
Abstract
Objective:
High doses of opiate substitution pharmacotherapy are associated with greater treatment retention and lower illicit drug consumption, although the neurobiological bases of these benefits are poorly understood. Dysfunction of the anterior cingulate cortex (ACC) is associated with greater addiction severity and mood dysregulation in opiate users, such that the beneficial effects of substitution pharmacotherapy may relate to normalisation of ACC function. This study aimed to investigate the differential impact of methadone compared with buprenorphine on dorsal ACC biochemistry. A secondary aim was to explore the differential effects of methadone and buprenorphine on dorsal ACC biochemistry in relation to depressive symptoms.
Methods:
Twenty-four heroin-dependent individuals stabilised on methadone (n=10) or buprenorphine (n=14) and 24 healthy controls were scanned using proton Magnetic Resonance Spectroscopy and compared for metabolite concentrations of N-acetylaspartate, glutamate/glutamine, and myo-inositol.
Results:
(1) Methadone was associated with normalisation of dorsal ACC biochemistry (increased N-acetylaspartate and glutamate/glutamine levels, and decreased myo-inositol levels) in a dose-dependent manner; (2) buprenorphine-treated individuals had higher myo-inositol and glutamate/glutamine levels than methadone-treated patients in the right dorsal ACC; and (3) myo-inositol levels were positively correlated with depressive symptoms in participants stabilised on buprenorphine.
Conclusions:
These findings point to a beneficial role of high-dose methadone on dorsal ACC biochemistry, and suggest a link between elevated myo-inositol levels and depressive symptoms in the context of buprenorphine treatment.
Introduction
Opiate substitution treatment is a well-established and effective approach for the management of opiate dependence (Amato et al., 2005; Mattick et al., 2008, 2009), with stabilisation on methadone and buprenorphine associated with greater rates of treatment completion, reduction of illicit drug use and improvements in social and health status. Methadone is a full agonist with high binding affinity at mu opiate receptors, whereas buprenorphine is a partial agonist at mu receptors and an antagonist at kappa opiate receptors (Tetrault and Fiellin, 2012). Both methadone and buprenorphine have superior beneficial effects on retention rates and illicit drug use when given in higher doses, with direct meta-analytical comparisons showing either similar effects of high doses of both drugs (Barnett et al., 2001; Farré et al., 2002) or mildly superior effects of methadone in both retention rates and illicit drug consumption (Mattick et al., 2009; West et al., 2000). Whilst high doses of methadone and buprenorphine are associated with increased treatment retention and reduction of illicit drug taking (Amato et al., 2005; Barnett et al., 2001; Farré et al., 2002), there have been concerns about dose-related detrimental effects on brain health and cognitive performance (Curran et al., 2001; Mintzer et al., 2004; Tramullas et al., 2007). We recently demonstrated that opiate-dependent individuals stabilised on substitution pharmacotherapy have functional abnormalities and decreased concentrations of glutamate/glutamine (Glx) and N-acetylaspartate (NAA) metabolites in the dorsal anterior cingulate cortex (dACC) (Yücel et al., 2007). While ACC biochemistry has been associated with indices of addiction severity (e.g. number of previous withdrawals) among opiate-dependent individuals (Hermann et al., 2012), no previous study has examined the relationship between opiate substitution dose and cingulate biochemistry, nor examined whether there are differential effects between methadone and buprenorphine.
The dACC is also thought to play a key role in the neurocircuitry of mood regulation (Phillips et al., 2003). There is evidence of reduced dACC volume in depressed patients (Vasic et al., 2008), and of a unique link between increased dACC grey matter (GM) and a greater ability to regulate negative affect (Giuliani et al., 2011). Previous studies indicate that methadone and buprenorphine may have opposing effects on mood regulation by virtue of their differential impact on brain function. For example, an association was demonstrated between reduced ACC metabolism and dysthymia scores among heroin users, but this link was not evident in patients stabilised on methadone, suggesting that methadone may improve mood (Galynker et al., 2007). Furthermore, peak methadone concentrations have been associated with significant reductions in depressive reactivity in methadone-treated heroin users (Savvas et al., 2012). Conversely, we recently found that pituitary gland volume was correlated with depression scores among individuals treated with buprenorphine but not methadone (Lorenzetti et al., 2010), suggesting that buprenorphine may not be as effective as methadone in normalising brain systems involved in stress reactivity. These findings are also consistent with clinical data demonstrating that buprenorphine is less effective than methadone in reducing illicit drug use among depressed heroin users (Kosten et al., 2004).
The primary aim of this study was to investigate the differential impact of methadone and buprenorphine dose on cingulate biochemistry. We hypothesised that higher doses of methadone and buprenorphine would normalise dACC biochemistry. A secondary aim was to explore the differential effects of methadone and buprenorphine on dACC biochemistry in relation to depressive symptoms. We hypothesised that participants on buprenorphine, but not methadone, would demonstrate a relationship between abnormal dACC biochemistry and depressive symptoms.
Materials and methods
Participants
Twenty-four opiate dependent individuals and 24 healthy controls participated in the study; sample characteristics have been previously described (Yücel et al., 2007) Opiate-dependent individuals were stable on methadone (n=10) or buprenorphine (n=14), with each group statistically matched on age, years of education, full IQ score, duration of opiate use, severity of dependence and depression and anxiety scores (see Table 1). Patients were screened using the patient edition of the Structured Clinical Interview for DSM-IV Axis I Disorders (First et al., 2001) to ensure that they did not meet criteria for a current anxiety, mood, or psychotic disorder. Some opiate-dependent participants were still using other drugs on a weekly basis over the preceding 3 months (n=3 cannabis, n=4 benzodiazepines and n=2 heroin within the methadone subgroup; and n=2 cannabis, n=2 benzodiazepines, and n=5 heroin in the buprenorphine group). Importantly, none of these participants used psychostimulant drugs (cocaine, amphetamine or ecstasy) on a regular basis. Irrespective, all subjects were required to abstain from using any substance for at least 24 h prior to the scanning session, although they were permitted to take their usual morning opiate prescription. Beck Depression Inventory (BDI) scores ranged between 0 and 29 (M=12, SD=12.4) in the methadone-treated group, and between 0 and 31 (M=13.9, SD=10.7) in the buprenorphine-treated group. Methadone doses ranged between 5 mg and 150 mg (M=41.4, SD=43.5), whereas buprenorphine doses ranged between 1 mg and 40 mg (M=10.2, SD=10.9). Duration of therapy ranged between 8 and 124 months (M=47, SD=43.14) in the methadone-treated group, and between 3 and 64 months (M=20.28, SD=16.78) in the buprenorphine-treated group; differences in treatment duration were not significant between the groups. All subjects provided informed consent and the study was approved by the local hospital ethics committee.
Demographic and clinical characteristics of participants treated with methadone and buprenorphine.
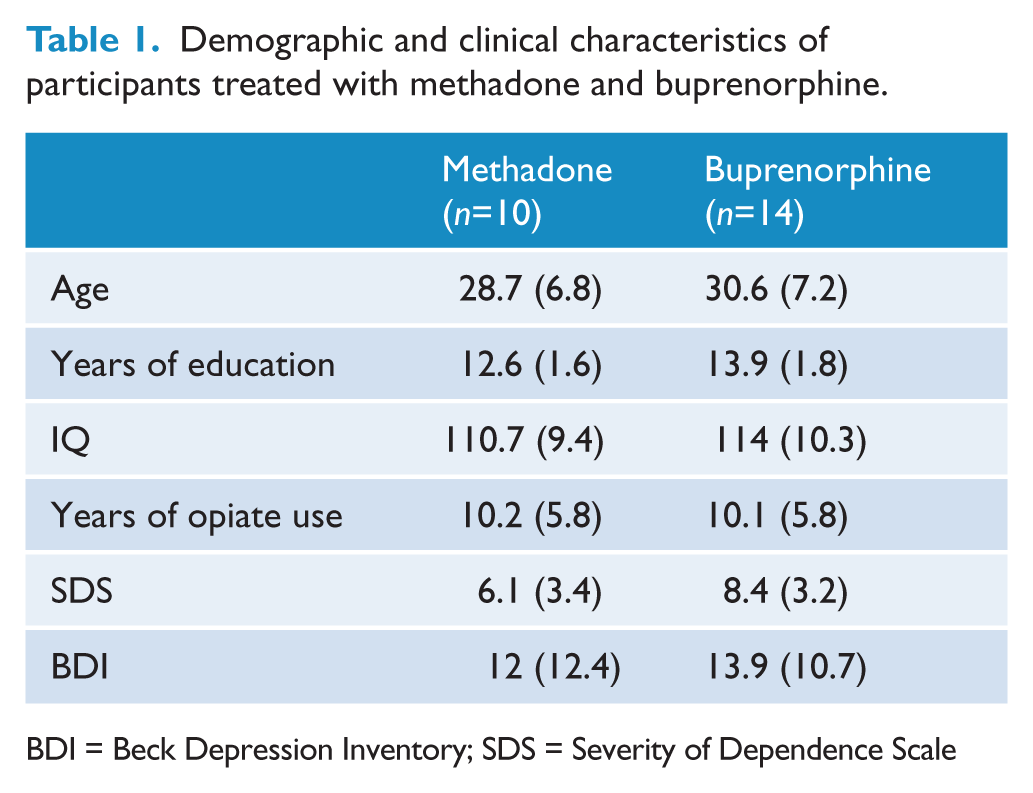
BDI = Beck Depression Inventory; SDS = Severity of Dependence Scale
Magnetic resonance imaging acquisition and determination of metabolite concentrations
All individual magnetic resonance imaging (MRI) sequences were acquired in a single scanning session using a 3T GE Signa LX whole body scanner (GE Healthcare, Milwaukee, WI, USA). Volume localised proton magnetic resonance spectroscopy (1H-MRS) was recorded using a standard short-echo point resolved spectroscopy sequence (PRESS; TR=3000 ms, TE=30 ms, NEX=128; with a nominal voxel size ~6.5 cm3) from the dACC bilaterally. MRS voxels were placed in each hemisphere, separated by the medial wall boundary of the frontal lobes, so as to encompass the dACC. The posterior boundary of the voxel was ~10 mm posterior of a vertical line from the anterior commissure (AC) orthogonal to the AC–PC (posterior commissure) line. The inferior border was ~5 mm superior to the top edge of the corpus callosum while the medial border was 1–2 slices lateral to the parasagittal slice on the T1 structural image for each hemisphere. For volume-localised spectra, absolute levels of N-acetyl compounds (NAA + NAAG), myoinositol (mI), and Glx were measured, as well as choline-containing (Cho) and creatine + phosphocreatine (Cr) metabolites (see Table 2), although the latter metabolites were not of primary interest to the current study.
Dorsal anterior cingulate cortex metabolite concentrations in healthy controls, methadone- and buprenorphine-treated participants.
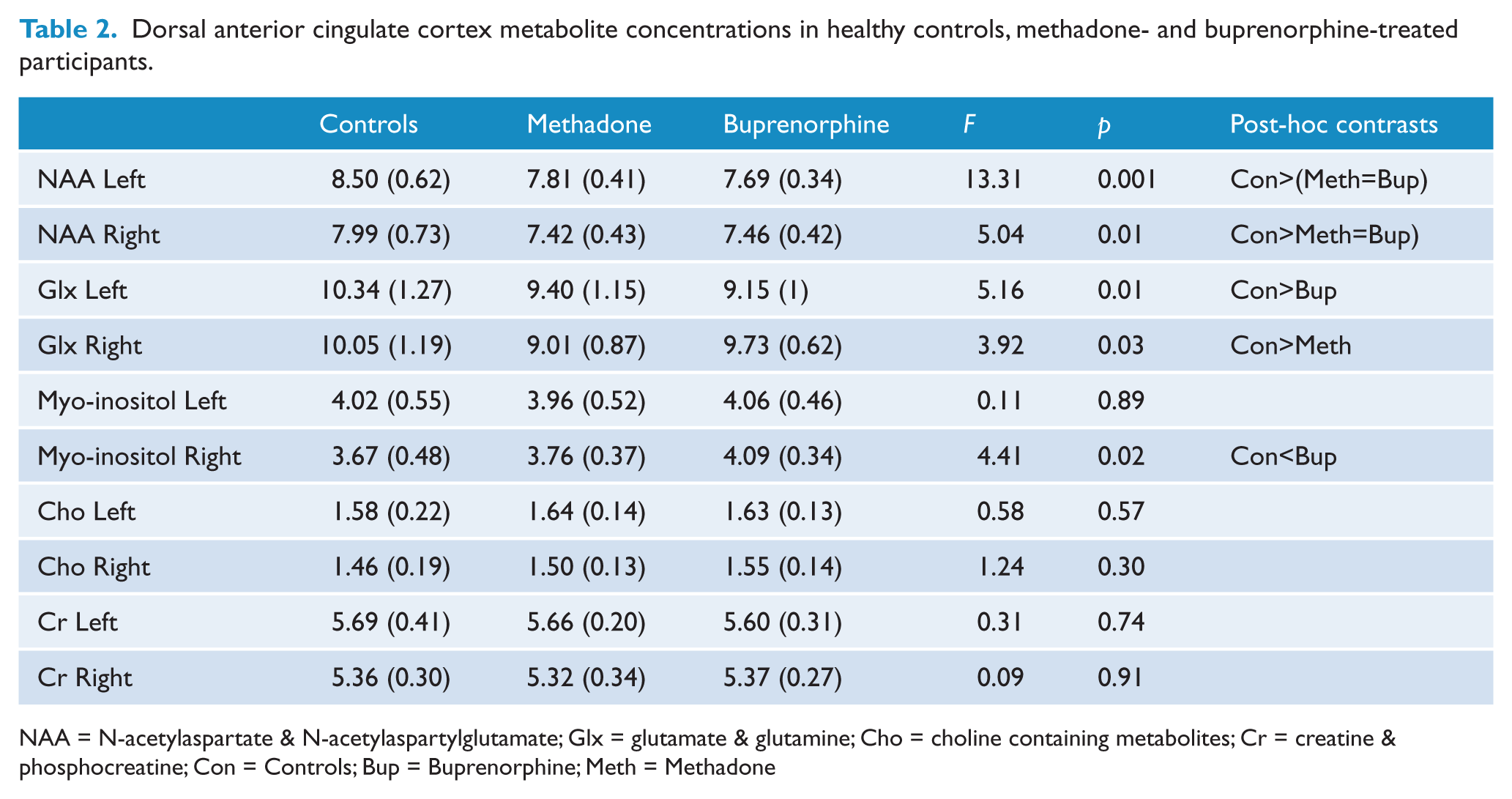
NAA = N-acetylaspartate & N-acetylaspartylglutamate; Glx = glutamate & glutamine; Cho = choline containing metabolites; Cr = creatine & phosphocreatine; Con = Controls; Bup = Buprenorphine; Meth = Methadone
The metabolite content of voxels was corrected for differences in tissue composition using segmentation information, derived from coronal 3-D spoiled gradient-recalled echo in the steady state images, recorded during the same scanning session. The brain component was separated from non-brain in the structural images, using the Brain Extraction Tool of FSL (www.fmrib.ox.ac.uk). The images were then segmented into GM, white matter and cerebrospinal fluid using the FAST module of FSL. The proportion (probability) for each of the three tissue types was then determined for the volume and position of each MRS voxel, using a module (Masterton et al., unpublished data, 2005) written in IDL for iBrain (www.brain.org.au/software.html). Where there was a correlation of 0.2 (Spearman’s rank-order correlation coefficients, two-tailed) for the amount of GM in the voxel and any metabolite, we corrected for the influence of GM using a previously described formula (Free et al., 1995), cMRS=mMRS-(gradient [mGM-meanGM]), where cMRS is corrected MRS; mMRS is the measured MRS concentration; gradient is the gradient of the regression line between the GM tissue volume of interest and the average amount of GM in the entire sample of subjects; mGM is the measured GM fraction; and mean GM is the mean GM tissue volume of the entire sample. These corrected metabolite concentrations were used for subsequent analyses. Results are presented in institutional units approximating millimolar (ppm) concentration. The residual signal corresponds to, and is fitted by, additional broad peaks representing unknown metabolites and other factors such as macromolecular components with short T1 relaxation times. The MRS parameters used for this study provided robust signals both for the healthy control and both the methadone and buprenorphine groups, with a signal-to-noise ratio of 16.12 (SD=2.23), 17.60 (SD=2.26) and 15.93 (SD=2.23), respectively, and a FWHM of 0.06 ppm (SD=0.01) for all three groups. The output from LCModel also includes the Cramér–Rao lower bounds (CRLB), which is a measure of reliability of the fit. The mean CRLB for NAA, mI and Glx were 5.3, 8.4 and 9.9, respectively, for healthy controls, 5.1, 7.1 and 9.6, respectively, for methadone participants and 5.9, 7.1 and 9.3, respectively, for the buprenorphine group.
Statistical analysis
We performed a one-way ANOVA, followed by post-hoc Bonferroni tests, to examine possible differences between the three groups (methadone, buprenorphine and controls) on levels of NAA, myo-inositol and Glx in the dACC. Since our main interest was to capture potential differences in MRS-indexed metabolite levels between methadone-treated and buprenorphine-treated individuals, we also performed paired t-tests to examine differences in metabolite concentrations between the two groups. Mixed-design repeated-measures ANOVAs (2 Hemisphere * 3 Group) were used to examine differences between left and right metabolite concentrations as a function of group. Finally, following our initial hypotheses, we conducted Spearman Rho correlation analyses including methadone vs. buprenorphine doses, MRS-indexed metabolite levels and depression scores. Two individuals (one from the methadone subgroup and one from the buprenorphine subgroup) were excluded from these correlation analyses because they had dose levels higher than 2 SD above the rest of their treatment subsample (i.e. 150 mg methadone and 40 mg buprenorphine).
Results
Group differences in metabolite concentrations between individuals on methadone vs. buprenorphine pharmacotherapy
Group comparisons revealed that buprenorphine-treated individuals (but not methadone-treated individuals) had higher right dACC myo-inositol levels (p=0.016) and lower left dACC Glx levels (p=0.014) than healthy controls. On the other hand, methadone-treated individuals (but not buprenorphine-treated individuals) had lower right dACC Glx levels (p=0.023) than healthy controls (Table 2). Planned t-test paired comparisons between methadone and buprenorphine subgroups showed that buprenorphine-treated individuals had higher right dACC myo-inositol (t=−2.28, p=0.032) and Glx levels than methadone-treated individuals (t=−2.38, p=0.027). In addition, the repeated-measures ANOVAs showed a Group*Hemispheric interaction effect for dACC myo-inositol concentrations, F(2,45)=3.63, p<0.05. The subsequent univariate ANOVAs showed this effect was driven by the lack of significant differences between left and right myo-inositol concentrations in the methadone (p=0.14) and especially the buprenorphine group (p=0.81), whereas controls had left higher than right values (p<0.001). No other Group*Hemispheric interactions were observed for the other metabolites.
Correlations between pharmacotherapy, depressive symptoms and metabolite concentrations
Increased methadone dose was significantly associated with decreased mean dACC myo-inositol levels (r=−0.73, p=0.039) and increased levels of NAA (r=0.76, p=0.03) and Glx (r=0.71, p=0.05) (see Figure 1). In contrast, buprenorphine dose did not correlate with any metabolite level measure. However, higher myo-inositol levels were significantly correlated with higher depressive scores (r=0.56, p=0.048) within the buprenorphine group (see Figure 2). There were no significant correlations between dACC biochemistry indices and severity of opiate dependence, duration of opiate use, or duration of substitution pharmacotherapy.
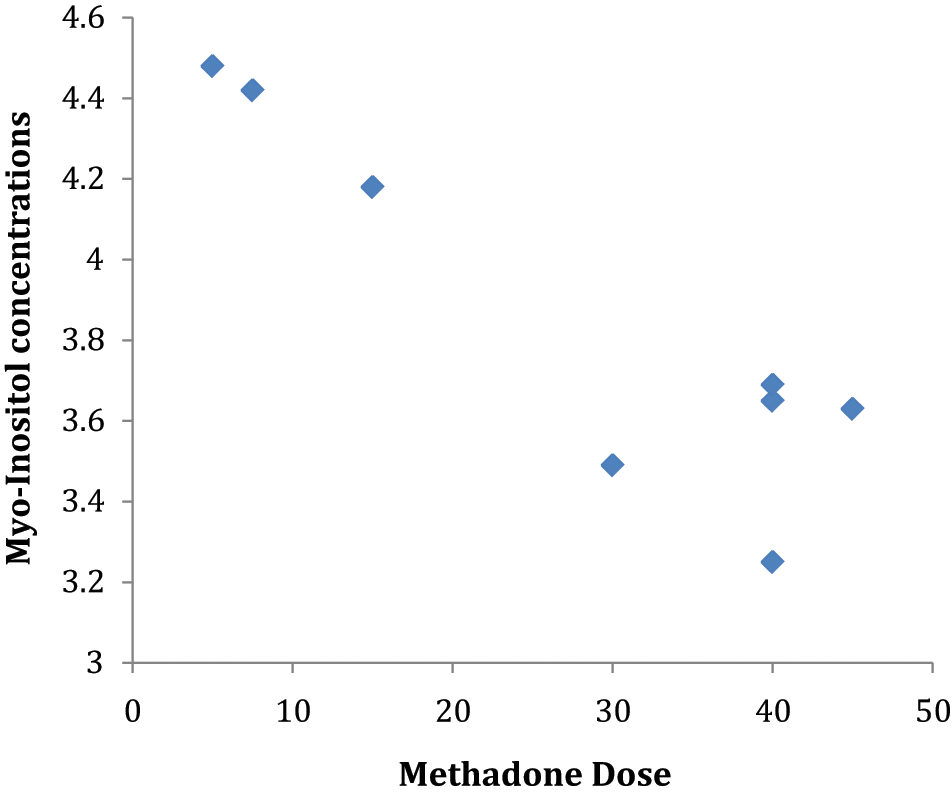
Correlation between methadone dose and myo-inositol within the methadone-treated group.
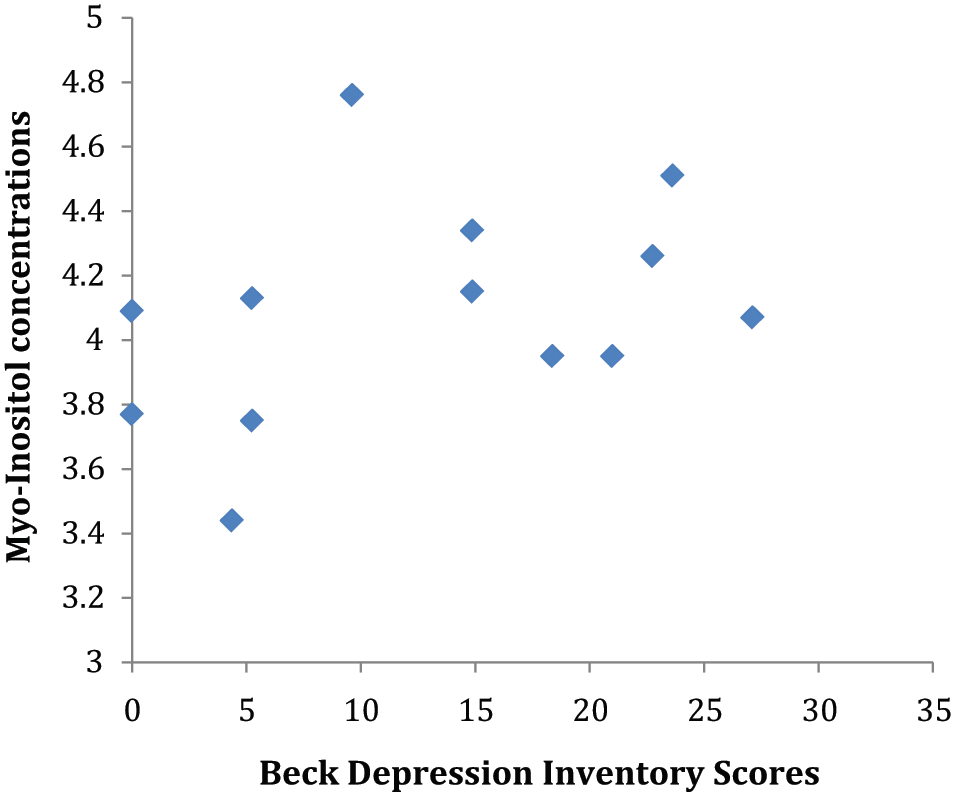
Correlation between Beck Depression Inventory (BDI) scores and myo-inositol within the buprenorphine-treated group.
Discussion
The present study yielded two major findings: (1) higher methadone doses are associated with normalisation of dACC biochemistry in heroin-dependent individuals (decreases in myo-inositol, and increases in NAA and Glx levels); and (2) buprenorphine treatment is associated with increased dACC myo-inositol levels, which are positively correlated with depressive symptoms. Our first finding points to a beneficial role of high-dose methadone on dACC biochemistry, and provides neurobiological support for treatment studies that highlight high-dose methadone is associated with better treatment outcomes (Amato et al., 2005; Barnett et al., 2001; Farré et al., 2002). Previous neuroimaging studies have found that methadone treatment helps normalise brain phospholipid balance (Kaufman et al., 1999), as well as frontal cortical and ACC regional metabolism (Galynker et al., 2007). Behavioural studies have also demonstrated that methadone treatment normalises error detection during decision-making, one of the main functions of the ACC (Ersche et al., 2005). The novelty of our findings is to demonstrate a dose-related effect of methadone in normalising the biochemistry of a brain region that is important for changing one’s drug use (Romero et al., 2010), as well as cognitive control of craving and relapse (Paulus et al., 2005; Volkow et al., 2010).
It is important to note that in the case of Glx there was an apparent discrepancy between group comparisons (where the buprenorphine group demonstrated higher Glx levels) and correlation analyses (where methadone dose seemed to normalise Glx functioning). These findings raise the possibility that only higher doses of methadone contribute to normalisation of dACC glutamate function. However, the significance of fluctuations in Glx levels within opiate dependence is still debated; for example, recent evidence indicates that Glx levels are associated with the number of previous withdrawal episodes in opiate users on substitution pharmacotherapy (Hermann et al., 2012). As such, more research is needed to establish the relevance of Glx concentrations in relation to glutamate alterations and clinical phenotypes among opiate users seeking treatment.
Our second finding demonstrates a link between buprenorphine treatment, elevated dACC myo-inositol and depressive symptoms. Elevations in myo-inositol levels are thought to represent increased glial hypertrophy and/or proliferation (Danielsen and Ross, 1999). These elevations have been observed across different groups of addicted individuals, including amphetamine, ecstasy, cocaine, toluene and alcohol users, and have been interpreted in terms of reactive processes occurring in response to drug-induced neuroadaptations (Licata and Renshaw, 2010). Consistent with this notion, animal models have demonstrated opiate treatment-induced myo-inositol elevations in brain regions rich with opioid receptors, which tend to subside after drug cessation (Gao et al., 2007; Xiang et al., 2006). Because the co-use of other drugs (particularly psychostimulants) is minimal in our sample, and both subgroups (methadone- and buprenorphine-treated participants) were matched on severity of opiate dependence, the dACC myo-inositol finding appears to specifically relate to treatment with buprenorphine. Indeed, an increased myo-inositol level in buprenorphine-treated individuals (compared with the methadone group) suggests that buprenorphine is less able to restore prior opiate-induced neuroadaptations. However, given that this effect was restricted to the right dACC, further studies are required to confirm and better understand the clinical significance of this finding.
The positive correlation between dACC myo-inositol levels and depression scores support the view that dACC dysfunction is associated with poor mood regulation (Giuliani et al., 2011; Vasic et al., 2008). This result may help explain the clinical finding that buprenorphine treatment (compared with methadone) is less effective in reducing illicit drug consumption among depressed heroin users (Kosten et al., 2004). This result is also consistent with our previous finding linking pituitary gland volumes with depression scores among individuals treated with buprenorphine (Lorenzetti et al., 2010). Alterations in the function of both the ACC and the hypothalamic–pituitary–adrenal axis are thought to underlie abnormal stress regulation in addicted individuals (George and Koob, 2010; Li and Sinha, 2010). Combined evidence from both studies suggests that buprenorphine treatment may be less able to normalise brain systems relevant to mood regulation in long-term heroin users.
A number of limitations may have affected our findings, including the relatively small sample size of the two treatment groups, which increases the risk of Type II errors in both group and correlation analyses. The lack of indices of cingulate opiate receptor occupancy is also a limitation, as such data would clarify the association between opiate substitution dose, its impact on cingulate function (e.g. degree of receptor occupancy), and variations in mood states (Greenwald et al., 2003). Further follow-up studies are warranted to examine if the effects of opiate substitution treatment on brain metabolites are enduring following detoxification.
Footnotes
Funding
The study was supported by the Colonial Foundation and the National Health and Medical Research Council (NHMRC). MY is supported by a NHMRC Senior Research Fellowship Award (I.D. 1021973). AV-G is supported by a José Castillejo Fellowship from the Spanish Ministry of Education and Science (JC2010-0195). RV-L is supported by a José Castillejo Fellowship from the Spanish Ministry of Education and Science (JC2010-0193).
Declaration of interest
The authors report no conflicts of interest. The authors alone are responsible for the content and writing of the paper.