
Abstract
Oestrogen increases facial allodynia through its actions on activation of the MAPK extracellular-signal regulated kinase (ERK) in trigeminal ganglion neurons. This goal of study was to determine which oestrogen receptor is required for behavioural sensitization. Immunohistochemical studies demonstrated the presence of oestrogen receptor alpha (ERα) in nuclei of larger neurons and cytoplasm of smaller neurons, and the novel oestrogen receptor G-protein coupled receptor 30 (GPR30) in small diameter neurons that also contained peripherin, a marker of unmyelinated C-fibres. Specific agonists for ERα (PPT) and GPR30 (G-1), but not ERβ (DPN), activated ERK in trigeminal ganglion neurons
Introduction
Trigeminal pain disorders such as migraine and temporomandibular disorder (TMD) are among the most common neurological conditions (1–3) and are two to three times more prevalent in women than in men (1, 4). The sex disparity in the prevalence of trigeminal pain has been attributed to fluctuations in oestrogen (5), and trigeminal pain is concentrated during the reproductive years, often beginning at menarche and declining after menopause (6). Trigeminal pain varies across the menstrual cycle, increasing during the perimenstrual period (7), and in some cases at mid cycle (8). Although clinical data suggest that oestrogen increases susceptibility to migraine, the fall in serum oestrogen levels at the time of menstruation may precipitate an attack, suggesting a complex relationship between oestrogen levels and the severity of trigeminal pain.
Exogenous oestrogens can also affect nociception in the trigeminal system. For example, oestrogen replacement therapy decreases thermal pain thresholds (9) and may trigger migraine with aura (10). Oral contraceptive use increases the risk of TMD pain (11). These effects may result from oestrogen acting directly on the trigeminal system, which is rich in oestrogen receptors (ERs) (12).
Oestrogen may affect a cell through classical nuclear pathways or by signal transduction cascades initiated at cell surface membrane receptors. These ERs may include either cytoplasmic or nuclear forms of oestrogen receptor α (ERα), ERβ or the more recently described GPR30, a G-protein coupled receptor that has been identified as a novel ER (13–15). In response to oestrogen, GPR30 activates downstream second messenger pathways, including the MAPK extracellular-signal regulated kinase (ERK) (16). ERK activation is a specific marker of nociceptor activation (17) that is required for thermal and mechanical hyperalgesia in several models of neuropathic and inflammatory pain (18, 19). Oestrogen increases facial allodynia through activation of ERK in trigeminal ganglion neurons (20), but the specific receptor mediating these changes has not been determined.
In order better to characterize the effects of oestrogen on trigeminal nociception, we analysed the distribution of GPR30 and the classical oestrogen receptor ERα in the trigeminal ganglion and investigated the effects of selective agonists for ERα or GPR30 on ERK activation and facial allodynia. In order to determine how ERs may be regulated by oestrogen and inflammation, we examined changes in protein expression of ERα and GPR30 in the presence and absence of oestrogen and peripheral inflammation. Our results support the hypothesis that GPR30 is a mediator of oestrogen-related trigeminal nociception and indicate that either GPR30 or ERα can mediate trigeminal sensitization. Furthermore, differential regulation of these receptors by oestrogen and inflammation may contribute to migraine and TMD pain.
Methods
Animals
Female Sprague–Dawley rats aged 60–75 days (Harlan, Indianapolis, IN, USA) were used for all studies. Rats were maintained on a daily 12 h light, 12 h dark schedule with
Ovariectomy
Rats were rapidly anaesthetized with 4% isoflurane in compressed oxygen, and anaesthesia was maintained on 1% isoflurane. Dorsal bilateral incisions were made midway between the lower ribs and the iliac crest, and both ovaries were isolated and removed. Both incisions were closed with suture clips. Animals then received Buprenex (0.1 mg/kg) and were allowed to fully regain consciousness before returning to the animal facility.
Western blot
Trigeminal ganglia were harvested and homogenized in cold lysis buffer (20 m
Immunohistochemistry
Two weeks following ovariectomy, rats were deeply anaesthetized with sodium pentobarbital (50 mg/kg i.p.) and perfused transcardially with 0.1
Microscopy and analysis
Fluorescent digital images were obtained using a Nikon Eclipse 90i upright microscope and a Nikon C1 confocal imaging system using a 20× objective and a frame size of 1024 × 1024 pixels. Sections processed omitting the primary antibody were used to control for non-specific fluorescence. Detector gain and laser intensity were initially set using control slides, and all images were subsequently collected under the same photomultiplier detector conditions and pinhole diameter. In double-label experiments, single-antibody-labelled sections were used to control for channel bleedthrough, and multichannel images were acquired using sequential excitation. For cell counting, individual images were montaged into a single image using Adobe Photoshop CS2 to prevent double-counting at the image edges. Twice the peak intensity of cells processed in the absence of primary antibody was used as the threshold for counting a cell as GPR30+. At least 1000 cells were counted from each of four animals for each condition. At least 100 neurons were measured per trigeminal ganglion section.
Single- and double-labelled neurons were counted from digital images to obtain a percent score for colocalization of GPR30 with each phenotype marker. The percentage of neurons labelled with both markers was determined by counting the total number of neurons containing GPR30 immunoreactivity and identifying the number of cells labelled with the second neuronal marker. The same procedure was repeated to count the Alexafluor 488-labelled cells. We determined the percentage of peripherin and NFH immunoreactive neurons that also contained GPR30 by dividing the number of double-labelled neurons by the total number of neurons that were immunoreactive for each marker. The experiment was repeated using two sets of independent samples.
Cell diameters were measured using the ruler function of Adobe Photoshop CS2. The length of the short and long axes of each neuron in every selected file was measured. The diameter of each cell was calculated as the average of length and width. Data are reported as mean ± standard error, with a
Tissue culture
For each experiment, trigeminal ganglia from four to five cycling female rats were dissected from the base of the skull, rinsed in Hank's balanced salt solution, minced and suspended in digestion medium containing Leibovitz L15 medium with 1 mg/ml BSA, 250 U/ml of CLSPA collagenase, 1 U/ml of ESL elastase, and 5 U/ml of PAPL papain (enzymes from Worthington Biochemical Corp., Lakewood, NJ, USA). An equal volume of 30% Stractan (Larex, White Bear Lake, MN, USA) in Leibovitz's L15 was added to the digest. After mixing by inversion, samples were centrifuged in a swinging bucket rotor at 4 °C or 10 min at 500
In vitro pharmacological treatments
We used synthetic 17β-oestradiol (Sigma E-2758, St Louis, MO, USA), the most potent and receptor-specific form of oestrogen, to study the effects of oestrogen. The 17β-oestradiol was initially dissolved in ethanol at a concentration of 10−1
Immunocytochemistry
Following pharmacological treatments, cells were fixed for 10 min in 4% paraformaldehyde at room temperature, rinsed, then permeabilized for 15 min in 0.2% Triton X-100 and blocked for 1 h in 1× PBS containing 5% NGS. Coverslips were incubated in rabbit anti-p-ERK (no. 4370, 1:200; Cell Signaling Technology, Danvers, MA, USA) and mouse anti-NeuN (1:1000; Chemicon) in 1× PBS containing 3% NGS overnight at 4 °C followed by washing and secondary antibody staining with goat antirabbit Alexa Fluor 568 and goat antimouse Alexa Fluor 488 (1:200; Molecular Probes) for 30 min at 25°C. Images were acquired at 20× magnification using a Nikon confocal microscope. A minimum of 100 cells from three independent cultures were analysed for each experimental group. Cells immunoreactive for NeuN were scored as p-ERK positive or negative by an observer who was unaware of the treatment group.
Orofacial sensitivity assay
Mechanical sensitivity of the orofacial region was assessed using the behavioural assay described previously (20). Briefly, rats were conditioned to traverse a clear plastic tunnel attached to a behavioural arena in order to gain access to a bottle containing 0.1
CFA and selective agonist treatment
Two weeks following ovariectomy, rats were given a single 50-μl injection of CFA (
Statistical analysis
Data were analysed using Graph Pad Prism version 4.0 software (Graph Pad, San Diego, CA, USA). Behavioural data and immunocytochemistry were analysed using one-way
Results
GPR30 expression in rat trigeminal ganglion
In order to determine whether GPR30 protein is expressed in the trigeminal ganglion, we performed Western blots on homogenates of trigeminal ganglia from ovariectomized female Sprague–Dawley rats using a GPR30-specific antibody. Membranes showed a single band (Fig. 1) that was eliminated by preincubation of the antibody with the immunizing peptide, indicating that the antibody is specific for GPR30 (data not shown).
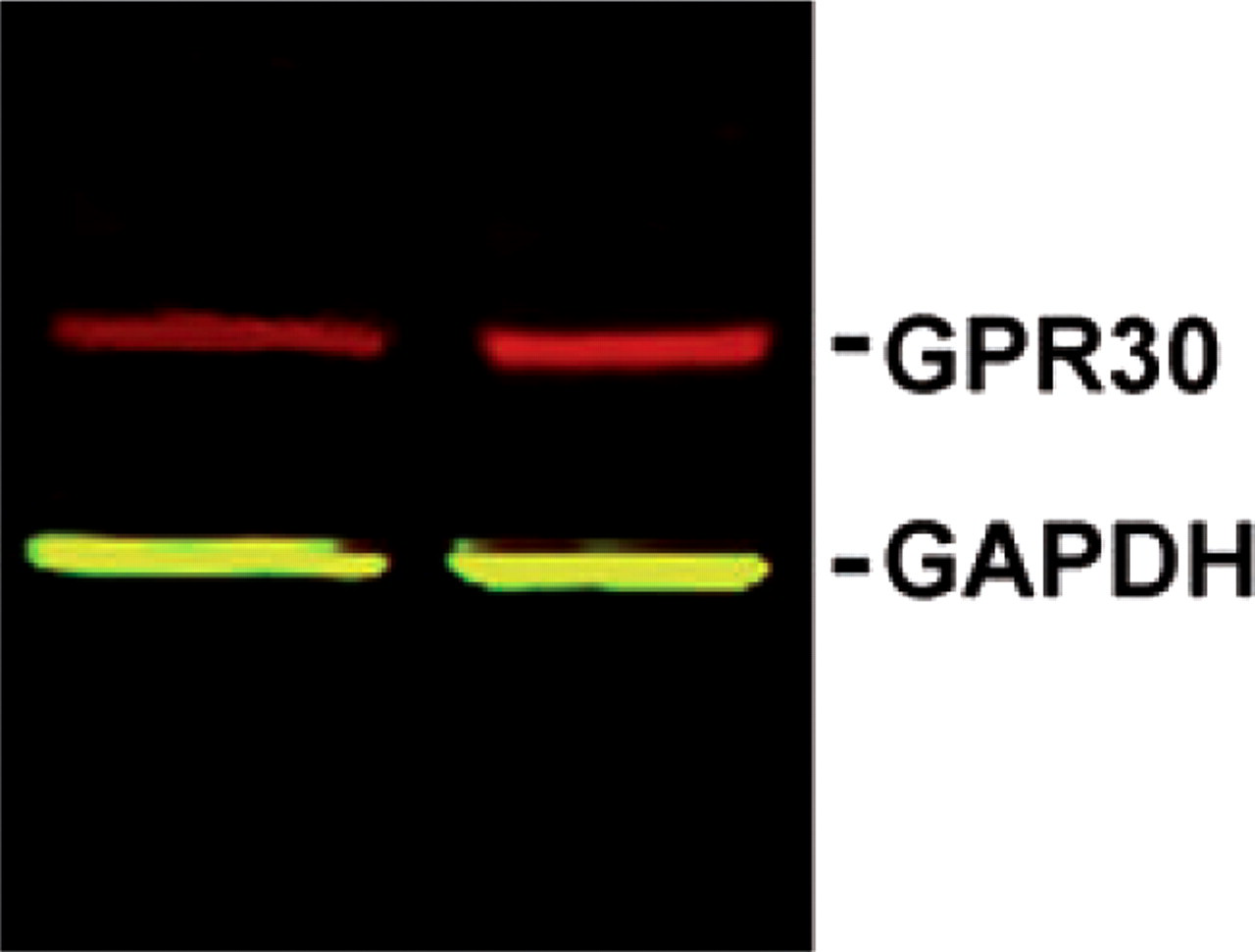
GPR30 protein is present in rat trigeminal ganglion. Western blot of trigeminal ganglion lysates from ovariectomized female rats labelled with antibodies to GPR30 (red) and the loading control GAPDH (green).
GPR30 expression in trigeminal neuron subpopulations
To determine the trigeminal ganglion cell types that express GPR30, immunohistochemistry was performed on frozen sections using the same GPR30 antibody used in Western blots. GPR30 immunoreactivity was localized to neurons (Fig. 2). We observed immunofluorescence throughout the neuronal cytoplasm of positively labelled cells. Immunofluorescence was also specific for GPR30, as preincubation of the antibody with the immunogen peptide eliminated staining (data not shown).
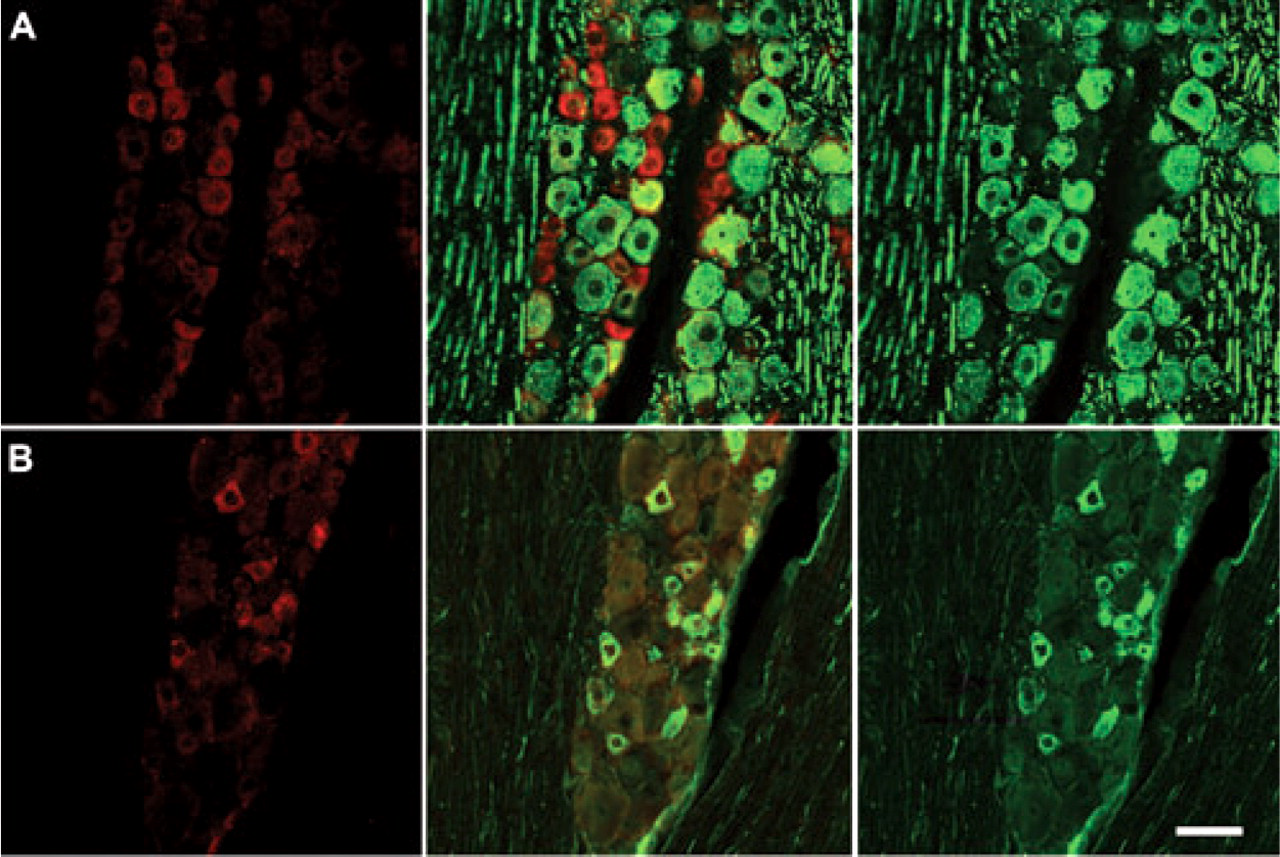
GPR30 is localized to unmyelinated neurons in the trigeminal ganglion. (A) Double-label immunohistochemistry for GPR30 and NFH, a marker of myelinated neurons. There is a small degree of overlap between GPR30 and NFH. (B) Double-label immunohistochemistry for GPR30 and peripherin. GPR30 and peripherin are highly co-localized. Panels on the left are single-labelled for GPR30. The middle row of panels contains merged images of the left and right columns. The right row of panels are single-labelled neurofilament H (top) or peripherin (bottom). Scale bar (50 μm) applies to all images.
To characterize the expression of GPR30 within trigeminal ganglion subpopulations, we used double-label immunohistochemistry to determine the colocalization of GPR30 with peripherin, a marker of neurons with unmyelinated axons and neurofilament H (neurofilament 200), a marker of neurons with myelinated axons (Fig. 2). The localization of GPR30 expression was primarily restricted to neurons with unmyelinated axons. Seventy-six per cent of GPR30+ neurons were peripherin positive, and 73% of peripherin-positive neurons were immunoreactive for GPR30. Thirty-four per cent of GPR30+ neurons were NFH+, and GPR30 was present in 32% of NFH+ neurons.
In order to investigate the neuronal phenotypes that express GPR30 in the trigeminal ganglion, we measured the diameters of cells that were immunoreactive and non-immunoreactive for GPR30. GPR30 was expressed across a broad range of cell sizes (Fig. 3), but the average diameter of GPR30+ cells (23.9 ± 0.8 μm) was significantly smaller (

GPR30 is present in small diameter neurons. Histogram shows cell diameter measurements for trigeminal ganglion neurons that were GPR30 immunoreactive (black bars) and GPR30 non-immunoreactive (gray bars).
ERα immunohistochemistry
Because ERα is a putative mediator of oestrogen-modified nociception, we used immunohistochemistry to label ERα in the trigeminal ganglion. We observed prominent ERα immunoreactivity throughout the trigeminal ganglion that was localized to neurons. ERα immunoreactivity was present in both nuclear and cytoplasmic compartments (Fig. 4A). On visual inspection, cells with a predominately cytoplasmic ERα immunoreactivity pattern were principally small neurons, whereas cells with a predominately nuclear ERα staining pattern had larger diameters. In order to quantify the distribution of immunoreactivity, we measured the diameters of neurons with cytoplasmic-predominant and nuclear-predominant staining patterns. We defined a nuclear predominant pattern as a cell with a nuclear intensity greater than twice that of the cytoplasmic intensity. Figure 4B shows the diameters of these neuronal populations. Cells containing prominent cytoplasmic ERα immunoreactivity had a mean diameter of 19.1 ± 0.3 μm, which was significantly smaller (

Cytoplasmic and nuclear ERα immunoreactivity is present in the trigeminal ganglion. (A) Immunohistochemistry for ERα in trigeminal ganglion from ovariectomized female rat. ERα staining was present in neurons in both cytoplasmic and nuclear locations. (B) Histogram showing cell diameter measurements for neurons that were immunoreactive for nuclear and cytoplasmic ERα. Cytoplasmic ERα was present in predominately small neurons and nuclear ERα was present in larger neurons.
Colocalization of GPR30 and ERα
We used double-label immunohistochemistry to determine the colocalization of GPR30 and ERα. ERα and GPR30 were present in distinct but partially overlapping populations. We observed neurons that expressed both GPR30 and ERα, and neurons that expressed only ERα or GPR30 (Fig. 5). Ten per cent of trigeminal ganglion neurons expressed both GPR30 and ERα, 22% expressed ERα and 35% expressed GPR30.

GPR30 and ERα are present in distinct but overlapping populations in the rat trigeminal ganglion. Immunohistochemistry for GPR30 (red) and ERα (green) shows ERα positive (A), GPR30 positive (B) and ERα/GPR30 double-positive (C) neurons. Scale bar (50 μm).
ERK activation by selective agonists for ERα, ERβ and GPR30
Oestrogen has been demonstrated to activate ERK in primary cultures of dorsal root ganglion and trigeminal ganglion neurons (35, 36) and in the rat trigeminal ganglion
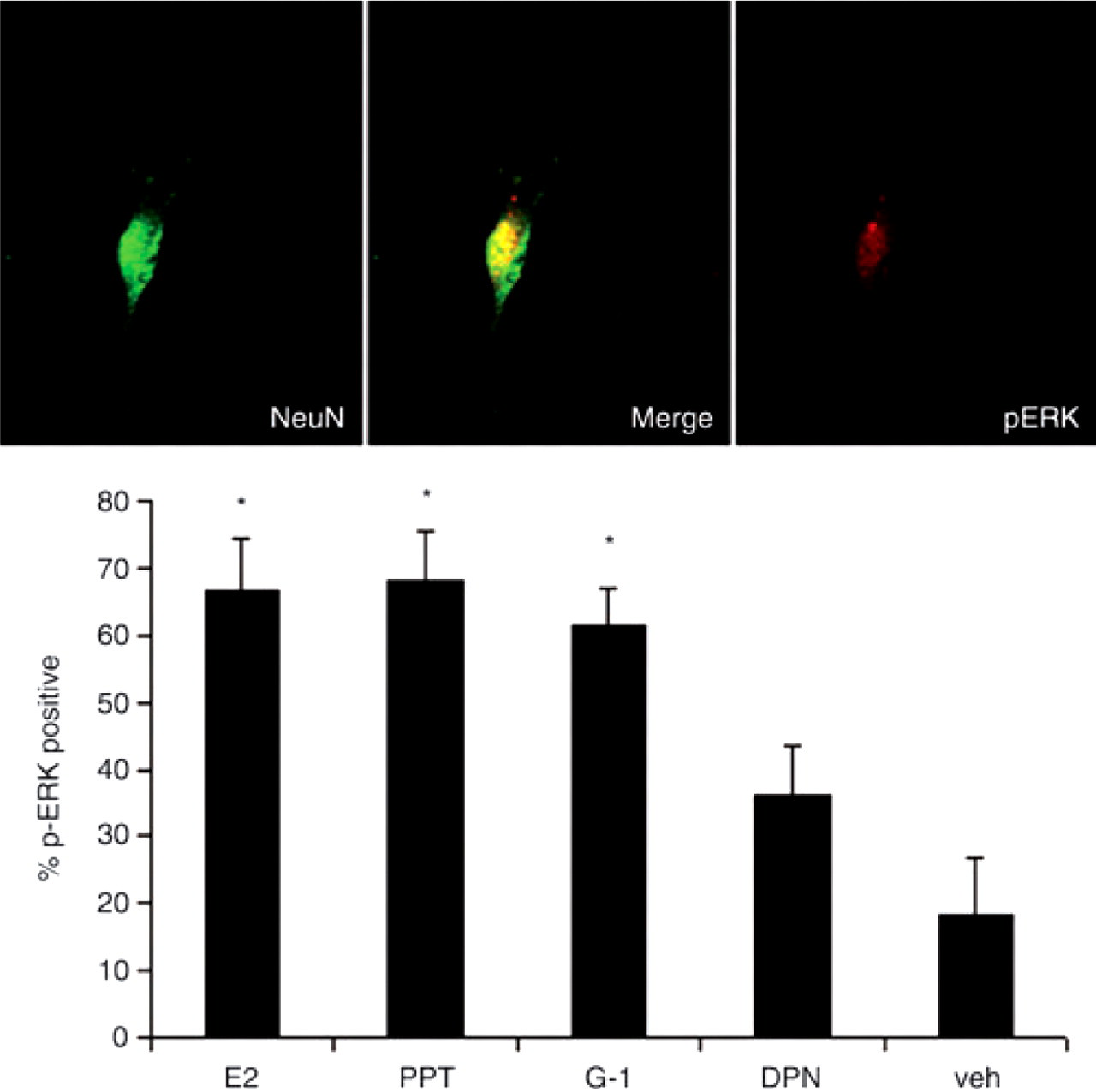
Selective activation of GPR30 or ERα activates ERK in trigeminal ganglion neurons
Selective agonists for GPR30 and ERα increase orofacial sensitivity
We have previously demonstrated that oestrogen increases facial allodynia through activation of ERK (25). To better characterize which ER mediates increased trigeminal sensitization, we examined the effects of PPT and G-1 on facial mechanical nociception following injections of CFA into the masseter, using the behavioural model described previously (20). We conducted a previous study to determine the monofilament force resulting in a 50% withdrawal threshold in ovariectomized rats (25). A sub-threshold filament was subsequently used for the testing of whisker pad in order to measure allodynia. In order to assess secondary allodynia, mechanical sensitivity of the whisker pad was measured using a 0.16-g monofilament 24, 48 and 72 h following CFA injection and selective agonist treatment. Injection of either PPT or G-1 enhanced withdrawal responses in the setting of peripheral inflammation at 24 h compared with vehicle-treated controls (Fig. 7).

Selective activation of GPR30 or ERα enhances orofacial allodynia. Specific agonists for ERα (PPT) and GPR30 (G-1) increased withdrawal response to stimulation of the whisker pad in CFA-treated ovariectomized female rats. For each animal, the left masseter was injected intra-muscularly with 50 μl CFA (1:1 in saline) and PPT (1 mg/kg), G-1 (1 mg/kg) or vehicle were injected subcutaneously. Withdrawal response to stimulation of the whisker pad with a von Frey filament (0.16 g) was assessed at 24 h after agonist administration. Data are shown as mean ±
Oestrogen effects on expression of GPR30 and ERα
In order to determine whether oestrogen treatment changes the protein content of ERs in the trigeminal ganglion, we used Western blotting to compare ERα and GPR30 in trigeminal ganglion samples from OVX and OVX+E2 rats, 24 h after E2 or vehicle administration. A significant increase was observed in levels of ERα in ganglia from oestrogen-treated animals compared with vehicle-treated controls (Fig. 8). Two size variants of ERα 66 kD and 90 kD, were present in the trigeminal ganglion. Both variants were up-regulated by oestrogen, 1.81-fold and 1.76-fold, respectively. GPR30 protein content of the trigeminal ganglion was not significantly altered by oestrogen replacement.

Oestrogen treatment increases expression levels of ERα, but not GPR30, in the trigeminal ganglion of ovariectomized female rats. Estradiol valerate (10 μg/kg in sesame oil) or vehicle was administered by subcutaneous injection and trigeminal ganglia collected 24 h later. (A) Representative Western blots of trigeminal ganglion from ovariectomized rats treated with oestrogen or vehicle. (B) Effects of oestrogen on protein level of the 66 kD ERα isoform. (C) Effects of oestrogen on the protein level of the 90 kD ERα isoform. (D) Effects of oestrogen on GPR30 protein levels. Data are shown as mean integrated intensity relative to GAPDH ±
Inflammation effects on expression of GPR30 and ERα
In order to determine whether peripheral inflammation of the trigeminal nerve alters expression of ERα and GPR30 in the trigeminal ganglion, we compared ERα and GPR30 protein levels using Western blots of trigeminal ganglion samples taken from groups of ovariectomized rats with CFA-injected or vehicle-injected masseter muscles. Inflammation of the masseter increased GPR30 expression 2.25-fold in the trigeminal ganglion compared with vehicle-treated controls, whereas it did not significantly change ERα expression in the trigeminal ganglion (Fig. 9).

Peripheral inflammation increases expression of GPR30 but not ERα in trigeminal ganglion from ovariectomized female rats. CFA (1:1 in saline) or vehicle was administered intra-muscularly into the masseter muscle and trigeminal ganglia were collected 24 h later. (A) Representative Western blots of trigeminal ganglion from rats with or without masseter inflammation. (B) Effects of inflammation of the masseter muscle on GPR30 protein level. (C and D) Effects of inflammation of the masseter muscle on ERα protein level for the 66 kD (C) and the 90 kD (D) isoforms. Data are shown as mean integrated intensity relative to GAPDH ±
Discussion
Although it is well established that oestrogen modifies orofacial pain, the type and location of receptors mediating these changes have remained unclear. Results of the current study support roles for both the novel ER GPR30 and ERα in primary sensory neurons in oestrogen-modified nociception. Furthermore, our data show that these receptors are differentially regulated by oestrogen and inflammation, which may have important implications for the modulation of trigeminal pain during the menstrual cycle.
GPR30 is present in the trigeminal ganglion
Our results show that GPR30 is present in the trigeminal ganglion of female rodents. GPR30 expression was localized to small, unmyelinated, peripherin-positive neurons, suggesting that GPR30 is localized to nociceptors in the trigeminal ganglion. Identification of a novel ER in the sensory pathway is potentially significant to oestrogen-modified trigeminal sensitization. Female animals are more sensitive to trigeminal nociception than male (37, 38), especially during the high oestrogen phase of the oestrous cycle (39, 40). Furthermore, oestrogen replacement increases excitability of trigeminal afferents and enhances inflammatory sensitization of the trigeminal system (25, 41, 42). The presence of GPR30 in likely nociceptors suggests that GPR30 is a possible mediator of oestrogen-modulated peripheral sensitization.
ERα is present in both cytoplasmic and nuclear compartments
ERα immunoreactivity was present in both cytoplasmic and nuclear compartments of trigeminal ganglion neurons. Previous studies of ERα in the rat trigeminal ganglion have restricted analyses to cells with nuclear labelling, discounting the potential importance of cytoplasmic ER immunoreactivity (12, 35). These results are intriguing in light of the dual role of ERα in both classical nuclear pathways and ‘non-genomic’ actions through signal transduction pathways. ERα-dependent activation of signal transduction molecules is thought to originate from receptors sequestered in signalling complexes in the cytoplasm or at the plasma membrane (43). Previous studies have shown that oestrogen modifies nociception through non-genomic mechanisms by activating ERK in the trigeminal ganglion (25).
The current results suggest that this mechanism predominates in nociceptive neurons, whereas, in larger diameter neurons, where ERα immunoreactivity is more prominent in the nucleus, oestrogen may preferentially function through direct genomic mechanisms.
ERα and GPR30 are expressed in distinct populations
Because the present data show that cytoplasmic ERα and GPR30 are expressed in small-diameter neurons, the colocalization of ERα and GPR30 in the trigeminal ganglion was analysed. Results show that ERα and GPR30 are present in distinct, but partially overlapping, neuronal populations, and that the majority of small trigeminal neurons express one of these receptors. Expression of either ERα or GPR30 by the majority of nociceptors in the trigeminal ganglion suggests a basis for the high sensitivity of the trigeminal system to oestrogen.
ERK is activated through GPR30 and ERα
Results show that selective agonists for either GPR30 or ERα induce rapid ERK activation in trigeminal ganglion neurons, suggesting that ERK activation mediated by oestrogen (25, 35) can occur through either ERα or GPR30. Furthermore, these data suggest that trigeminal GPR30 is functional in activating signal transduction pathways in trigeminal ganglion neurons, which may be relevant to oestrogen's modification of nociception.
There is considerable precedent for the activation of ERK by either ERα or GPR30 in many oestrogen-sensitive cell types (44–46), and signalling through either receptor has been demonstrated in the same cell type (44).
ER-evoked activation of ERK may be significant to the pathogenesis of oestrogen-modified sensitization. ERK phosphorylation in sensory neurons is a specific marker of nociceptive stimulation (17) and a mediator of mechanical hyperalgesia in pain models of peripheral nerve injury and inflammation (47, 48). As oestrogen increases mechanical allodynia of the orofacial region through activation of ERK (20, 25), ERK phosphorylation resulting from ERα or GPR30 activation may be important to the pathogenesis of trigeminal pain. Activation of ERK in the unmyelinated neurons expressing ERα or GPR30 could lead to increases in neuronal excitability, resulting in allodynia.
GPR30 and ERα participate in trigeminal sensitization
The results of the current study demonstrate that selective agonists for either GPR30 or ERα enhance secondary mechanical allodynia of the orofacial region, suggesting that both GPR30 and ERα are capable of mediating pro-nociceptive responses to oestrogen in the trigeminal system. The data from
By identifying GPR30 as a putative modulator of sensitization in the trigeminal system, the current results reveal increased complexity of pro-nociceptive effects of oestrogen in the trigeminal system. The presence of GPR30 in the trigeminal ganglion presents a novel pathway through which oestrogen may modulate the activity of sensory neurons, and which may be relevant to effects of hormonal fluctuations on migraine and facial pain.
ERα is increased by oestrogen, whereas GPR30 is increased by inflammation
Results show that inflammation in the trigeminal distribution up-regulates levels of GPR30 protein in the trigeminal ganglion. Although the effect of inflammation on GPR30 expression has not been previously addressed, oestrogen and inflammation have been shown to have additive effects on trigeminal excitability and nociceptive responses (37, 39, 42). The current data suggest that peripheral inflammation during episodes of trigeminal pain may increase GPR30 expression and lead to enhanced modulation of nociception by oestrogen. Furthermore, since inflammation did not modify expression levels of ERα, peripheral trigeminal inflammation may shift the balance of oestrogen signalling in trigeminal neurons towards GPR30.
The current data show that the presence of oestrogen increases levels of ERα protein in the trigeminal ganglion. The relationship between serum oestrogen and ERα receptor expression is complex and appears to be highly dependent on both the oestrogen level and tissue studied, but these data are consistent with several previous studies showing that oestrogen increases ERα in sensory neurons (12, 49). GPR30 expression, by contrast, was not changed by oestrogen replacement. This result is consistent with studies of hamster ovary showing that GPR30 expression was not changed by oestrogen replacement (50).
In conclusion, the results of this study demonstrate that both GPR30 and ERα pathways can modulate trigeminal nociception, providing new insight into the complex relationship between oestrogen and trigeminal pain.
Footnotes
Acknowledgements
This project was supported by NIH Grant P20 RR016475 from the INBRE Program of the National Center for Research Resources, R21 DE01582, National Headache Foundation, KUMC Lied Foundation and Biomedical Research Training Program.