the continued expression of tissue-specific homeobox genes throughout adulthood (Pfeiferová et al. 2025). These studies link the developmental origin to the niche-specific function of the fibroblasts, but the distinguishing features between oral and skin fibroblasts at a single-cell level had been underexplored. Emerging scRNA-seq studies have addressed this challenge, revealing that multiple fibroblast subpopulations are found in the oral mucosa and skin and providing novel insights into understanding their functional significance in health and disease.
Oral and cutaneous barriers closely resemble each other at the histologic level. Yet, fibroblast composition and function in these anatomic sites are expected to differ. In this review, we provide an up-to-date review on research development in the single-cell biology of fibroblasts in the oral mucosa and skin. Specifically, we discuss the gene markers that define fibroblast diversity in these barrier tissues from mouse and human models. We further discuss the scRNA-seq and mechanistic studies that investigate the role of these fibroblast subpopulations in the context of wound healing and chronic inflammatory disease conditions.
Overview of the Structural Similarities and Differences in Oral Mucosa and Skin
Oral mucosa and skin share structural similarities and differences (Fig. 1). In humans, keratinized oral mucosa, also known as gingiva, and skin possess stratified squamous epithelial structures. The underlying connective tissue compartments are divided into papillary and reticular layers based on location and extracellular matrix features. The papillary layer is located beneath the epithelium, in which thin collagen and elastin fibers form tightly packed projections into the epithelium. The reticular layer is deeper and possesses thick and interwoven collagen and elastic fibers, creating a meshwork that provides mechanical support. In palatal gingiva, there is a submucosal adipose layer that contains minor salivary glands, similar to a human subcutaneous adipose layer surrounding eccrine and apocrine glands. In contrast, skin has other specialized apparatuses, such as hair follicle, sweat glands, and arrector pili muscles that are not present on oral mucosal layers. In the human oral cavity, buccal mucosal lining consists of nonkeratinized epithelium and is highly elastic and rich in glandular tissues, contrasting gingiva that is immovable and keratinized. The dorsum of the tongue has a thick keratinized layer and specialized papillar structures, whereas the ventrum is thin, nonkeratinized, and highly vascular. The oral mucosae in mice share these features, although all regions of the murine oral mucosa are keratinized and the thickness of each histologic layer differs between species. This review focuses on fibroblasts that are found in the connective tissue layers of oral mucosa and skin.
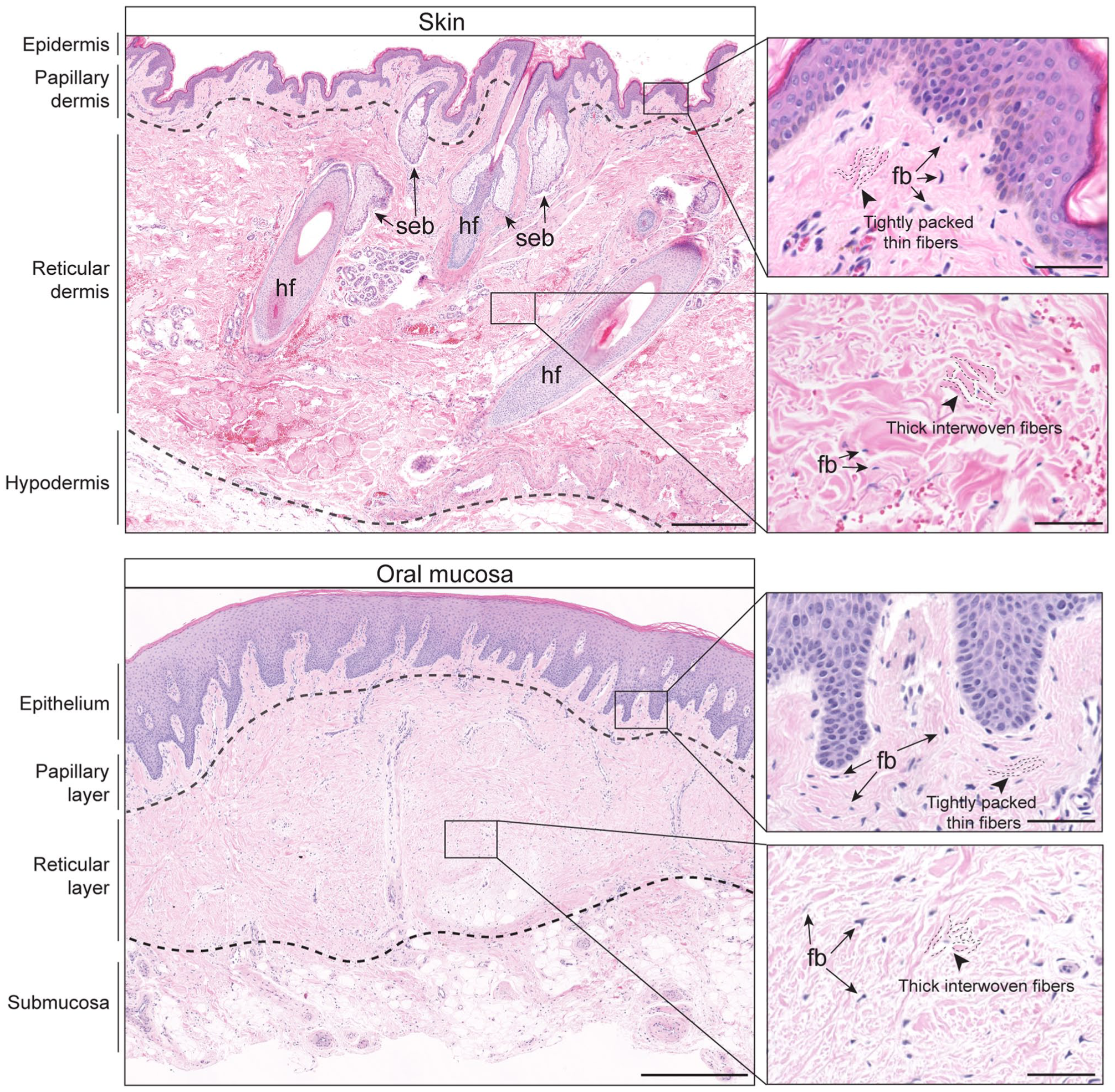
Histologic features of human skin and oral mucosa. Top: hematoxylin and eosin–stained image of human forearm skin shows different histologic compartments of epidermal, dermal, and hypodermal layers demarcated by dashed lines. Specialized appendages such as hair follicles (hf) and sebaceous gland (seb) are present in normal skin. Scale bar, 500 µm. Bottom: hematoxylin and eosin–stained image of human palatal gingiva with similarly layered architecture as compared with skin composed of keratinized stratified squamous epithelium, underlying lamina propria and submucosa. Scale bar, 1 mm. Inset images show high-magnification view of a papillary connective tissue layer that has thin and tightly packed collagen fibers and a reticular layer with thick and interwoven collagen fibers as indicated by dotted lines. Arrows point to spindle-shaped fibroblastic cells (fb). Scale bar, 50 µm.
Fibroblast Heterogeneity in the Oral Mucosa and Skin during Homeostasis
Dermal Fibroblasts
Since the 1970s, it was speculated that dermal fibroblasts from the papillary versus reticular layers were distinct by their differential proliferative capacity in vitro (Harper and Grove 1979). This was later demonstrated in murine models, in which fibroblasts within the papillary layer of neonatal skin were CD26+ (also known as DPP4) and the reticular layer and hypodermis consisted of DLK1+ or SCA1+ fibroblasts, respectively (Driskell et al. 2013). These early findings have been validated and utilized by multiple scRNA-seq studies (Thompson et al. 2022; Almet et al. 2025). Joost et al. (2020) identified 4 distinct fibroblast subpopulations from a total of 340 cells, each marked by the differential expression of Dcn, Gpx3, Sparc, or Plac8 in adult murine skin. Sparc+ and Dcn+ fibroblasts were found in the papillary and reticular layers and were predicted to play key roles in extracellular matrix and cytoskeleton organization. In contrast, Plac8+ and Gpx3+ fibroblasts were located in the adventitial and hypodermal layers, respectively, which were validated in situ. Almet et al. (2025) recently integrated fibroblast datasets from the murine scRNA-seq studies that examined unwounded and wounded conditions, which included a total of 107,778 cells. In unwounded conditions (Haensel et al. 2020; Phan et al. 2020), the fibroblast clusters with gene expression scores corresponding to upper dermis separated distinctly from those in lower dermis. Dpp4, Crabp1, Fabp1, and Prdm1 were enriched in subcluster fibroblast VIII (upper dermis), whereas Ly6a, Dlk1, and Cnn1 were found in fibroblast III (lower dermis; Almet et al. 2025), thereby displaying a distinct spatial organization within skin.
In an scRNA-seq study, Tabib et al. (2018) found 2 major dermal fibroblast populations in healthy human forearm skin, in which DPP4+/SFRP2+ fibroblasts had enriched gene expression for matrix deposition whereas FMO1+/LSP1+ fibroblasts largely expressed the CXCL12 chemokine. In a study by Boothby et al. (2021), DPP4 and SFRP2 were consistently detected in nonlesional human skin fibroblasts, although the focus was on fascial fibroblasts. Another study identified 5 fibroblast clusters in normal human abdominal skin from which DPP4, CD74, and COL6A5 emerged as key markers in distinguishing subpopulations (Philippeos et al. 2018). This study demonstrated a spatial localization of COL6A5+ fibroblasts to the papillary dermis, whereas DPP4 expression was more diffuse throughout reticular dermis, as supported by a study that examined trunk skin samples (Vorstandlechner et al. 2020). Similarly, an scRNA-seq analysis of inguinoiliac skin identified 4 fibroblast meta-clusters (Solé-Boldo et al. 2020): APCDD1+ papillary fibroblasts, CTHRC1+ reticular fibroblasts with a secretory phenotype, CCL19+APOE+ inflammatory fibroblasts, and a mesenchymal population associated with dermal papilla niche. To resolve the intrinsic heterogeneity in scRNA-seq results, Ascensión et al. (2021) integrated human skin datasets from 4 published studies and identified 3 major dermal fibroblast subpopulations. These included fibroblasts enriched in extracellular matrix–related genes such as ELN, MMP2, and SFRP2; those that express inflammation-related genes such as APOE, C7, and CYGB; and those with genes implicated in specialized dermal functions such as DKK3, TNN, and TNMD. Together, these complementary scRNA-seq findings identify gene features and a spatial niche for each fibroblast subpopulation, as summarized in Figure 2 and Table 1.
Single-Cell RNA Sequencing Studies of Fibroblasts in Homeostatic Skin and Oral Mucosa.

NA, not available.
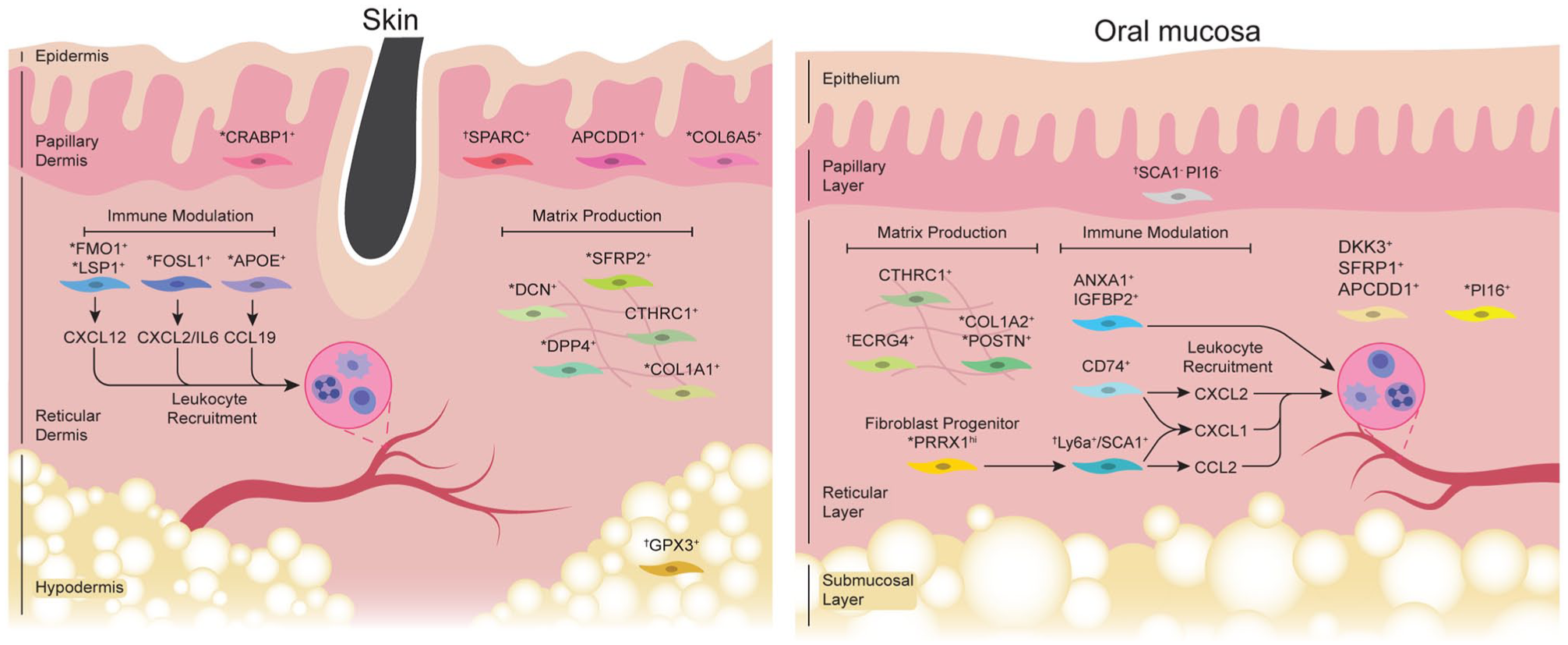
Fibroblast subpopulations in skin and oral mucosa during homeostasis. Left: fibroblast subpopulations identified from normal skin. The papillary layer is composed of APCDD1 + , CRABP1+, SPARC + , and COL6A5 + fibroblast subsets. The reticular layer contains matrix-secreting fibroblasts, including CTHRC1+, COL1A1+, DPP4+, DCN+, and SFRP2+, and immunomodulatory fibroblasts characterized by elevated expression of APOE and FMO1/LSP1, which also express CCL19 and CXCL12 for regulating leukocyte recruitment. GPX3+ fibroblasts are found in the hypodermis. Right: oral fibroblast subsets identified during homeostasis. When compared with skin, papillary oral fibroblasts remain undercharacterized. The reticular layer is composed of matrix-producing fibroblasts, including COL1A2+/POSTN+, CTHRC1+, and ECRG4+ fibroblasts. Immunomodulatory fibroblasts in the reticular layer, such as CD74+ and ANXA1/IGFBP2+ fibroblasts, express CXCL1 and CXCL2 for neutrophil recruitment. In anterior palatal gingiva, PRRX1hi fibroblast progenitors give rise to SCA1+ submucosal fibroblasts that express chemokines. Other oral fibroblast populations, such as PI16+, DKK3+, SFRP1+, and/or APCDD1+ fibroblasts, remain functionally uncharacterized. Human fibroblast subsets are illustrated unless indicated as follows: *Fibroblast subset reported from human and murine models. †Fibroblast subset reported in murine models only.
Oral Fibroblasts
Our group reported that murine palatal gingiva harbors 6 fibroblast subtypes as identified from the scRNA-seq (Ko et al. 2023). The top 4 fibroblast clusters had enriched expression of Ly6a (encoding for SCA1), Pi16, Ecrg4, and Wif1. Validation experiments revealed that Ly6a+/SCA1+ fibroblasts were found in the submucosal layer and had enriched expression of Ccl2, Ccl7, and Cxcl1, whereas Pi16+ fibroblasts resided in the reticular layer. The study further characterized a Prrx1hi/Wif1+ subset as a fibroblast progenitor population that differentiates into and replenishes SCA1 + and PI16 + fibroblasts under homeostatic or wound-healing conditions. A recent scRNA-seq study directly compared murine dorsal skin and buccal mucosa (Overmiller et al. 2024) and found a relative absence of Dpp4+ stromal cluster 2 in the buccal mucosa as compared with skin. This finding is consistent with the reports that link DPP4+ fibroblasts to skin fibrosis (Vorstandlechner et al. 2021).
There is an abundance of human scRNA-seq studies that examine healthy oral tissues. Anterior hard palate contains PRRX1hi+ fibroblasts with a pseudotime trajectory toward a fibroblast cluster that expresses CXCL1 and CCL2 (Ko et al. 2023). This PRRX1hi fibroblast subset was enriched with Wnt-modulatory genes such as WIF1, SFRP2, and WNT16. Similarly, Caetano et al. (2021) found that tooth-associated healthy gingiva has fibroblast clusters that express genes for matrix production and tissue repair (COL1A2, POSTN), the Wnt-signaling pathway (SFRP1, APCDD1, DKK3), and immune modulation (CXCL1, CXCL2, CD74, HLA-DR). In a follow-up study, the authors spatially mapped SFRP1+/POSTN+ fibroblasts to the subepithelial niche with a proposed function in epithelial maintenance, whereas the fibroblasts enriched in cytokine transcripts were found near the sulcular pocket niche (Caetano et al. 2023). Williams et al. (2021) demonstrated that human buccal mucosae exhibit different proportions of fibroblast subpopulations when compared with the marginal gingiva from clinically healthy patients. Specifically, fibroblast subtype 1.3 was more abundant in the buccal mucosa and had enriched expression of APCDD1, WNT5A, and DKK3, whereas marginal gingiva had an increased proportion of fibroblast type 1.2, which had elevated expression of CXCL13, CTHRC1, and POSTN. Buccal mucosa and gingiva had fibroblast types 1.3, 1.4, and 1.5, which were implicated in biological processes that promote leukocyte proliferation (ANXA1, IGFBP2), granulocyte chemotaxis (CXCL1, 2, 8), and complement activation (C3, CFD; Williams et al. 2021). These results agree with the study by Zhao et al. (2022) reporting that fibroblasts from buccal mucosae express CXCL1, CCL2, and CXCL12 and likely communicate with CXCR3+, ACKR1+, and CCR2+ plasmacytoid dendritic cells and T cells. Fibroblasts in the healthy tongue also express CXCL- and CCL- chemokines and likely communicate with immunocytes, although subset-specific analysis was not reported (Zhou et al. 2025). Together, these studies suggest that active leukocyte recruitment is an important aspect of normal mucosal immune surveillance during homeostasis and may be driven by distinct oral fibroblast subsets (Fig. 2).
Fibroblast Subpopulations in Wound Healing
Wound healing is a complex biological process characterized by hemostasis, inflammation, proliferation, and remodeling. Fibroblasts play a critical role in every phase, with distinct subpopulations emerging over time and across spatial niches in dermis (Fig. 3A). In a multi-omics study, Foster et al. (2021) examined murine dorsal skin wound healing across 2-, 7-, and 14-d time points and found an “activated-responder” fibroblast subset that expressed inflammation-related genes such as Ccl6, Il1b, and Il1a. The study also demonstrated an emergence of mechanofibrotic fibroblasts that expressed Dpp4, Fn1, Acta2, and En1 during the proliferative phase. Almet et al. (2025) integrated public scRNA-seq datasets from 5 independent studies that examined skin wound healing in mice. They reported that Ly6a+ fibroblasts predominate the lower dermis by day 2 and persist throughout wound closure time point. By comparison, Crabp1+ fibroblasts were located in the upper dermis and emerged by day 7 postwounding, with a presumed function in wound remodeling as Crabp1 is associated with regenerative competence (Abbasi et al. 2020; Phan et al. 2021). The study also identified proinflammatory Prg4+ fibroblasts that appear transiently in the early stages of healing and are later replaced with Col25a1+ and Pamr1+ fibroblasts that occupy the upper and lower regions of the scar, respectively.
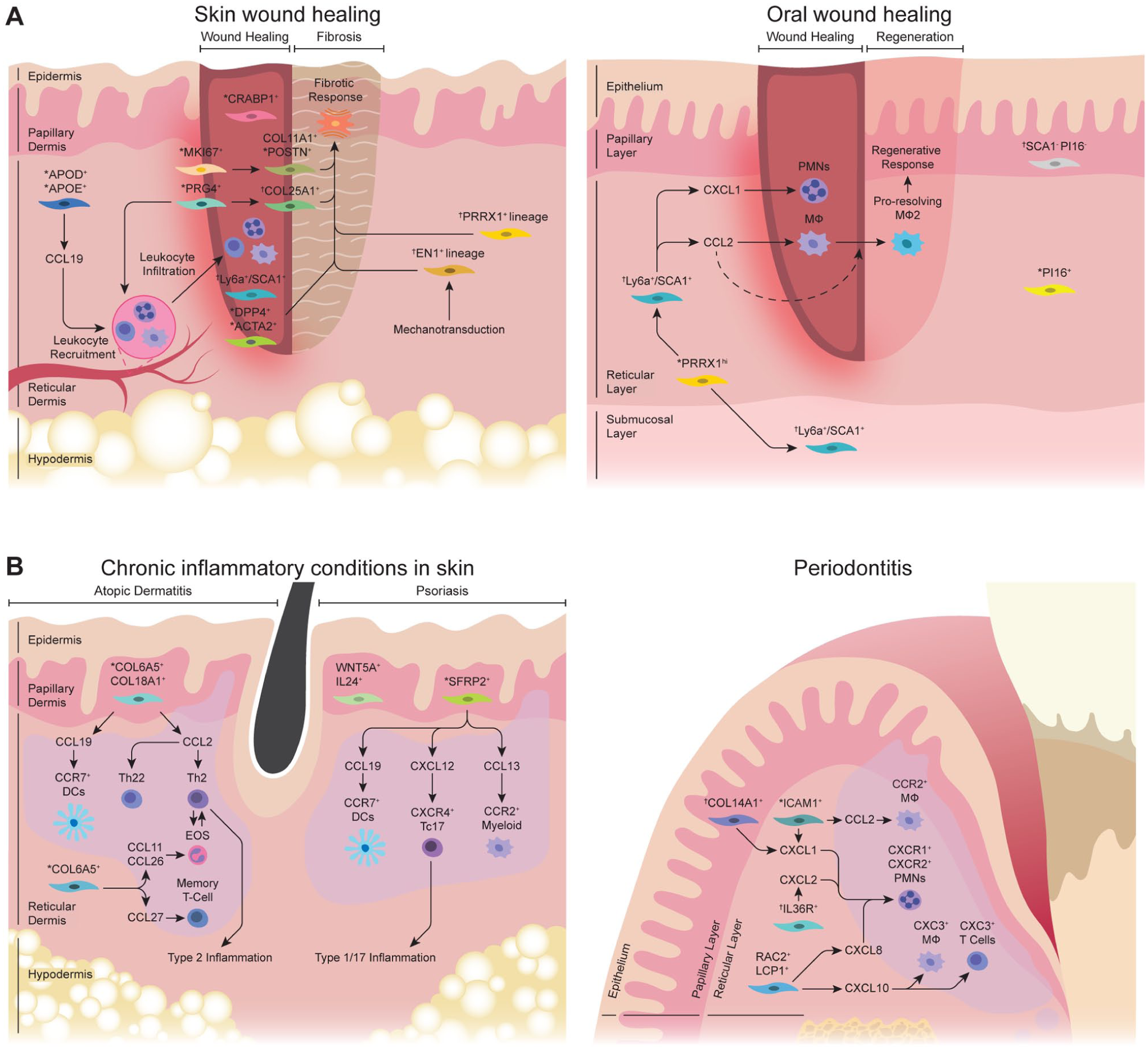
Fibroblast subpopulations under perturbed conditions in skin and oral mucosa. (
In a recent study, Liu et al. (2025) employed scRNA-seq and spatial transcriptomic approaches to examine human skin wound repair over 1-, 7, and 30-d time points. The study identified 4 major fibroblast clusters, each designated as a mesenchymal (POSTN+), papillary (ELN+LEPR+), inflammatory (C3+), and proliferating fibroblast subset (MKI67+). The proliferative fibroblasts emerged mainly on day 7 and were then replaced by COL11A1+/POSTN+ mesenchymal fibroblasts that expressed collagen-associated genes by day 30. In contrast, APOD+/APOE+ proinflammatory fibroblasts expressed inflammation-related genes such as C3 and CCL19 and declined in numbers from day 7 to 30. The increasing composition of the COL11A1+/POSTN+ fibroblast subset in a later stage of wound healing implicates a role in scar formation since elevated levels of POSTN and COL11A1 are found in human fibrotic diseases (Deng et al. 2021; Ma et al. 2024). When compared with the skin wound-healing literature, scRNA-seq studies that examine oral wounds across multiple time points are scarce, although mechanistic studies that examine select fibroblast subpopulations are available.
Functional In Vivo Oral and Skin Wound-Healing Studies
scRNA-seq studies have identified distinct fibroblast subsets during wound healing, but their biological relevance is underexplored. Our group reported that postnatal fibroblasts with active Prrx1 enhancer element (Prx1enh+) are abundant in the anterior hard palate but minimal in skin (Ko et al. 2023). These fibroblasts become highly proliferative upon oral wound induction and differentiate toward SCA1+ fibroblasts to expedite healing via CCL2 expression and macrophage recruitment (Fig. 3A). Moreover, genetic deletion of Ikbkb in Prrx1 lineage–positive oral fibroblasts suppresses their inflammatory function and delays wound healing in the palate (Chen et al. 2024), highlighting the role of immunomodulatory fibroblasts for appropriate oral wound healing. In contrast, skin wound-healing literature has focused on inhibiting profibrotic fibroblast subpopulations to reduce scar formation. For instance, genetic ablation of Prrx1 lineage–positive ventral skin fibroblasts in mice can abrogate scarring in skin wounds (Leavitt et al. 2020). In addition, DPP4+ skin fibroblasts are labeled by Engrailed-1+ lineage and possess an intrinsic ability to form scar in skin injury (Rinkevich et al. 2015). Engrailed-1 expression in skin fibroblasts is also induced by applying tension to injured wounds via mechanotransduction pathways (Mascharak et al. 2021). These mechanistic studies offer valuable insights into developing future therapeutic strategies that precisely target select fibroblast subsets to achieve a more regenerative healing outcome.
Fibroblast Subpopulations in Chronic Inflammatory Conditions
Periodontal Disease
Periodontitis is a prevalent chronic inflammatory disease characterized by persistent connective tissue inflammation and progressive destruction of tooth-supporting structures. There exists abundant scRNA-seq studies that investigate the cellular and transcriptional dynamics in human periodontitis (Caetano et al. 2021; Williams et al. 2021; Caetano et al. 2023). These studies collectively point to an inflammatory gingival fibroblast subset expressing CCL- or CXCL- cytokines as a prominent feature when comparing periodontitis with healthy groups (Fig. 3B). Williams et al. (2021) demonstrated that the fibroblast subset marked by APOD and COL11A1 is more abundant in periodontitis as compared with a healthy group and is predicted to promote the recruitment of CXCR1+/CXCR2+ neutrophils by expressing CXCL1, CXCL2, CXCL5, and CXCL8. Cell-cell ligand-receptor analysis also implicated interactions between fibroblasts that express CCL2, CXCL12, CX3CL1, and CCL19 and leukocytes that express cognate receptor genes such as CX3CR1+CCR2+ monocytes/macrophages and CCR7+CXCR4+ lymphocytes in periodontitis. Caetano et al. (2023) utilized spatial transcriptomics and identified an inflammatory fibroblast subpopulation that expresses CXCL8 and CXCL10 to be localized in proximity to leukocytic infiltration that included neutrophils, CXCR3+ macrophages, and T cells. While these human scRNA-seq datasets point to a potential role of inflammatory fibroblast subsets in periodontitis progression, they represent a snapshot of an established lesion and thus require a closer examination across varying time courses using animal models.
Murine Studies on Fibroblasts in Periodontitis
Several studies have investigated the role of fibroblasts using murine models of periodontitis. Kondo et al. (2022) demonstrated that Col14a1+ fibroblasts rapidly upregulate Ctsk to initiate connective tissue degradation as early as 1 d after ligature placement. A follow-up study demonstrated that Col14a1+ fibroblasts highly express Cxcl1 and Cxcl12 for recruiting neutrophils as well as Il6 and Cxcl10 for activation of Il17a/f-expressing innate lymphoid cells (Kondo et al. 2023). Another study reported that IL36R+-responsive gingival fibroblasts express Cxcl1 and Cxcl12 to recruit destructive neutrophils (Liu et al. 2024). The detrimental impact of neutrophils is also supported by the functional studies that inhibit their inflammatory functions (Kim et al. 2023; Qiu et al. 2024), which may be driven by gingival fibroblasts. By leveraging the existing human scRNA-seq database (Caetano et al. 2021; Williams et al. 2021), our group recently reported that ICAM-1 (intercellular adhesion molecule 1) is a reliable surface marker that identifies inflammatory fibroblasts when combined with lineage-negative and pan-fibroblast markers in human and mouse models (Kim et al. 2024). Genetic inhibition of inflammatory NF-κB activity in Prrx1 lineage–positive fibroblasts, which labeled nearly 80% of ICAM-1+ fibroblasts, paradoxically potentiated bone loss and caused excessive neutrophilic infiltration in ligature-induced periodontitis. This was partly explained by the failure to clear neutrophilic bodies by macrophages whose recruitment depended on CCL2 expression by the ICAM-1+ gingival fibroblasts.
Chronic Inflammatory Diseases of the Skin
Similar to oral mucosa, the skin barrier is susceptible to chronic inflammation (Fig. 3B). Atopic dermatitis (AD) is a prevalent chronic skin disease characterized by overt type 2 inflammation and allergic pruritus. scRNA-seq studies on human atopic skin have largely validated dysregulated epithelial barriers and inflammatory milieu but also revealed unexpected changes in dermal fibroblasts. Reynolds et al. (2021) reported that the number of fibroblast subsets that expresses CXCL12 and CCL19 is significantly higher in AD lesions when compared with healthy skin. Consistent with this, He et al. (2020) demonstrated that human atopic lesions had increased numbers of a COL6A5+COL18A1+ fibroblast subpopulation that also expressed CCL19 and CCL2 when compared with nonlesional skin. Spatial transcriptomic analysis revealed that this fibroblast subset was found near the inflammatory foci, which contained CCR7+ dendritic cells and an increased frequency of type 2 and 22 T-helper cells in AD skin (Mitamura et al. 2023).
Psoriasis is another common chronic skin disease characterized by thick scaly plaques and is biased toward type 1 and 17 inflammation (Greb et al. 2016). scRNA-seq studies on psoriasis identified a proinflammatory fibroblast population marked by upregulated expression of CCL19, TNFSF13B, and CXCL12 (Hughes et al. 2020). In another study of psoriasis, Ma et al. (2023) reported that SFRP2+ fibroblast subpopulation transitions from a profibrotic to proinflammatory state and becomes a major producer of cytokines such as CCL13, CCL19, and CXCL12. These cytokines were predicted to interact with recipient cells such as CCR2+ myeloid cells, CCR7+ dendritic cells, and CXCR4+ cytotoxic T cells that express IL17A/F. In a study that directly compared AD and psoriasis by scRNA-seq, the immune signature in the fibroblasts from AD was more prominent than that of psoriasis such that there was an increased expression of COL6A5, CCL27, and CCL26 with the latter 2 being chemoattractant for memory T cells and eosinophils (Zhang et al. 2023). Similarly, a recent study identified a WNT5A+IL24+ fibroblast population to be uniquely present in psoriatic skin but not in healthy or AD skin and that this fibroblast subset disappeared upon disease remission (Francis et al. 2024).
Functional Studies on Fibroblasts in Skin Inflammatory Diseases
In a murine AD study using epicutaneous ovalbumin sensitization, fibroblast subsets exhibited an inflammatory phenotype marked by increased expression of Ccl2, Ccl7, Ccl8, and Cxcl5 as compared with control groups (Leyva-Castillo et al. 2022). Similarly, in a murine psoriasis model that used topical imiquimod, fibroblast subpopulations displayed enriched expression of Ccl2, Cxcl12, and Cxcl14, which was minimal in control groups (Guo et al. 2024). These murine studies show that fibroblast subpopulations are an important effector cell type that amplifies inflammatory sequalae in chronic skin diseases. Recent studies that target dermal fibroblasts by genetic deletion of Ikbkb demonstrated that perturbation of fibroblasts alone was sufficient to induce AD-like symptoms in mice (Nunomura et al. 2019; Ko et al. 2022). This was due to the upregulation of eosinophil chemoattractant Ccl11 in a Col6a5+ fibroblast subset, which caused excessive eosinophilia and amplification of a type 2 immune response in the skin (Ko et al. 2022). Table 2 summarizes murine and human scRNA-seq studies that include fibroblast analyses on chronic inflammatory conditions in skin and oral mucosa.
Single-Cell RNA Sequencing Studies of Fibroblasts for Perturbed Conditions in Skin and Oral Mucosa.

FB, fibroblast; NA, not available; POD, postoperative day.
Concluding Remarks and Future Perspectives
Fibroblasts constitute heterogeneous subpopulations, each equipped with a specialized gene expression profile and spatial distribution in skin and oral mucosa. Other than extracellular matrix–associated genes, these fibroblast subsets differentially express transcripts that are implicated in inflammation and distinct molecular pathways. A major limitation in interpreting individual scRNA-seq studies is the inconsistent cell nomenclature and varying number of cell identities. This likely stems from a combination of technical variations, such as cell dissociation protocols and/or computational workflow, which can vary widely among studies. To address this, an integrative approach to categorize fibroblast subsets has been performed for skin fibroblasts (Ascensión et al. 2021; Almet et al. 2025), which consistently identified DPP4+ fibrosis-associated fibroblasts, C7+ immunomodulatory fibroblasts, and CRABP1+ wound-remodeling fibroblasts. Another limitation is that fibroblast-specific genes found in mouse models may not be found in human fibroblasts, such as Lrig1, Dlk1, and Ly6a. In a recent study, Ascensión and Izeta (2024) performed comparative analysis of mouse and human skin fibroblasts and identified multiple conserved subsets, including those expressing CXCL12 and IL6.
An integrated scRNA-seq atlas of human oral and craniofacial tissues is available (Fernandes Matuck et al. 2024). However, a detailed fibroblast-centric analysis across oral mucosal sites is lacking and remains a promising future direction. In the oral cavity, different anatomic sites are under various functional demands, and site-specific disease susceptibility is a well-known phenomenon. Similar fibroblast features implicated in chemotaxis and complement activation are found in healthy gingiva and buccal mucosa (Williams et al. 2021). This may be explained by the shared neural crest developmental origin and/or exposure to oral microbiota. Yet, it is worth noting that fibroblasts in these anatomic sites have not been compared under perturbed conditions (i.e., wounding), which may reveal divergent molecular pathways. An integrative oral fibroblast atlas may also identify stable and intrinsic markers for distinct oral fibroblast subset that must precede development of new mouse models for mechanistic studies in the future.
While informative, scRNA-seq analysis alone cannot resolve the spatial context of cells in a given niche. Spatial transcriptomics is a rapidly evolving methodology that can unravel gene expression patterns within complex tissues, and its application for examining oral structures is continually expanding (Caetano and Sharpe 2024; Haller et al. 2024). In the skin biology field, this technology has been employed for human wound healing and chronic inflammatory diseases conditions (Ganier et al. 2024; Steele et al. 2024; Liu et al. 2025). As the detection voxel size improves down to 2 µm (Visium HD) and the single-cell resolution becomes capable of detecting up to 5,000 transcripts (Xenium), we anticipate that future studies will provide a spatial context to oral fibroblast identity and function at an unprecedented resolution. A comprehensive analysis of various oral tissues in health and disease will be the crucial next step toward understanding remarkable fibroblast heterogeneity in the oral cavity, identifying functionally relevant fibroblast subsets, and developing precision therapy targeting distinct subpopulations.
Author Contributions
K. Prasongyuenyong, K.I. Ko, contributed to conception and design, data acquisition, analysis, and interpretation, drafted and critically revised manuscript; W.S. Kim, contributed to conception and design, data acquisition, analysis, and interpretation, drafted and critically revised manuscript; Z. Chen, contributed to conception and design, drafted the manuscript. All authors gave final approval and agreed to be accountable for all aspects of the work.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by funding from the National Institutes of Health (to K.I.K.; R01-DE030415 and R01-AR082951).