
Abstract
Ethnocentrism denotes a positive orientation toward those sharing the same ethnicity and a negative one toward others. Previous models demonstrated how ethnocentrism might evolve intergenerationally (vertically) when ethnicity and behavior are inherited. We model short-term intragenerational (horizontal) cultural adaptation where agents have a fixed ethnicity but have the ability to form and join fluid cultural groups and to change how they define their in-group based on both ethnic and cultural markers. We find that fluid cultural markers become the dominant way that agents identify their in-group supporting positive interaction between ethnicities. However, in some circumstances, discrimination evolves in terms of a combination of cultural and ethnic markers producing bouts of ethnocentrism. This suggests the hypothesis that in human societies, even in the absence of direct selection on ethnic marker–based discrimination, selection on the use of fluid cultural markers can lead to marked changes in ethnocentrism within a generation.
Ethnocentrism, and more generally in-group bias, is a widely observed empirical phenomena in human societies. It has many aspects and occurs in various forms (LeVine and Campbell 1972). In many contexts, people seem to divide the population into those who are considered as part of their group or their “type” (what we will call the in-group) and the rest who are seen as outsiders (the out-group). Where such distinctions are made, there is often a propensity for more positive behavior toward the in-group than toward the out-group. For example, it has been found that, under experimental conditions, even arbitrary group assignments produce in-group positive and out-group negative behavior (Tajfel et al. 1971; Brewer 1979).
In order to explore possible mechanisms that might produce such phenomena, minimal computer simulation models have been presented in which evolutionary processes lead to ethnocentrism emerging over time (Hammond and Axelrod 2006; Jansson 2013). 1
In these models, agents are located on a spatial grid and evolve intergenerationally (i.e., vertically) with new agents being born, inheriting parent traits, and old agents dying. Both interaction and reproduction are localized in neighborhoods on the grid, and agents cannot change their behavior or location during their lifetimes.
Results from these models show that eventually agents come to favor their in-group, which is defined by an observable ethnic marker (or color). The ethnic marker evolves in the same way and at the same rate as behavioral traits through mutation and selection over generations. Hence, these models focus on long-term intergeneration vertical evolutionary dynamics—no intragenerational learning can occur.
We present a model with a different emphasis by considering short-run intragenerational horizontal cultural dynamics related to a fixed ethnic marker and evolving fluid group identities that are not related to ethnicity.
Our model produces results that contrast with the previous models, showing how purely horizontal adaptation can lead to the emergence of ethnocentric behavior. This results from the formation of subcultures that discriminate in terms of ethnicity in combination with fluid group identities.
This article is structured as follows. Firstly, we describe an existing canonical simulation model of ethnocentrism and other relevant work. Then, we present the motivation and assumptions of our model. We then describe our model in detail and present results obtained from a number of simulation experiments. We then interpret and discuss the results. We conclude with the implications of the model results in the context of wider research questions.
Previous Models
Hammond and Axelrod Model of Ethnocentrism
Hammond and Axelrod (2006) introduced a seminal artificial society, agent-based, simulation model of ethnocentrism. In their model, agents are represented with three traits: (1) a color; (2) an in-group strategy, and (3) an out-group strategy. There are four different colors representing different ethnicities. Each strategy takes a binary value of either to cooperate or defect in a Prisoner’s Dilemma (PD) game. 2 Hence, there are 4 × 2 × 2 = 16 different possible agent types.
Agents are situated on a 50 × 50 grid. 3 Each grid location may contain a maximum of one agent or be empty. Initially, the grid is empty, but in each time period, a new “immigrant” agent is placed in a random empty location on the grid. The traits of these new agents are generated randomly from the sixteen different types possible.
In each time period, agents also play the pairwise single-round PD game with their four neighbors (north, south, east, and west) on the grid. 4 When a game is played, each agent compares its color to its partner. If they match, it plays the strategy specified by its in-group trait. If they do not match, then the out-group strategy is used. Hence, agents that share a color consider themselves as an in-group. When an agent cooperates, it incurs a cost, but it receives a benefit when its partner cooperates. The benefit is three times greater than the cost. Over the time period, each agent accumulates the costs and benefits of their game interactions producing a final payoff value.
At the end of a time period, agents may reproduce and/or die. Reproduction involves making a copy of the reproducing agent into an empty neighboring grid location if one exists. Each trait of a reproduced agent is mutated with low probability. 5 The probability that an agent will reproduce is a function of its payoff. High payoffs mean an agent is more likely to reproduce. 6 Agents die with a fixed probability of 10 percent, this creates space on the grid allowing new agents to enter.
An agent is defined as ethnocentric if it has a cooperative in-group strategy and an out-group defect strategy—that is, it donates only to neighboring members of its in-group (those sharing the same color). Results from simulation runs show high levels of ethnocentrism (of the order of 80 percent) emerge. This result holds over a range of parameter settings such as grid size, number of colors, and mutation rate. However, it was subsequently found that the spatial localization of both reproduction and game interaction are necessary conditions for ethnocentrism to emerge. 7
Tag Models
A similar but more general treatment of in-group cooperation can be found in “tag” models. Here, individual agents have a trait (or tag) that is visible to other agents and not hardwired to any particular kind of behavior or behavioral tendency (Geisel 1961; Holland 1993).
The in-group in these models is defined as those that have close or matching tags (Riolo 1997; Riolo, Cohen, and Axelrod 2001; Hales 2000, 2010; Shutters and Hales 2013; Jansen and van Baalen 2006). Agents store a single tag 8 that takes a value from a very large set (as opposed to a small set of colors) and a single strategy (either cooperate or defect). There is no separate in-group/out-group strategy, but agent interactions are often biased (in terms of more likely to occur or more likely to be positive) toward the in-group. The Hammond and Axelrod model is a specialized tag model with a small set of possible tags (colors), explicit in-group/out-group strategies, and strict spatial interactions.
Riolo, Cohen, and Axelrod (2001) introduced a tag model that allows agents to redefine their in-group over contiguous ranges of tag values (represented as real numbers) where agents do not store a strategy but rather are hardwired to cooperate with their in-group and defect on their out-group. Hence, they have no possibility to evolve different strategies for their in-group and are effectively forced to cooperate with others sharing exactly the same tag (Roberts and Sherratt 2002). Variants of this latter model have incorporated an evolving strategy such that agents can defect on their in-group (Edmonds 2006; Shutters and Hales 2013).
Fu et al. (2012) apply evolutionary set theory to examine the conditions under which in-group and out-group cooperation can emerge where differential strategies can be applied to them. They find that a large number of groups are required with a high level of group migration (relative to strategy mutation) to support in-group cooperation.
Axelrod, in his book on the evolution of cooperation, briefly discussed the effect that fixed labels (similar to the ethnic markers we use in our model) could have on interactions between agents playing the Iterated PD (Axelrod 1980). He described how these could lead to poor cooperation between those not sharing the same marker. However, our scheme is not directly comparable to the work presented there. 9
Motivation and Assumptions
Some forms of ethnocentric behavior may be long standing, passed down from generation to generation, but this does not explain its sudden emergence within a generation. We are interested in short-term, within generation, emergence of ethnocentric behavior such as sudden upsurges of ethnocentrism within communities.
In an intragenerational context, ethnic identity is relatively fixed, but other cultural traits may be fluid. It is rare for ethnicity to change within a generation but common for other more fluid traits to do so (e.g., style of dress or opinions).
In the models previously discussed, each agent holds a single marker trait (either a color or a tag) that is not fixed but evolves along with other agent traits (such as strategies). Consequently, these models cannot be interpreted as modeling the intragenerational emergence of ethnocentrism because in this context the ethnic marker would be fixed.
Also, in the previous models, agents are hardwired to view others with matching markers as members of their in-group. We are interested in the situation where in-group identification is itself a fluid and evolving trait such that agents have the ability to change their in-group definitions (possibly ignoring ethnicity completely).
We ask the following question: can discrimination based on fixed markers (which we interpret as ethnic group membership) evolve even when the markers themselves cannot evolve?
In order to address this question, we have created a model in which we endow agents with (1) an intragenerationally fixed ethnic marker (similar to color in the Hammond and Axelrod model) that does not change and three intragenerationally fluid traits: (2) a cultural identity (a cultural tag—similar to a tag in the previous tag models); (3) a definition of how an agent identifies its in-group based on ethnic marker and cultural tag; (4) a set of strategies for interacting with in-group and out-group individuals when they encounter them. Each of these traits is described in more detail in the Model section.
By incorporating a fluid in-group definition, we capture, in a minimal way, the notion that agents can form dynamic and composite in-group identities based on both fixed ethnic markers (which do not change) and fluid cultural tags (which do change). Hence, the ethnic marker is fixed, but the way agents define their in-group is not.
In our model, ethnic markers are stable classifications that others can observe. They might represent genetically determined and/or vertically transmitted cultural characteristics (ethnicity), but they could just as well represent any observable characteristic that is relatively stable during an agent lifetime (e.g., accent, gender, social class, nationality). The requirement is that they are stable relative to short-term horizontal intragenerational cultural change. 10 Hence, the functional aspect of ethnicity we model could relate to any piece of information that is visible to others but cannot be easily imitated or changed and therefore is intragenerationally stable.
In our model, cultural tags represent publicly identifiable, imitable, and mutable markers that can be evolved horizontally, intragenerationally, such as clothing style, publicly expressed opinions, or other fluid group identifiers.
The evolutionary process in our model is based on imitation and innovation occurring within the lifetime of the agents. Agents do not reproduce or die but inhabit a fixed-size population. If an agent detects that another is doing better than they are (in terms of payoffs from a simple donation game), they imitate the other agent. Imitation involves copying all the fluid traits of the other: its cultural tag, in-group definition, and its in-group/out-group strategies but not the ethnic marker (which cannot be copied or changed).
While imitation is a complex process in human societies, we use this minimal method. Our assumption is that agents are not able to identify and copy the individual traits that lead to success but rather they emulate wholesale others who outperform themselves. This could result from cognitive limitations in complex social environments. Alternatively, this assumption could represent social conformism. In human societies, such conformism can result from a desire to “fit in,” fear of sanctions, norms, or other social monitoring processes. This is sometimes termed “social docility” (Simon 1990). This is a critical assumption in our model (and the others discussed) because if agents could intelligently and selectively copy specific traits purely for their own benefit, then it would be unlikely that any cooperation would emerge. This approach implements a form of replication in an evolutionary process.
Innovation involves agents spontaneously changing their fluid traits randomly. The assumption is that occasionally agents may change their behaviors for numerous reasons. These may include an error in imitation or some other contingent event resulting in a change of attitude. We do not model these processes directly but rather introduce random noise with low probability. This implements a form of “mutation” in an evolutionary process.
We do not constrain interactions between agents by spatial proximity; hence, there is no space in our model. We do not model space because it has been shown that spatial interaction in itself favors the emergence of ethnocentrism, but we are interested in interactions based purely on tag and marker processes (Jansson 2013). 11
However, game interactions are constrained by in-group membership, with agents preferring to interact with their in-group. This is inspired by the notion that a densely populated locality or Internet community offers a wide choice of interaction opportunities, but individuals prefer to interact with those within their in-group when trust is an issue. 12 We do not model the specific mechanisms by which agents perform this searching, but such mechanisms could include social networks, institutional and social gathering places, clubs, meeting places and online forums, and so on.
The assumptions that game interactions are constrained to the in-group but imitation is population wide are known to produce in-group cooperation from the previous models discussed. By adopting them in our model, we expect to see high levels of in-group cooperation emerge.
Model Description
Here, we describe our model in detail—we term it the “ethno-cultural tag” model.
Agents store traits that determine their behavior. They interact by playing donation games and through selective imitation, which involves agents copying the traits of others. Agents spontaneously innovate by occasionally changing their traits randomly. This supports a minimal form of horizontal (within generation) cultural evolution where some traits spread and others disappear or spontaneously appear. Hence, the behaviors of agents change over time, but agents do not die or reproduce.
Firstly, we describe the traits stored by each agent, then the events and parameters that regulate them. Finally, we describe sequencing of the events during a simulation run of the model.
Agent Traits
The model consists of fixed-size population of N agents. Each agent stores five traits: (1) an ethnic marker, (2) a cultural tag, (3) an in-group selector (which specifies the in-group definition used by the agent), (4) an in-group strategy, and (5) an out-group strategy. Table 1 summarizes the agent traits. In the following section, we describe each of these traits in more detail.
Summary of Agent Traits.

Note: Each agent stores these five traits. NE and N are model parameters. The meaning of these is discussed in detail in the text. NE = number of ethnicities; N = number of agents in population.
The ethnic marker and cultural tag
The ethnic marker is a fixed observable marker preassigned to each agent. Agents assigned the same ethnic marker are said to share the same ethnic group. The fluid cultural tag is an observable marker indicating cultural group membership. Those who share the same cultural tag are said to share the same cultural group. Agents may change their fluid cultural tag based on the intragenerational horizontal evolutionary processes (see later), but their ethnic marker is preassigned and immutable. We allow for N possible unique cultural tags. This means that it is possible, maximally, for all agents to hold a distinct cultural tag or, conversely, for all agents to share the same cultural tag.
The in-group selector
Agents can only distinguish between other agents by observing their ethnic marker and cultural tag. That agents do not remember others as individuals is a simplification. In a sufficiently large population, many interactions will not be with individuals one knows, yet one has to decide how to behave toward them. The in-group selector determines how an agent decides if others are part of their in-group based on the two (ethnic and cultural) observable traits.
The in-group selector can take one of four possible types; thus, an agent defines its in-group as one of: those with the same ethnic marker as itself (ethnic), those with the same cultural tag as itself (cultural), those with the same ethnic maker and the same cultural tag (both), or any other agent without restriction (none).
13
Agent strategies
Agents store two strategies that are used during game interactions (see below). They store one for the in-group and one for the out-group. These are independent of each other and to their in-group selector. They can be either donators (cooperators) or “shirkers” (defectors) with respect to the in-group and separately with respect to the out-group (as defined by the in-group selector).
This means an agent can hold one of four possible strategy combinations: shirk against both the in-group and out-group (ss); donate to both the in-group and out-group (dd); donate to in-group, shirk on out-group (ds); or shirk on in-group, donate to out-group (sd).
Initialization
At the start of each simulation run, all the fluid agent traits (the in-group selector, the cultural tag, and the in-group and out-group strategies) are initialized to random values from their range (shown in Table 1). 14 Fixed ethnic markers are initialized such that the population of N agents is equally divided between the number of ethnicities (NE). Hence, if NE = 2, this means that 50 percent of agents share one ethnic marker and 50 percent share the other.
Model Events and Parameters
Interaction—A donation game
Agents interact, in pairs, by playing a “donation game” in which one initiating agent must decide if to provide help (a donation) to the other partner agent. If the initiator decides to donate, it incurs a cost (C) to itself while the receiving partner agent gains a benefit (B). We consider the situation where B > C, hence the benefit-to-cost ratio B/C > 1. If B/C = 2, then this means the benefit is twice the cost.
Thus, a receiving agent benefits more from the action of the donor than the cost incurred by the donor. For example, this could occur if the donor had a surplus of some good, which is of little value to themselves but of great value to a receiving agent. Alternatively, an agent could up-vote or otherwise positively comment on content provided by another in an online forum.
During a donation game, the initiating agent uses its in-group selector to decide if its partner is an in-group or an out-group member. It then enacts the behavior (or strategy) indicated by its in-group strategy or out-group strategy, respectively. Each strategy takes one of two types, either “donate” or “shirk” (meaning don’t make a donation). Only the initiating agent selects a strategy and plays the game. The partner agent is passive either receiving a donation or not. 15
Over time, different pairs of agents play the donation game and accumulate a total payoff based on benefits received minus costs incurred.
Imitation
Periodically, an agent compares its payoff to another partner agent in the population. If the other agent has a higher payoff, then all of their traits, excluding the ethnic marker, are copied by the initiating agent (overwriting their existing traits). This scheme implements a heuristic in which agents copy those who are doing better (in terms of payoff), implementing a form of horizontal cultural reproduction.
Innovation
Periodically, with low probability, an agent changes each of its fluid traits: cultural tag, selector, in-group strategy, and out-group strategy (independently) to a random value from their range. This implements a form of cultural mutation. The mutation rate probability M is used for each fluid trait other than the cultural tag to which a larger rate, 10M, is applied. We inherit this assumption from previous models. It has been shown to be sufficient to produce cooperative groups without the need for strict spatial interaction and is explicit or implicit in most previous tag models. 16 This implies that agents are more likely to change their observable cultural tag than their basic behaviors toward others. Another way to view the larger mutation rate applied to the cultural tag is that it is functionally equivalent to an agent holding several cultural tags with each independently mutating with probability M.
Selecting partners
For both game interaction and imitation events, agents need a method to select a partner agent from the population.
For game interaction, this involves an initiating agent randomly selecting a partner from its in-group (as defined by its in-group selector). If there are no other agents within its in-group, then a partner is selected randomly from the entire population. For imitation events, a partner is randomly selected from the entire population—ignoring the in-group selector. Hence, agents imitate over the entire population, but game interact within their in-group. Note, results from experiments that relaxed this strict assumption are discussed in the Further Experiments section.
Table 2 summarizes the parameters of the model with the values used for the experiments that follow. Further experiments with different values are discussed in the Further Experiments section and in the Online Appendix in Supplemental Material.
Summary of Exogenous Model Parameters.

Note: The specific values used for B and C were 0.2 and 0.1, respectively.
Sequencing of Events
The simulation sequences events into cycles, which represent some notional unit of time. Each cycle involves three phases: (1) game interaction, (2) imitation, and (3) innovation (in that order).
In phase 1 (game interaction), each agent in the population is selected in a random order and initiates a game interaction. This involves it selecting a partner, based on the mechanism described above, and then playing a donation game with the selected partner (also described above). After a game interaction, the initiating agent and the partner’s payoffs are updated accordingly.
In phase 2 (imitation), each agent in the population is selected in a random order and initiates an imitation interaction. The initiating agent selects a partner randomly from the entire population. If the partner has a higher payoff, then all its traits are copied (other than the ethnic marker). Only the initiating agent decides whether to copy or not based on payoffs. 17 The partner agent is passive and does not perform any imitation action even if it has a lower payoff than the initiating agent. 18
In phase 3 (innovation), each agent randomly changes its traits (other than the ethnic marker) based on the mutation (M) parameter: independently, the in-group and out-group strategies are flipped (from donate to shirk or vice versa) with probability M. Also with probability M, the in-group selector is replaced by a random variant. 19 With probability 10M, the cultural tag is replaced by a randomly selected tag.
A simulation run involves repeatedly executing phases 1, 2, and 3 for some number of cycles after which the run terminates. Further details on the implementation of the simulation model can be found in the Online Appendix in Supplemental Material.
Simulation Results
Method
A number of simulation runs (experiments) were performed with different parameter settings and the following measures were collected: the donation rate (dr), which indicates the proportion of all game interactions that led to a donation occurring; the interethnic donation rate (ie), which indicates the proportion of the donations made that involved agents with different ethnic markers; the intercultural donation rate (ic), which indicates the proportion of the donations made that involved agents with different cultural tags; the proportion of each of the four in-group/out-group strategy types in the population: ss, sd, ds, and dd; and the proportions of the four different in-group selector types in the population: none (sn), cultural (sc), ethnic (se), and both (sb). These measures are summarized in Table 3.
Summary of Measures Collected from Simulation Runs.

Each simulation run was executed for 3,000 cycles. For each run, statistics were based on averages over the last 1,000 cycles. Each run was replicated twenty times with different initial pseudorandom number seeds. Averages and standard deviations of the “1,000-cycle averages” were then calculated over these twenty runs. Hence, the standard deviations are over these twenty data points not the within run variation.
Results
Populations with a single ethnic marker
Firstly, we consider the results obtained with only one ethnicity in the population (NE = 1). In this case, agents cannot, in practice, discriminate based on ethnic markers since all agents share the same marker—the population is ethnically homogeneous. This case serves as a baseline indicating the results obtained when no distinctive ethnic markers exist. Hence, these results are presented in order to better understand the results obtained when more than one ethnicity is introduced in the subsequent experiments.
Results for NE = 1 are shown in the first row of Table 4. Notice that the donation rate is high (dr > .9). The interethnic donation rate (ie) is zero because all donations must, by definition, be between agents of the same ethnicity. The intercultural donation rate (ic), not shown in Table 4, was negligible (< .01 with low variation) indicating that almost all donations were between agents sharing the same cultural tag.
Results Obtained, in Terms of Measures Listed in Table 3, for One and Two Ethnicities (NE) Averaged over Twenty Independent Runs.
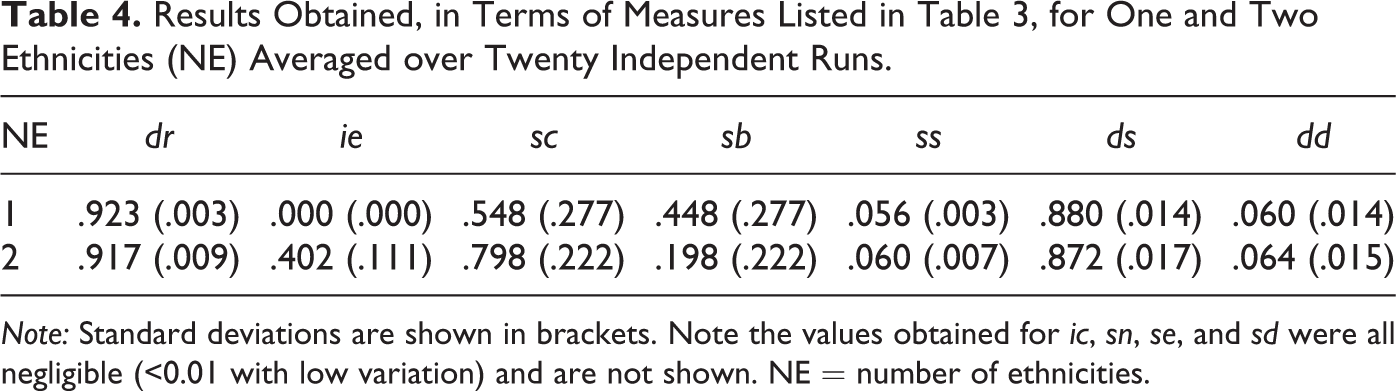
Note: Standard deviations are shown in brackets. Note the values obtained for ic, sn, se, and sd were all negligible (<0.01 with low variation) and are not shown. NE = number of ethnicities.
The dominant in-group selectors are cultural (sc) and both (sb), which are more or less equally split and have high (and equal) standard deviations. The none (sn) and ethnic (se) selectors, not shown in Table 4, were negligible.
The dominant strategy is in-group donation and out-group shirking (ds ≅ .9) while the unconditional donation and shirking (dd and ss) strategies have roughly equal low values around 6 percent. The in-group shirking and out-group donation strategy (sd) was negligible and is not shown.
Taken together, these results indicate the predominance of agents that donate to their in-group defined as those with a matching cultural tag.
The split between cultural (sc) and both (sb) selectors is not surprising because, in this case (with only a single ethnicity), the sc and sb selectors produce exactly the same functional behavior. Both define the in-group as those with a matching cultural tag because ethnicities always match. Hence, there is no selective pressure between them resulting in a passive “drift” between the two. The high and equal standard deviations of sc and sb indicate that each individual simulation run produces a variation of results with sc and sb being inversely proportional to each other.
The high donation rate (dr) and in-group donation plus out-group shirking strategies (ds) are consistent with results obtained in previous evolutionary tag models (Riolo 1997; Riolo, Cohen, and Axelrod 2001; Hales 2000). In those models, high levels of cooperation (donation) were obtained when game interaction was based on tag similarity. This donation sustaining process results from a dynamic process of tag group formation (based on shared tag values) followed by their dissolution. This has been equated with a form of “group selection.” 20 An explanation of this process with a comparison of a number of similar models is given in Hales (2010); hence, we will not discuss this in detail here because this is not a novel result from our model. However, in order to indicate the kind of tag group dynamics involved, we give a brief outline before we proceed to discussion of results for multiple ethnicities.
Figure 1 shows a visualization of the emergence and dissolution of cultural tag groups taken from a time series of part of an individual simulation run. Notice that cultural tag groups (agents sharing a tag) form and dissolve over time. Groups undergo a “life cycle” of stages that might be called: seeding, growth, decay, and death. Typically, a new small cooperative group forms with a cultural tag that is otherwise unused (due to innovation). This usually comprises two agents with strategies and selectors such that they donate to each other.
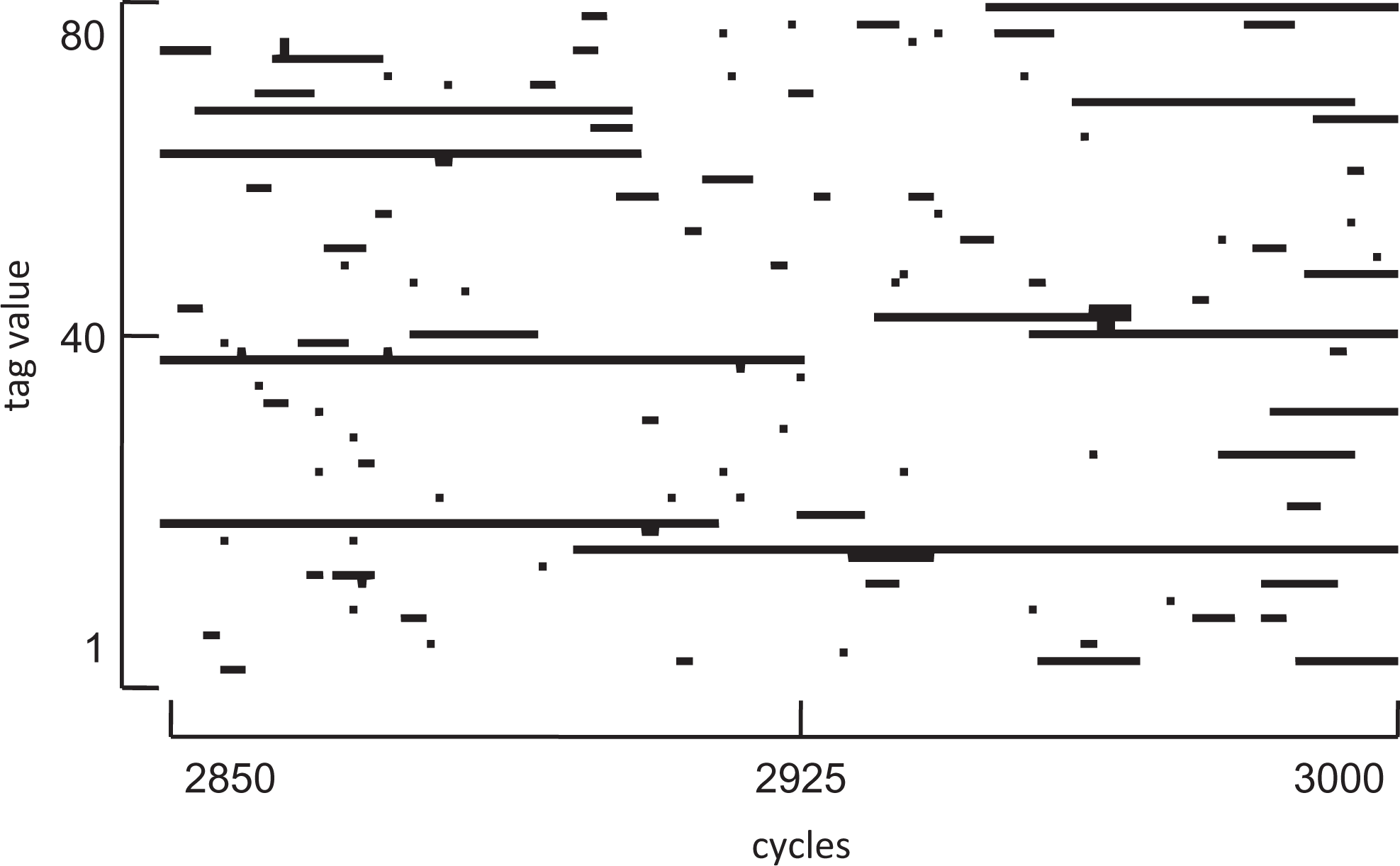
Time series of cultural tag groups for a portion of a single run. Simulation time is shown on the x-axis and cultural tags on the y-axis. A dot is plotted if at least one agent holds the given tag at the given time. Hence, a long line indicates a cultural group lasting some continuous time period, whereas a single dot or short line represents a short-lived transient group. Only the first 80 tags are shown over the last 150 cycles; however, the dynamics are typical over the entire space of the run. This run is for the number of ethnicities = 2.
The agents in the group do well in terms of payoffs because each donates to the other. Other agents imitate them creating more donating group members. Consequently, the group grows. When the group is large, shirkers eventually appear by innovation of strategy—these decrease the payoff to other agents in the group since the shirkers exploit them. However, the shirkers temporarily do better than the donators in that group, since they gain donations but do not give donations. This means others imitate the shirkers (becoming shirkers themselves), leading to a decline in the advantage of being in that group. Eventually by imitation, agents move to other groups and the cultural tag disappears from the population.
So long as new groups are created more quickly than they are dissolved, by shirking members, then high levels of donation can be maintained through this ongoing dynamic process. In our model, this condition is met by the assumption that the cultural tag mutation rate is higher (10M) than the strategy and selector mutation rate (M).
Populations with two ethnic markers
As can be seen in Table 4, the experimental results for populations with two ethnicities (NE = 2) show a similar donation rate (dr) and distribution of strategies as the single ethnicity case (NE = 1). Hence, introducing ethnic diversity into the population does not effect these. However, the distribution of in-group selectors is different. The cultural selector was more dominant at approximately 80 percent (sc ≅ .8) and the both selector was consequently lower at approximately 20 percent (sb ≅ .2). Again notice (as before in the single ethnicity case) the high and equal standard deviations for the cultural (sc) and both (sb) selectors indicating significant differences between individual simulation runs where sc and sb are inversely related.
It is important to note that with two ethnicities, the cultural and both selectors have different functionalities—which was not the case in the single ethnicity case. Specifically, the both selector defines the in-group based on ethnic marker and cultural tag. Hence, an agent using a both selector excludes those who do not share its ethnic marker (in this case, half the population) from its in-group. This precludes interethnic donation when combined with the ds strategy (i.e., donate to in-group, shirk on out-group).
Table 4 shows the average interethnic donation rate (ie) as a proportion of all donations made. Since the population is equally split between two ethnicities, we would expect ie ≅ .5 if no discrimination based on ethnic marker was occurring. Given ie ≅ .4, this means that the number of interethnic donations are approximately 20 percent less than would be expected if no discrimination on ethnic marker was occurring. Hence, we see an inversely proportional relationship between ie and sb where sb equates to the proportionate reduction in ie over the expected value. This is because almost all agents holding a both selector also hold a ds strategy and hence will not donate to others with a different ethnicity
Figure 2 shows example time series for simulation runs with two ethnicities. Notice that the cultural and both selectors do not persist in some fixed proportion over time but rather compete and vary over time. When the both selector is high, then interethnic donation (ie) and cultural sector (sc) are low. In general, ie is inversely proportion to sb and proportional to sc.
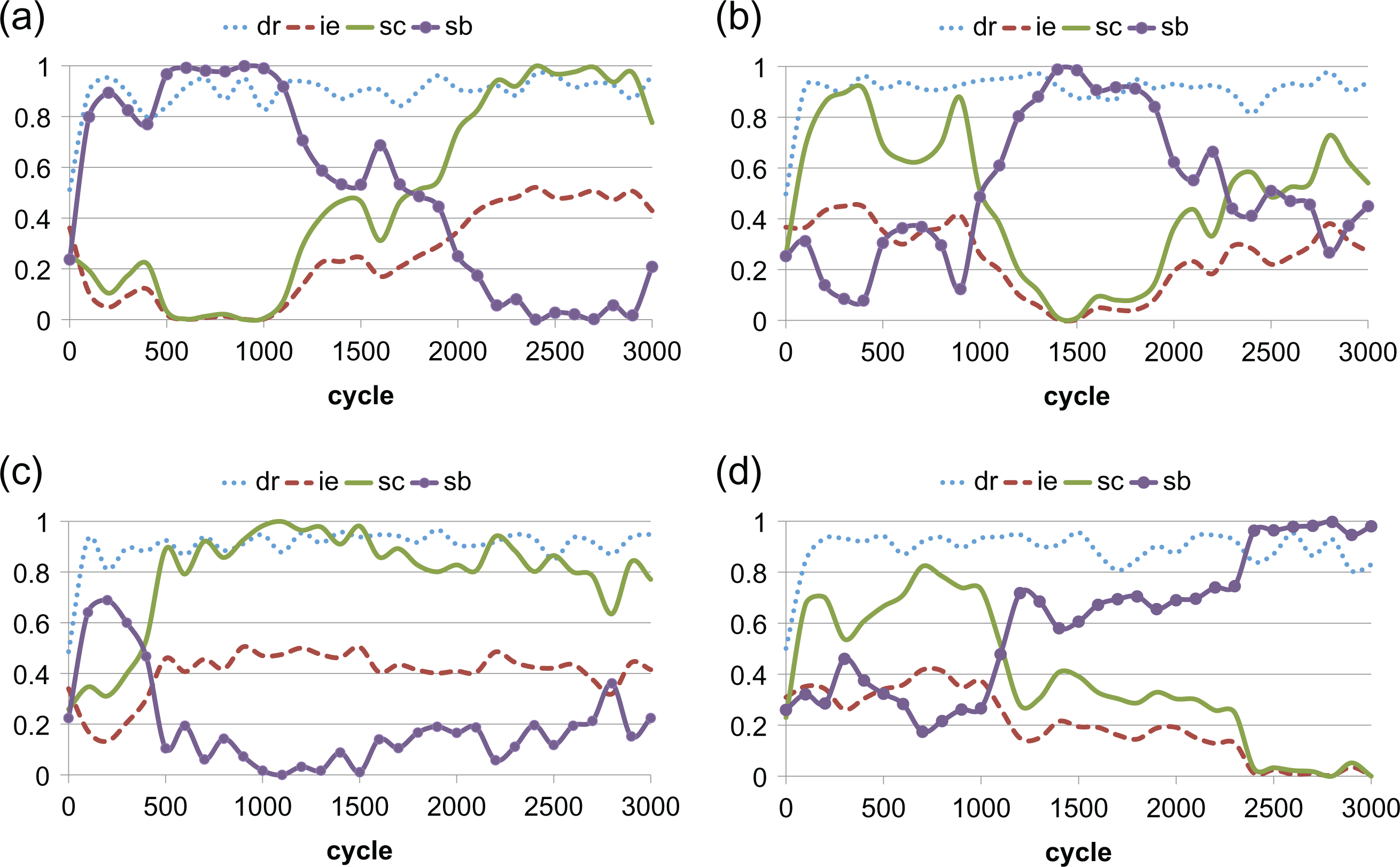
Time series of four independent simulation runs with two ethnicities (number of ethnicities = 2). Shown measures are donation rate (dr), interethnic donation rate (ie), cultural (sc), and both (sb) selectors. (a) Shows domination of the population by the cultural selector after initial domination by the both selector. (b) Shows an oscillation between the two. Notice that when sb is high, ie is low. (c) Shows a more typical run in which sc dominates. (d) Shows a run where sb comes to dominate.
The macro dynamics in Figure 2 are the aggregate of the group micro dynamics shown in Figure 1. To understand the relation between them, we examine the micro dynamics of the cultural tag groups. We found that almost all tag groups of size >5 comprised agents holding the ds strategy combined with either the both or the cultural selector for the majority of their existence. It is very rare that these selectors are mixed within a group.
This can be understood by examining the group formation process. New groups are formed through mutation on the cultural tag moving an agent to a currently empty tag value. This produces a group of size one (a one-seed). A one-seed with either a cultural or both selector has no in-group members. This results in a game interaction with a random partner from the entire population and hence the enactment of the out-group strategy. Hence, an out-group shirk strategy combined with either of these selectors maximizes agent payoff because it will not make a donation. Minimally, it will receive a payoff of zero but could receive more if it happens to receive a donation from another agent.
A one-seed grows if it can recruit other agents through imitation. This will happen if another agent chooses the one-seed for imitation and has a lower payoff. In which case, the agent will join the group by copying the one-seed tag, strategy, and selector. At this stage, a group will only produce positive payoffs for its members if its members donate to the in-group. Given each group is in competition with many other such groups, those with in-group donate are more likely to be imitated. This two-stage selection process inhibits a one-seed from growing unless it is either a cultural or both selector combined with the ds strategy.
But groups do not grow indefinitely, rather they have finite life span during which they grow, reach a maximum size, decline, and finally die leaving the tag empty for a new seed to potentially enter and start the process again. This occurs because strategy mutation will eventually produce a mutant that holds a shirk in-group strategy thus exploiting the in-group by not making donations but still receiving them. The mutant thus receiving a high payoff will recruit others through imitation that will also exploit the in-group. Exploited agents will receive the lowest payoff (of −C) and hence are likely to leave the group through imitation of higher scoring agents in other groups. Eventually, the group will contain only in-group shirkers and produce zero payoffs for all. This explains the ss value shown in Table 4.
Another form of mutation that sometimes invades part of a group is the out-group donation strategy (dd). This happens because it is functionally equivalent to the ds strategy for groups that have gone beyond the early stage since they do not engage in out-group interactions. This explains the dd value shown in Table 4. Figure 3 shows two small example groups taken from a run to illustrate this life cycle.
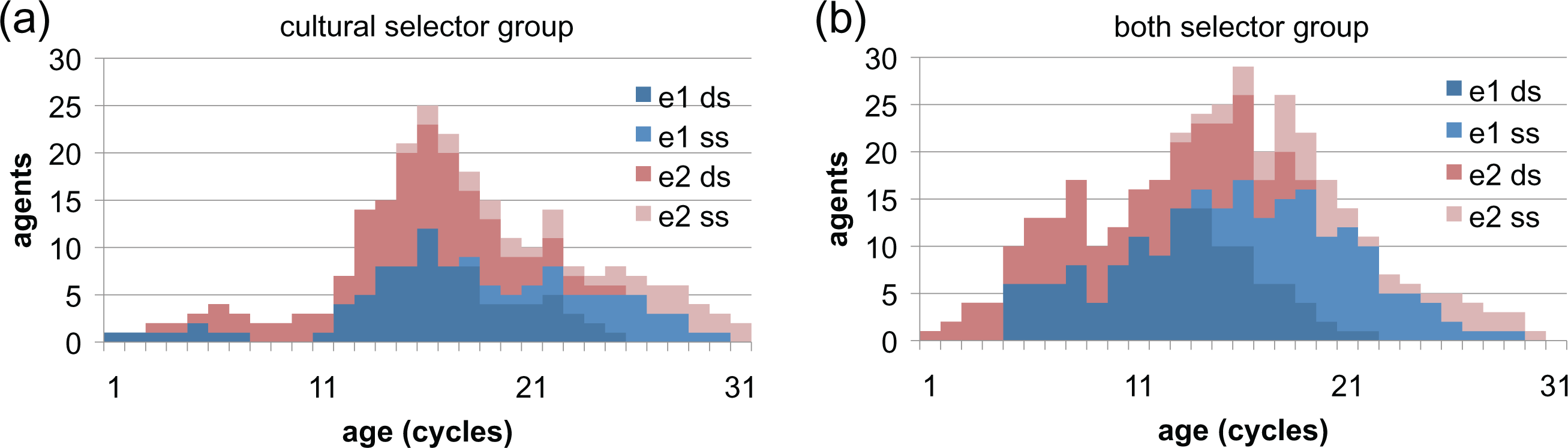
Time series (stacked columns) of two example cultural tag groups from a simulation run with two ethnicities. (a) Shows a group composed entirely of cultural selectors and (b) a group of both selectors. The two darker shades indicate ethnicity one (e1) and the two lighter shades indicate ethnicity two (e2). The lighter shades within each ethnicity indicate the in-group shirk strategy (ss), the darker the in-group donate (ds) strategy. Notice the groups start with a single agent “one-seed,” grow, become invaded by in-group shirking mutations, and then decline and die.
We have seen that almost all groups comprise cultural or both selectors combined with the ds strategy initially. But what causes the both selector to be less successful than the cultural selector when there is more than one ethnicity? One way to understand this is to consider the early-stage evolution of groups.
Some early-stage game interaction possibilities are shown in Figure 4. Notice that in the case of mixed ethnicity groups, donations are constrained or reduced. Comparing Figure 4b and f notice that in the latter case, no donations occur because the agents are of different ethnicity. Hence, both selector groups are less likely to grow. Yet, in many cases, both selector groups do succeed.
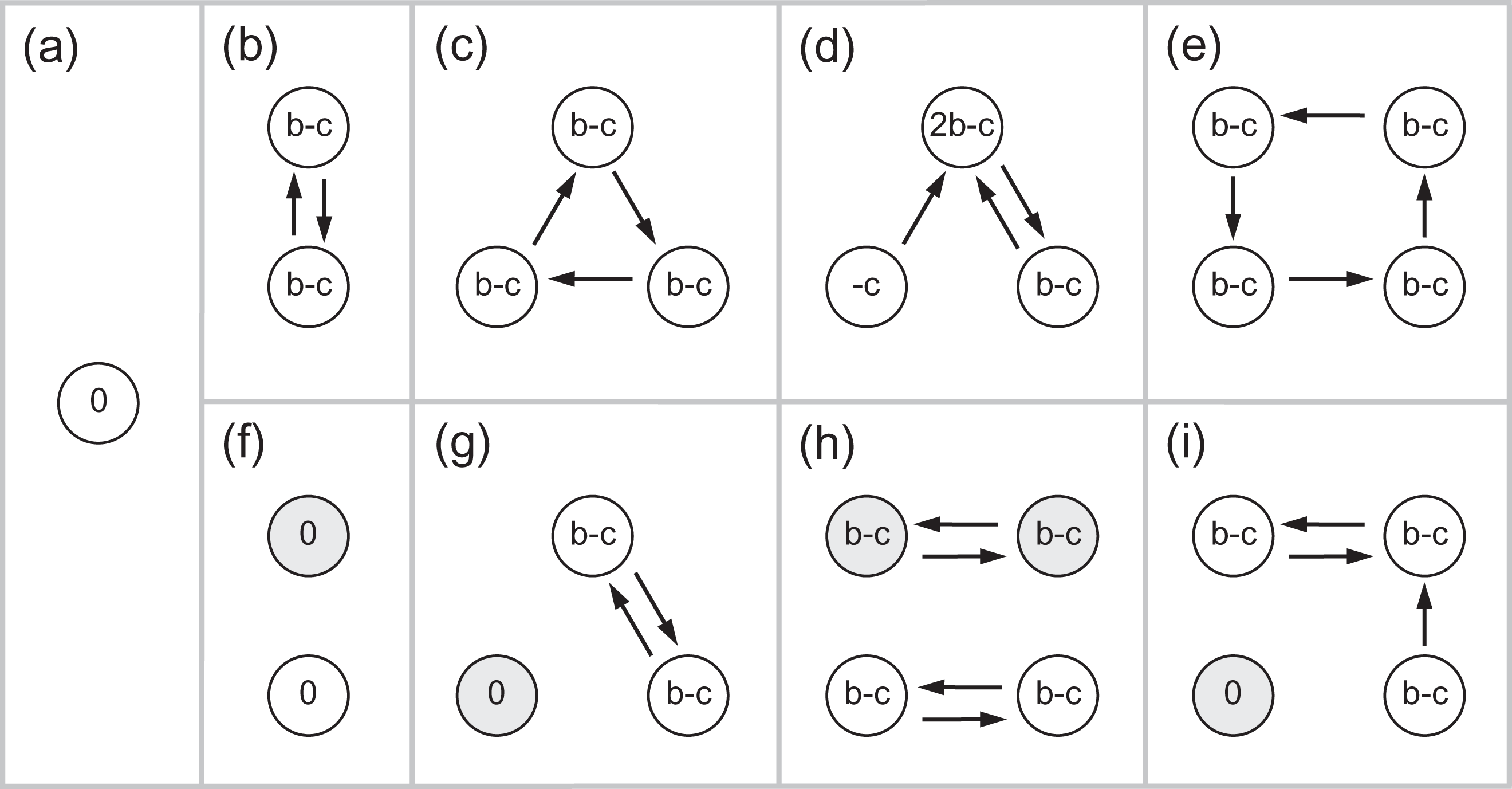
Some possible game interactions between agents within small (early stage) tag groups. Circles represent agents and indicate payoffs, arrows donations, shading represents ethnicity. (a–e) Show ethnically homogenous groups and (f–i) show mixed groups. Mixed groups constrain interactions when combined with a both selector (shown). For cultural selector groups interactions are not constrained since ethnicity is ignored.
Recall that in the baseline runs (comprising only one ethnicity), the cultural and both selectors are equally successful because they are functionally equivalent. Similarly, they are functionally equivalent within an ethnically homogenous tag group. But there is no mechanism for sorting ethnicities between tag groups, so groups that grow always become ethnically well mixed. But in the early stage of a group, when it is of small size, there is a reasonable probability that it will be ethnically homogenous by chance. An ethnically homogenous both selector group can compete equally (producing the same number of donations) as a cultural selector group. This can be considered as a form of noise that inhibits donation because ethnicities are effectively randomly distributed between groups.
Once a group has at least two of each ethnicity, a both selector will produce the same number of donations as a cultural selector since each agent can find an in-group member of the same ethnicity to donate to. Notice that the both selector group in Figure 3b is ethnically homogenous for the first four cycles, and in cycle 5, it is split between both ethnicities.
Given the early-stage interaction possibilities, it is evident that both selector groups are more likely to be successful (and grow) if they are ethnically homogenous at the early stage. This can be seen in Figure 5, comparing the ethnic homogeneity of cultural and both groups during their early stages. This clearly illustrates that successful both groups (in terms of reaching a size 10 or greater) are associated with higher levels of ethnic homogeneity than similarly successful cultural groups.
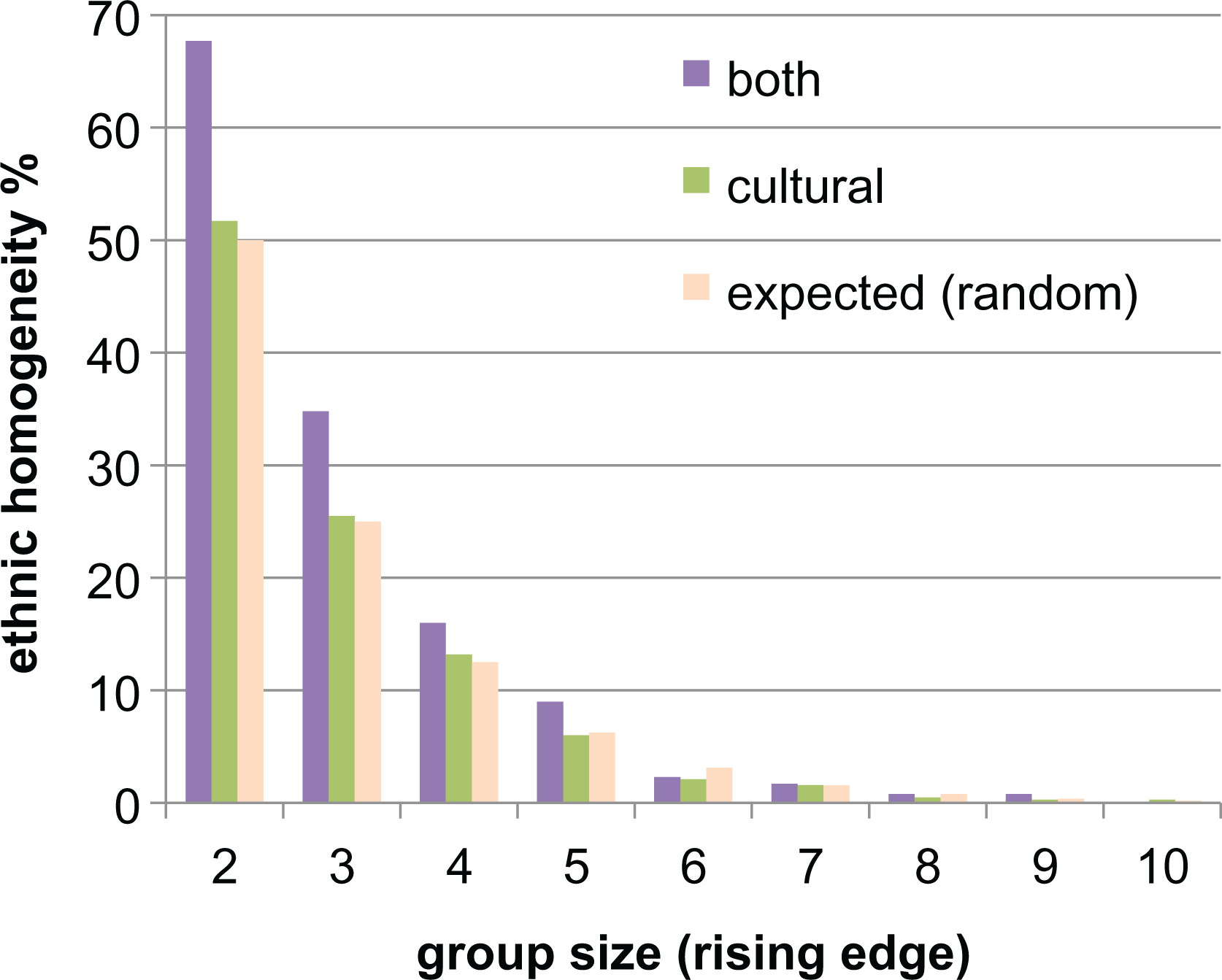
Early-stage ethnic homogeneity for groups reaching size ≥ 10 agents, shown for each stage of initial (rising edge) group size from 2 to 10. Notice that almost 70 percent of both selector groups were ethnically homogeneous for size = 2 (compared to an expected 50 percent from random mixing), indicating that early-stage ethnic homogeneity is associated with the growth of both selector groups. Calculated over 3,000 cycles for the number of ethnicities = 2.
Most tag groups never grow beyond small size. The majority of groups (≅80 percent) never grow beyond one-seeds and only ≅10 percent reach a size of at least ten agents. However, the majority of game interactions occur within larger groups. This can be seen from the distribution of group sizes and associated areas shown in Figure 6.
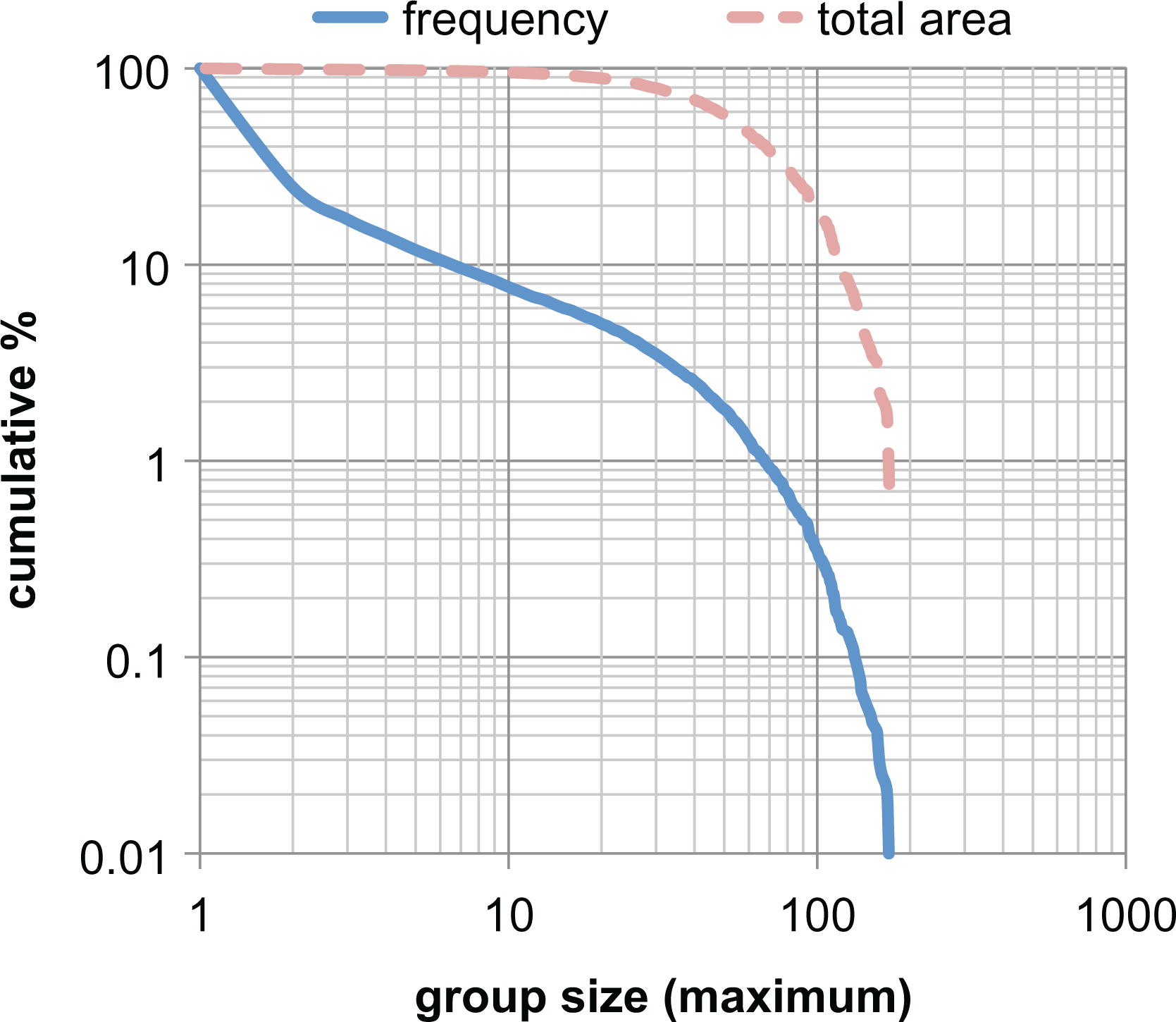
Cumulative distribution of tag group sizes and their total area for a simulation run. Size indicates the maximum number of agents that a group reaches and area indicates the sum of the size of the group for each cycle of its existence. Calculated over 3,000 cycles for number of ethnicities = 2. Results comprise approximately 3 × 10 4 groups with a total area of 3 × 10 6 .
The area of a group measures the sum of all sizes of a group over its entire existence. Hence, a one-seed that existed for only one cycle would have area = 1. A group that lasted three cycles starting as a one-seed, then size 2 and finally size 3 would have area = 6. Thus, the areas of the groups in Figure 3 are 249 and 363, respectively. Figure 6 shows the cumulative total area for all groups reaching a given maximum group size (or above). Note that only about 10 percent of groups reach a size of 10 or more, but almost all area (and hence game interactions) occur within such groups. Notice also that only about 0.3 percent of groups reach size 100 but they comprise 20 percent of the total area. The age of groups (not shown) follows a similar distribution with about 5 percent lasting >50 cycles and only about 0.1 percent reaching >250. No group gets beyond an age of 500 cycles.
Large groups have the side effect of producing many new one-seeds through mutation on the tag—thus potentially reproducing a new copy of the group. Specifically, a group will produce area ×10M new seeds over its lifetime because the tag mutation rate is 10M. For M = .001, this equates to area/100 new seeds. Hence, large groups can be viewed as engines of seed production spawning new copies of themselves. This leads to a positive feedback process that biases the production of new one-seeds toward currently successful selectors. In this context, although both selector groups are less likely to grow, if large groups do form and come to take over a significant proportion of the population, then there is a lock-in process because cultural seed production is inhibited.
Further Experiments
We performed further experiments exploring parameters beyond the ranges given in Table 2. We summarize these briefly. For details, see the Online Appendix in Supplemental Material (see also Hales and Edmonds 2015).
Populations with more than two ethnic markers
Populations equally split between four ethnicities (NE = 4) produce sb ≅ 4 percent on average with low variance. Hence, interethnic donation (ie) is 4 percent lower than would be expected if no ethnic discrimination was occurring. This attenuated result arises because there is less chance of ethnically homogenous early-stage groups forming and of groups growing to include at least two agents of each ethnicity. Similarly, for NE = 8, sb is further attenuated (≅ 2 percent). Hence, the fewer effectively perceived ethnicities, the more likely are bursts of ethnocentrism.
Different population sizes
We ran experiments over a range of population sizes with two ethnicities. Small populations (N < 500) produced a high average sb (about 40 percent) with high variance between different runs—some runs produced sb values > 90 percent resulting in very low interethnic donation rates (ie). This occurs because small populations allow for large both selector groups (if they form) to take over the majority of the population and thus “crowd out” the production of cultural selector seeds due to the positive feedback process discussed previously. The same is also true for cultural selectors so large variances are observed between runs.
With higher values of N, the variance and average sb decreases converging to a value of sb ≅ 10 percent when N >> 1,000. This means that for larger populations, interethnic donation is about 10 percent less than would be expected with no discrimination. This indicates that both selector groups can still form and reduce interethnic donation but do not come to dominate the population. We tested populations up to N = 12,800 but do not have a proof that sb values would maintain this level as N → ∞.
Varying game and imitation partner selection
Our model assumes game interactions occur strictly within the in-group, whereas imitation is population-wide. When we relaxed this assumption by replacing this “all-or-nothing” partner selection approach with probabilistic rules, we found that similar results were produced so long as there was a significantly higher probability of in-group game interactions as compared to imitation interactions.
Varying the benefit to cost ratio
We experimented with different B/C ratios. We found that in general, we get comparable results when the benefit is greater than the cost (i.e., when B/C > 1).
Observations and Interpretations
Here, we make some general observations based on the results. We then make some points concerning the interpretation of the model.
Observations
High donation rates occur between agents sharing the same cultural tag
This is consistent with previous evolutionary tag models of cooperation and is not a novel result. High donation rates occur due to the dynamic formation and dissolution of in-group cooperative cultural tag groups with agents defining their in-groups with reference to these. The cooperation that emerges is due to generalized exchange occurring within groups (Takahashi 2000) and not due to direct reciprocation of donations (which is impossible here).
Cultural tag groups trump ethnic groups and no pure ethnocentrism emerges
The majority of agents come to ignore the ethnic marker in defining their in-groups. In all cases considered, agents do not come to define their in-group with reference to their ethnicity alone. Hence, “pure” ethnocentrism does not emerge.
A form of ethnocentrism does emerge based on in-groups defined by both the cultural tag and ethnic marker combined
In some circumstances, this may dominate the population leading to periods of very low interethnic donation rates, yet high overall donation rates. Such breakdowns in interethnic donation are contingent and unpredictable yet reversible (as shown in Figure 2 above). Thus, for significant periods of time, a population can evidence hardly any interethnic donation activity. An observer in such an artificial society would occasionally see hard-to-explain rapid short-term transitions from high interethnic donation to low and vice versa.
Ironically, it is the success of the cultural tag processes that allows for the promotion of discrimination based on ethnic markers and cultural tags. This occurs when small early-stage tag groups happen to be ethnically homogeneous through random variation in their composition. In this situation, discrimination on both ethnic marker and tag is functionally equivalent to tag only discrimination. Such groups grow and soon become ethnically well mixed but still define their in-group with reference to both ethnic marker and tag. Within such groups, donation only occurs between agents sharing the same ethnicity.
Interestingly, we found that this process occurs mainly through short-lived groups of comparatively small size, although a small number of longer lasting and larger groups serve to sustain the creation of those groups. However, overall we do not find any groups that last more than 500 simulation cycles. We also found that ethnocentrism attenuates with >2 ethnicities and with population sizes >>1,000, yet still persists.
Interpretation
The purpose of our model is that of theoretical exploration. It does not attempt to model the observed world in any direct sense but rather should be viewed as a thought experiment using an artificial society. It can suggest hypotheses about the observed world but does not, alone, prove anything about it. 21
The model assumes that within-generation adaptation, based on imitation and innovation toward improving individual benefit, are the only processes that determine agent behavior change. 22 The purpose is to establish the sufficiency of these mechanisms for obtaining the described outcomes. Other mechanisms might well reinforce or frustrate the ones described here. Thus, the point of this model is to be able to separate out chosen mechanisms in a way not possible in observed cases in real societies.
In our model, all agent traits, apart from the ethnic markers, are fluid and may be easily changed. We ignore social networks, other relationships between agents, and wider historical or cultural phenomena. Agents have no memory of past encounters and cannot recognize individuals. They can only distinguish others based on the ethnic marker and the cultural tag—nothing else.
People in real societies do not change their beliefs and behaviors based on simple imitation or random changes. People are attached to particular beliefs and practices for many reasons other than individual benefit. In fact, beliefs and practices may be the basis of identity itself and be held even when they are of no benefit to the individual at all—perhaps, even because they are of no benefit. Social behaviors and beliefs result from a complex interplay of upbringing, personal experience, social expectations, and norms and are not purely the result of adaptation of strategy and the definition of who is the “in-group.”
Also, real-world groups often have specific barriers to entry (and exit) depending on their nature. One cannot immediately enter or exit them costlessly. Entry may involve vetting procedures, a trial period, or some other cost. Exit may involve a penalty, loss of opportunity, or other sanction.
We view our model in a similar way to the famous segregation model of Schelling (1971). This showed that even if agents had no intolerant predispositions, their interactions produced high levels of spatial segregation based on fixed markers. Our model shows that even when agents have no specific in-group predispositions or discriminatory behaviors, their interactions can lead to discrimination based, partially, on fixed markers.
Yet the results from our model do not appear to relate to the traditional concept of ethnocentrism as widely used within the social sciences. Specifically, our model produces ethnocentric behavior through short-lived, small, highly dynamic groups where there are only two distinguishable ethnicities rather than long-run entrenched group identities.
Future Work
The model, although comparatively simple, evidences rich behavior that we have only explored one aspect of. Other future work could include: Introducing migration such that new ethnic minority agents arrive at some fixed of variable rate. Combining long-run generational vertical evolution (considered in previous models) with short-run cultural (horizontal) evolution considered in our model.
Conclusions
In our simulation experiments, we found no cooperative groups based on the ethnic marker alone. Also, the amount of intercultural donation was insignificant in all variants explored. Anything that could be interpreted as pure ethnocentrism did not arise. Ethnocentrism, when it did arise, occurred in conjunction with cultural discrimination. Unlike previous models (that follow Hammond and Axelrod 2006), our model showed how short-term horizontal cultural processes might explain the appearance of ethnocentrism, without a tendency for preferring one’s own ethnicity being “hardwired” by biological or, other forms of, vertical evolution.
Our model is consistent with the conclusions of Yamagishi, Jin, and Kiyonari (1999), in that in-group favoritism, in terms of distribution of resources, emerges from processes of generalized exchange rather than being linked directly to group identity. In our model, successful groups are where generalized exchange is working and hence attract new members via imitation. The agents in the model do not have expectations, but if they did, their experience might well have resulted in an expectation that in-group favoritism is a suitable strategy.
The hypothesis suggested by this work, that ethnic-based discrimination may result from within-generation in-group formation, has a number of interesting implications. It would mean that ethnocentrism, when it occurs, may be the result of a short-term process that just happens to use ethnic markers as part of the in-group definition rather than being purely based on them. 23
This does not mean that genetic (or other forms of vertical) evolution has played no role, but that we might have evolved to flexibly determine what our in-group is during our lifetime. In our model, horizontal cultural evolution spreads in-group definitions and strategies that favor the in-group over the out-group. Empirical experiments have shown that people given arbitrary group assignments come to act favorably to the in-group and less so to the out-group (Brewer 1979). That work clearly demonstrates that no imitation or innovation phase is required. Simply telling individuals they are part of a group produces the in-group behavior. This implies that humans are predisposed to favor any in-group so long as they know how it is defined. It is sufficient for an experimenter to give them their group assignments. Hence, it appears that people are not a priori fixated on a particular in-group definition but that this is fluid. Perhaps, slower forms of vertical evolution (including genetic evolution) could select for in-group favoritism irrespective of the group definition—as might be suggested by the “social intelligence hypothesis” of Kummer et al. (1997). In other words, genetic evolution might have adapted to the selective advantage of group cooperation by giving us this ability. Our general “groupishness” as a species (Ridley 1997) may have evolved in the past but not our salient groups in the present because they are culturally constructed and may change rapidly. In this context, ethnocentrism might be viewed as merely one possible expression of inherent groupishness but not a long term evolved phenomena.
Supplemental Material
Supplemental Material, JCR-16-0494.R1 - Intragenerational Cultural Evolution and Ethnocentrism
Supplemental Material, JCR-16-0494.R1 for Intragenerational Cultural Evolution and Ethnocentrism by David Hales and Bruce Edmonds in Journal of Conflict Resolution
Footnotes
Acknowledgments
We wish to thank the anonymous referees for their encouragement and significant effort in helping us to improve the clarity and focus of the work.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was partially funded by the UK EPSRC as part of the “Social Complexity of Immigration and Diversity” project, Grant Number EP/H02171X.
Supplemental Material
Supplementary material for this article is available online.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.